
不同固体培养基配方对台湾含笑花粉萌发的影响及台湾含笑杂交亲和性分析
2022-05-24 10:37刘向东殷云龙
植物资源与环境学报 2022年2期
刘向东, 殷云龙
〔江苏省中国科学院植物研究所(南京中山植物园), 江苏 南京 210014〕
中国的木兰科(Magnoliaceae)含笑属(MicheliaLinn.)植物种质资源丰富,共40余种,主要分布在亚热带植被区[1]151-152。该属植物花色丰富、花香浓郁、四季常绿,不仅是优良的园林绿化植物,而且是富含单萜烯类、倍半萜烯及含氧衍生物等化学成分的经济植物[2-4]。中国科学院华南植物园在20世纪90年代建立了国内首个木兰科植物种质基因库[2],从此以后,木兰科含笑属植物一直是国内开展树木资源引种保育和推广应用的目标植物类群之一。
台湾含笑〔M.compressa(Maxim.) Sarg.〕为木兰科含笑属常绿乔木,原产于中国台湾地区,生于海拔200~2 600 m的阔叶林中,是台湾地区主要用材树种之一[1]187-189。1982年,江苏省中国科学院植物研究所科研人员从福建引进了1株台湾含笑实生苗,该实生苗在南京表现出较好的适应性,目前已通过嫁接和种子繁育等方式进行繁殖,并已成功引种到苏北地区[5]。然而,与常见的木兰科植物相比,台湾含笑的花朵较小,花色不够鲜艳,观赏价值偏低[6]。将台湾含笑与其近缘种和远源种进行杂交育种,有望改善台湾含笑的观赏性状,提高台湾含笑的利用价值。
近年来,国内学者在含笑属植物杂交育种方面取得了一定进展。例如:龚洵等[7]以云南含笑(M.yunnanensisFranch. ex Finet et Gagnep.)为母本、灰岩含笑(M.calcicolaC. Y. Wu)为父本进行杂交育种,成功育成‘雏菊含笑’(‘Chuju hanxiao’)和‘春月含笑’(‘Chunyue hanxiao’)2个新品种。研究表明:木兰科植物存在属间远缘杂交亲和性,尤其是玉兰属(YulaniaSpach)与含笑属的杂交。例如:二乔玉兰〔Y.×soulangeana(Soul.-Bod.) D. L. Fu〕与南亚含笑(M.doltsopaBuch.-Ham. ex DC.)以及紫玉兰〔Y.liliiflora(Desr.) D. L. Fu〕与云南含笑具有杂交亲和性,不存在生殖隔离[8];‘红元宝’玉兰(Magnoliasoulangeana‘Hongyuanbao’)与金叶含笑(M.foveolataMerr. ex Dandy)也具有杂交亲和性[9]。然而,目前关于木兰科植物的属间远缘杂交和属内种间杂交育种研究较少[10],应加强相关研究。
为此,本文比较了不同质量浓度蔗糖和硼酸组合以及不同质量浓度CaCl2固体培养基对台湾含笑花粉萌发的影响;对台湾含笑及同属植物深山含笑(M.maudiaeDunn)和阔瓣含笑〔M.cavalerieivar.platypetala(Hand.-Mazz.) N. H. Xia〕以及玉兰属植物白玉兰〔Y.denudata(Desr.) D. L. Fu〕的花粉活力进行比较,并对台湾含笑同株异花授粉组、台湾含笑异株授粉组及以台湾含笑为母本的3个杂交组的杂交亲和性和结实情况进行观察和比较,以期为台湾含笑的育种和遗传改良奠定研究基础,并为木兰科含笑属的种间杂交和远缘杂交提供新证据。
1 材料和方法
1.1 材料
于2021年在江苏省中国科学院植物研究所苗圃地(东经118°49′52.67″、北纬32°03′32.96″,海拔40 m)进行杂交实验。该研究地属亚热带季风气候,四季分明,年降水量1 200 mm,年均气温15.4 ℃,年无霜期250 d左右,年均日照时数1 933.0 h。
各杂交组的母本为种植于江苏省中国科学院植物研究所苗圃地中长势一致的5株树龄20 a的台湾含笑植株。台湾含笑同株异花授粉组的花粉采自江苏省中国科学院植物研究所苗圃地内1株树龄20 a的台湾含笑实生苗,台湾含笑异株授粉组的花粉采自南京中山植物园蔷薇园内1株树龄40 a的台湾含笑实生苗,台湾含笑×白玉兰组的花粉采自南京中山植物园蔷薇园内1株树龄60 a的白玉兰植株,台湾含笑×深山含笑组的花粉采自南京禄口机场内1株树龄35 a的深山含笑植株,台湾含笑×阔瓣含笑组的花粉采自南京中山植物园球宿根花卉园内1株树龄12 a的阔瓣含笑植株。所有供试植株均长势良好,无病虫害且能够正常开花结实。
1.2 方法
1.2.1 花粉采集 于2021年2月至3月,分别在各种类供试植株的盛花期(植株45%以上的花开放)采摘未完全绽放的新鲜花朵,每株样株各采摘约50朵花;除去花被片,将带有雄蕊群的花柱置于硫酸纸上,置于阴凉通风处,等待药室张开,花粉自然散落;收集花粉,于4 ℃干燥条件下保存、备用。每个种类重复取样3次。
1.2.2 不同固体培养基配方对台湾含笑花粉萌发的影响 分别对不同质量浓度蔗糖和硼酸组合固体培养基(含质量分数1%琼脂糖,pH 7.0)中台湾含笑的花粉萌发情况进行比较,设置蔗糖质量浓度50、100、150和200 g·L-1,硼酸质量浓度100、200、300和400 mg·L-1,共16个组合,以不含蔗糖和硼酸的固体培养基(含质量分数1%琼脂糖,pH 7.0)为对照。此外,以含150 g·L-1蔗糖、100 mg·L-1硼酸和质量分数1%琼脂糖的固体培养基(pH 7.0)为基本培养基,对添加质量浓度0(对照)、100、200、300和400 mg·L-1CaCl2的培养基中台湾含笑的花粉萌发情况进行了比较。每个处理培养花粉约0.1 g,于25 ℃避光条件下培养8 h后,在OLYMPUS BX53F显微镜(日本Olympus公司)下观察花粉萌发情况。以花粉管长度大于花粉直径作为花粉萌发的标准,统计萌发花粉数,据此计算花粉萌发率,计算公式为花粉萌发率=(视野内的萌发花粉数/视野内花粉总数)×100%。使用Mage Analysis System V1.1.4软件测量花粉管长度。每个处理重复3次,每个重复至少统计5个视野中的花粉。
1.2.3 不同杂交组父本花粉活力比较 参考前人的研究结果[11-13]设置不同父本的固体培养基配方。阔瓣含笑花粉使用含质量浓度100 g·L-1蔗糖和100 mg·L-1硼酸的固体培养基(含质量分数1%琼脂糖,pH 6.0)进行培养,深山含笑花粉使用含质量浓度150 g·L-1蔗糖、300 mg·L-1硼酸和300 mg·L-1CaCl2的固体培养基(含质量分数1%琼脂糖,pH 5.0)进行培养,白玉兰花粉使用含质量浓度10 g·L-1蔗糖和200 mg·L-1硼酸的固体培养基(含质量分数1%琼脂糖,pH 6.4)进行培养,台湾含笑花粉使用前面实验筛选的最适固体培养基进行培养。每个处理培养花粉约0.1 g,于25 ℃避光条件下培养8 h后,在OLYMPUS BX53F显微镜下观察花粉萌发情况。统计萌发花粉数,据此计算花粉萌发率,根据花粉萌发率判断花粉活力。使用Mage Analysis System V1.1.4软件测量花粉管长度。每个处理重复3次,每个重复至少统计5个视野中的花粉。
1.2.4 人工授粉 人工授粉实验共设置5个杂交组,即台湾含笑同株异花授粉组、台湾含笑异株授粉组、台湾含笑×白玉兰组、台湾含笑×深山含笑组和台湾含笑×阔瓣含笑组。参照李颖婕[14]的方法,在台湾含笑盛花期选择母本样株朝阳方位的单花,在单花的花蕾期人工去雄后授粉,及时套袋并挂牌记录。每个杂交组合授粉70朵花。
1.2.5 不同杂交组杂交亲和性观察 采用苯胺蓝荧光染色法[15,16]观察不同杂交组的花粉在台湾含笑柱头上的萌发情况和花粉管的伸长情况。分别在授粉后1、2、3、4、6、8、10、12 h以及1、2、3、4、6、8 d取柱头,浸入装有FAA固定液〔V(甲醛)∶V(冰乙酸)∶V(体积分数50%乙醇)=1∶1∶8〕的离心管中至少24 h。每个杂交组每次取3个柱头。
将固定好的雌蕊轻轻分离出来,使用ddH2O清洗2~3次后,浸入质量体积分数5%NaOH溶液中,并置于60 ℃恒温水浴中软化脱色4 h;再使用ddH2O清洗2~3次后,用质量体积分数0.1%苯胺蓝溶液遮光染色10 min;用吸水纸吸去雌蕊表面多余的染液,在干净的载玻片上滴1~2滴甘油,压片后在OLYMPUS BX53F显微镜的紫外光源下观察花粉在柱头上的萌发情况和花粉管的伸长情况并拍照。
1.2.6 不同杂交组结实情况分析 于2021年5月下旬至6月上旬(即台湾含笑生理落果结束后)统计各杂交组的坐果数,参照王泽翻等[17]的方法计算坐果率,计算公式为坐果率=(坐果总数/授粉花朵总数)×100%。
同年10月中旬采集各杂交组的成熟果实(心皮开始开裂、假种皮变红),分别记录各杂交组的成熟果实总数及心皮总数和结实心皮总数,参照王泽翻等[17]的方法计算结实率和心皮结籽率,计算公式分别为结实率=(成熟果实总数/授粉花朵总数)×100%和心皮结籽率=(结实心皮总数/心皮总数)×100%。
将收集的果实置于阴凉通风处,待心皮自然开裂后剥出种子,用自来水浸泡2 d左右,待假种皮变软后将其洗去,统计各杂交组的种子总数。将种子晾干,随机选取50粒种子,使用万分之一分析天平称量总质量,重复称量5次,结果取平均值。根据称量结果计算种子的千粒质量。
1.3 数据统计分析
分别使用EXCEL 2010和Data Processing System 18.10软件对花粉萌发率和种子千粒质量进行数据统计和方差分析。
2 结果和分析
2.1 不同固体培养基配方对台湾含笑花粉萌发的影响
2.1.1 不同质量浓度蔗糖和硼酸组合对台湾含笑花粉萌发的影响 统计结果表明:台湾含笑花粉在不含蔗糖和硼酸的固体培养基中培养8 h未萌发,而在含不同质量浓度蔗糖和硼酸的固体培养基中培养8 h均能够萌发,各培养基中花粉的萌发率和花粉管长度见表1。由表1可见:在含150 g·L-1蔗糖和100 mg·L-1硼酸的固体培养基中,台湾含笑的花粉萌发率最高(46.4%),花粉管长度为236.0 μm;在含200 g·L-1蔗糖和200 mg·L-1硼酸的固体培养基中,台湾含笑的花粉萌发率最低(22.2%),花粉管长度最短(95.5 μm)。
由表1还可见:当固体培养基中硼酸质量浓度一定时,台湾含笑的花粉萌发率和花粉管长度总体上随着蔗糖质量浓度升高呈现明显的“单峰型”变化趋势;而当固体培养基中蔗糖质量浓度一定时,台湾含笑的花粉萌发率和花粉管长度却未随着硼酸质量浓度升高表现出明显的变化趋势。说明蔗糖质量浓度对台湾含笑花粉萌发的影响较大。

表1 含不同质量浓度蔗糖和硼酸的固体培养基对台湾含笑花粉萌发的影响
2.1.2 不同质量浓度CaCl2对台湾含笑花粉萌发的影响 统计结果(表2)表明:当固体培养基中不含CaCl2时,台湾含笑的花粉萌发率最高(44.6%),花粉管长度最长(260.1 μm)。总体来看,随着CaCl2质量浓度升高,台湾含笑的花粉萌发率和花粉管长度逐渐降低。说明CaCl2对台湾含笑的花粉萌发和花粉管伸长具有一定的抑制作用。
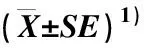
表2 含不同质量浓度CaCl2的固体培养基对台湾含笑花粉萌发的影响
2.2 不同杂交组父本花粉活力比较
根据5个杂交组父本花粉萌发率统计结果,同株异花授粉的台湾含笑花粉活力最高(76.2%);阔瓣含笑花粉活力较高(71.5%);深山含笑和异株授粉的台湾含笑花粉活力较低,分别为50.9%和46.4%;白玉兰花粉活力最低(43.9%)。
2.3 不同杂交组的杂交亲和性观察
对5个杂交组的杂交亲和性进行荧光显微观察,发现不同杂交组花粉的初始萌发时间、大量萌发时间及花粉管到达子房的时间均存在一定差异。
台湾含笑同株异花授粉组花粉的初始萌发时间最早,在授粉6 h开始萌发,在授粉2 d大量萌发;花粉管在授粉3 d汇成一束,到达花柱1/3处;花粉的整个萌发过程在5个杂交组中最长,花粉管在授粉8 d才能到达子房位置(图版Ⅰ-1~4)。
台湾含笑异株授粉组花粉的初始萌发时间较早,在授粉1 d开始萌发,在授粉2 d大量萌发;花粉管在授粉3 d汇成一束,到达花柱1/3处;花粉的整个萌发过程较长,花粉管在授粉6 d到达子房位置(图版Ⅰ-5~8)。
台湾含笑×白玉兰组花粉的初始萌发时间最晚且整个萌发过程较短,在授粉4 d才开始萌发并迅速大量萌发,花粉管内堆积大量胼胝质,阻碍花粉管的继续伸长;仅有少数花粉管在授粉6 d能够到达子房位置(图版Ⅰ-9,10)。
台湾含笑×深山含笑组花粉的萌发时间较早且整个萌发过程较短,在授粉1 d开始萌发,在授粉2 d大量萌发,且花粉管快速伸长并汇成一束,到达花柱1/3处;花粉管在授粉4 d即到达子房位置(图版Ⅰ-11~13)。
台湾含笑×阔瓣含笑组花粉的整个萌发进程较短,在授粉2 d开始萌发并迅速大量萌发,花粉管在授粉3 d汇成一束;花粉管在授粉4 d到达子房位置(图版Ⅰ-14~16)。
2.4 不同杂交组的结实情况分析
对5个杂交组的结实情况进行统计,结果见表3;各杂交组的成熟果实见图版Ⅰ-17。
统计结果表明:各杂交组生理性落果结束后,台湾含笑同株异花授粉组和台湾含笑×阔瓣含笑组的坐果率最高,均为100.0%;台湾含笑×深山含笑组和台湾含笑异株授粉组的坐果率较高,分别为75.0%和72.2%;台湾含笑×白玉兰组的果实完全脱落,坐果率为0.0%。各杂交组的结实率与坐果率接近,同样表现为台湾含笑同株异花授粉组和台湾含笑×阔瓣含笑组的结实率明显高于其他杂交组,台湾含笑×深山含笑组和台湾含笑异株授粉组的结实率相近。
各杂交组的心皮结籽率和种子千粒质量也表现出明显差异。台湾含笑同株异花授粉组的心皮结籽率最高,达84.4%,但该杂交组的种子千粒质量最低,仅87.0 g;台湾含笑×阔瓣含笑组和台湾含笑×深山含笑组的心皮结籽率相近,分别为50.3%和50.2%,这2个杂交组的种子千粒质量显著高于其他杂交组,分别为127.5和123.2 g;台湾含笑异株授粉组的心皮结籽率最低(43.6%),种子千粒质量居中(101.2 g)。

表3 台湾含笑不同杂交组的结实情况
3 讨 论
3.1 不同杂交父本的花粉活力差异
花粉活力高低是影响植物授粉成功与否的关键因子。高活性的花粉可以提高结实率,然而,由于受物种自身以及保存环境的湿度、温度、大气成分和大气压力等因子的影响,植物的花粉活力往往不能长久保持[18],不利于花期不同的物种间的杂交。因此,明确杂交父本的花粉活力是分析植物杂交亲和性的重要依据,也是避免因花粉活力低而导致结实率低的重要技术措施。本研究结果表明:在相同保存条件下,阔瓣含笑的花粉活力最高,深山含笑的花粉活力较高,2个种类的花粉活力均在50%以上,而台湾含笑和白玉兰的花粉活力均低于50%,花粉更容易失去活性。与台湾含笑异株授粉组相比,台湾含笑同株异花授粉组的坐果率和心皮结实率均明显提高,台湾含笑×深山含笑组和台湾含笑×阔瓣含笑组的坐果率和心皮结实率也明显提高,说明高花粉活力是台湾含笑杂交实验顺利开展的重要前提。
3.2 不同杂交组的杂交亲和性
杂交亲和性荧光显微观察发现,虽然各杂交组的花粉萌发时间和花粉管伸长速度存在明显差异,但这些杂交组的花粉均能够在台湾含笑柱头上萌发;白玉兰花粉萌发后积累了大量的胼胝质,阻碍花粉管继续伸长。说明白玉兰、阔瓣含笑和深山含笑均可在台湾含笑柱头上萌发,且阔瓣含笑和深山含笑与台湾含笑的杂交亲和性高。对5个杂交组结实情况的统计结果显示:台湾含笑×白玉兰杂交组未坐果,说明该杂交组不能够正常结实,不具备杂交亲和性;而台湾含笑×阔瓣含笑和台湾含笑×深山含笑2个杂交组均能够正常结出饱满种子且种子千粒质量显著高于其他杂交组,说明台湾含笑与阔瓣含笑和深山含笑的杂交亲和性较高。相关研究表明:染色体倍性差异直接影响不同物种间的杂交亲和性[19],倍性相同的物种往往比倍性差异较大的物种杂交亲和性高[20]。研究发现,含笑属植物染色体倍性较稳定,均为二倍体,染色体数为38[21],而木兰属植物染色体倍性差异较大,存在二倍体到六倍体多种染色体倍性,白玉兰为四倍体,染色体数为76[22]。因此,推测染色体倍性差异可能是台湾含笑与阔瓣含笑和深山含笑杂交亲和性高、与白玉兰不存在杂交亲和性的重要原因,具体原因有待深入研究。
此外,物种间杂交不亲和主要包括受精前杂交障碍和受精后杂交障碍2个方面。其中,受精前杂交障碍主要表现在花粉和柱头不亲和、花粉不能到达子房及无法成功受精等;受精后杂交障碍主要表现在胚胎败育、提前落果、杂交种子萌发率低或不萌发以及杂交后代植株不能正常生长等[23,24]。白玉兰花粉虽然能够在台湾含笑柱头上大量萌发,但在授粉后由于胼胝质异常沉积,导致多数花粉管变短、变粗,伸长异常,无法进入子房完成受精过程,仅少数花粉能顺利到达子房,完成受精过程,但果实却在生理落果期全部脱落,没有获得成熟果实,因此,台湾含笑与白玉兰杂交不亲和的主要原因是二者存在受精前杂交障碍,并存在一定程度的受精后杂交障碍。
综上所述,台湾含笑与同属种类深山含笑和阔瓣含笑的杂交亲和性较高,后期可以台湾含笑为核心种质开展含笑属植物的种质创新和杂交育种研究。另外,虽然台湾含笑与玉兰属种类白玉兰存在杂交不亲和问题,但其与玉兰属其他种类的杂交亲和性尚不清楚,有待深入研究。
猜你喜欢
无机盐工业(2022年7期)2022-07-11
食品安全导刊(2021年36期)2021-03-14
作文周刊·小学一年级版(2021年36期)2021-01-14
作文周刊·小学一年级版(2018年7期)2018-03-15
分析化学(2017年12期)2017-12-25
作文周刊·小学一年级版(2017年20期)2017-06-27
山东农业科学(2017年5期)2017-06-05
数字技术与应用(2016年10期)2017-04-01
东方教育(2016年14期)2017-01-16
作文周刊·小学一年级版(2016年14期)2016-05-10
