
构树转录因子BpbZIP1的鉴定及镉胁迫响应分析
2022-05-19 07:21陈思思谢牧洪崔茂凯李文凯徐正刚贾彩霞杨桂燕
植物研究 2022年3期
陈思思 谢牧洪 崔茂凯 李文凯 徐正刚 贾彩霞 杨桂燕
(西北农林科技大学林学院,陕西省经济植物资源开发利用重点实验室,杨凌 712100)
近些年,由采矿、废气排放、污水灌溉、重金属超标制品滥用等人为因素致使含有重金属的污水或者废料排放到土壤里,导致土壤受到严重的重金属污染。重金属在土壤中能引起土壤成分、结构和功能的改变,重金属离子通过抑制作物根系生长和光合作用,使作物减产,粮食的生产面积缩小。更为严重的是,重金属可以通过食物链转移到动物和人体内,严重危害动物和人的健康。随着我国工业化进程加快,我国土壤重金属污染问题日益加剧,在众多重金属污染中,镉(Cd)被确认为我国土壤的首要污染物。镉在自然环境中具有化学活性强、移动性大、毒性持久、易富集等特点,因此受镉污染土壤的治理难度很大。植物修复利用自然生长或遗传工程培育的植物来修复重金属污染土壤,是目前治理土壤污染的重要方法,具有较高的环保和经济价值。
构树()是重要的扶贫攻坚树种,具有生长迅速、环境适应性强、分布广泛、易于繁殖等优良特性,也是尾矿区和重金属污染地区的先锋树种和本土树种,对多种重金属元素具有良好的吸收和耐受力。但构树优良品系的开发不够,构树适应重金属的机理尚不清楚,这对发展构树生物修复不利。要充分发挥构树在重金属土壤修复中的功能,其前提是掌握构树重金属适应机制,通过常规或分子育种培育Cd高抗高耐受型构树新品种。因此,本研究旨在鉴定构树重要的Cd响应基因,为开展构树抗Cd分子育种提供候选基因。
转录因子(Transcription factors,TFs)是真核生物中重要的调控蛋白。在植物中,转录因子是许多信号网络中的关键调控因子,如植物生长、生物或非生物胁迫等。碱性亮氨酸拉链(basic leucine zipper,bZIP)是转录因子中重要的蛋白质家族之一。植物中的bZIP 一般由60~80 个氨基酸残基组成,含有1 个碱性结构域和亮氨酸拉链结构,氨基酸残基组成的碱性结构域与特异DNA 序列相结合,起核定位信号作用,亮氨酸拉链形成两亲性的α 螺旋结构,参与bZIP 蛋白与DNA 结合之前的二聚体化。在前期对构树响应Cd 胁迫的转录组学研究中,我们获得了受Cd 胁迫上调表达的众多基因,其中包括碱性亮氨酸拉链(basic leucine zip⁃per,bZIP)转录因子家族中的若干成员。
现代分子生物研究显示,bZIP 是植物生长过程的重要调节剂,如组织分化、细胞伸长、病原体防御、激素信号传导、光反应以及渗透控制。针对bZIP 调控植物抗逆的研究显示,bZIP 转录因子可参与抗病、抗寒、抗旱、耐盐和抗重金属离子等多种逆境胁迫调控。如,水稻()中过表达显著提高了植株对干旱、盐和PEG 渗透胁迫的耐受性,锌铁转运蛋白基因和参与水稻Zn和CdCl的吸收和转运;大豆()参与大豆耐药性、耐盐性、干旱耐受性的调控;苎麻()在响应高盐胁迫时起正向调控作用,在植物响应干旱和重金属Cd 胁迫时起负向调控作用。因此,本研究对响应Cd胁迫表达较为突出的1个bZIP成员(命名为)进行Cd胁迫下的表达和转基因酵母的抗Cd分析,以期为揭示构树Cd适应分子机制和抗逆分子育种提供候选基因。
1 材料与方法
1.1 材料与处理
植物材料:同批次构树组培苗转移至土壤生长至2年。
处理方法:使用分析纯的CdCl配置150 μmol·L的CdCl溶液,对构树进行胁迫处理,在处理后0、3、12、24、48、72 h 等时间点分别取根、茎、叶,将根、茎、叶用液氮速冻后,于冰箱中-80℃保存备用,每个处理包含5棵植株。
1.2 BpbZIP1基因的克隆与分析
在以构树Cd 胁迫下的转录组筛选出的上调表达基因中,筛选bZIP 家族成员,选择其中1 条表达值较高的基因(命名为)进行分析。基因的开放读码框(ORF)用ORF finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)确定。根据基因ORF 两端序列设计引物-和-(见表1),进行PCR 扩增。产物经回收纯化后与pMD-18T 载体连接并转化大肠杆菌DH5α感受态细胞。挑取阳性克隆扩大培养进行菌液PCR验证,对可获得目的片段的克隆测序。

表1 本研究涉及引物Table 1 The primers used in the study
利用Expasy ProtParam(http://web.expasy.org/protparam/)对确认的基因序列特征进行分析。利用BLASTP(http://blast.ncbi.nlm.nih.gov/Blast)进行序列同源性搜索。利用MEGA7 构建系统进化树。使用ClustalX 进行蛋白质多序列比对。使用CD-Search(https://www.ncbi.nlm.nih.gov/Struc⁃ture/cdd/wrpsb. cgi)和MEME(https://meme-suite.org/meme/tools/meme)进行BpbZIP1 的保守结构域分析。利用在线软件SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)推 测BpbZIP1 蛋白质二级结构,利用Swiss Model(https://swissmod⁃el.expasy.org/)推测BpbZIP1 蛋白质的三维结构。利用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bio⁃inf/Cell-PLoc-2/)进行蛋白质亚细胞定位预测。
1.3 BpbZIP1基因的表达分析
对1.1 中收集样品进行基因表达量分析。RNA 提取采用CTAB 法进行,RNA 经DNA 消化酶处理后采用PrimeScriptRT reagent Kit(CWBIO,康为世纪,中国)反转录为cDNA,稀释10倍后用作qRT-PCR 的模板。qRT-PCR使用SYBR Green Real time PCR Master mix(CWBIO)进行,内参基因为。所用引物如表1所示,定量引物为DL-F和DL-R。定量反应所用仪器为Applied Biosystems生产的StepOneReal Time PCR System。反应程序为:94 ℃预变性30 s;94 ℃变性12 s,60 ℃退火45 s,72 ℃延伸45 s,45个循环;81 ℃读板1 s,每个样品重复3次。采用2法对定量结果进行相对分析,表示为相对于内参基因相对于对照的表达值。数据使用SPSS 软件包(SPSS,Chicago,Illinois,USA)分析。样品变异性用标准偏差表示。不同时间点与0 h之间的表达差异用检验分析(<0.05)。
1.4 酵母表达载体构建及胁迫分析
根据基因序列特征及pYES2 酶切位点特性设计酵母表达载体引物BpbZIP1-JM-F 和BpbZIP1-JM-R(见表1)。通过PCR 反应获得含有酶切位点的序列,经酶切纯化后与pYES2连接获得重组载体pYES2-BpbZIP1。将pYES2-BpbZIP1 和空pYES2 载体分别转入酵母INVSC1中,分别记为INVSC1(pYES2-BpbZIP1)和INVSC1(pYES2),后者为对照。本研究所用工具酶均为宝生物(Takara)公司产品。
分 别 挑 取INVSC1(pYES2-BpbZIP1)和IN⁃VSC1(pYES2)单克隆置于含2%葡萄糖的SC-Ura液体培养基中,30℃震荡培养16 h,收集酵母菌体重悬于含2%半乳糖的SC-Ura液体培养基中,且调整OD=0.5,30℃继续震荡培养至OD=1.8,分别收集菌体进行Cd 胁迫处理。即,取相同量的上述INVSC1(pYES2-BpbZIP1)和INVSC1(pYES2)菌体,分别在含有0、0.2%、0.4%、0.6%、0.8%、1.0% CdCl的SC-Ura 液体培养基(含2%葡萄糖)中震荡培养30 h,测定酵母生长活性(OD)。数据使用SPSS软件包分析。样品变异性用标准偏差表示。相同处理下INVSC1(pYES2-BpbZIP1)和INVSC1(pYES2)之间的差异显著性用检验分析(<0.05)。
2 结果与分析
2.1 BpbZIP1基因基本生物信息
通过对构树转录组筛查鉴定获得基因(GeneBank 登录号:MW567468),根据获得的cDNA 序列设计引物BpbZIP1-F/R 进行PCR 验证,确定该基因序列ORF长1 713 bp,编码的蛋白包含570 个氨基酸,分子量为62 902.38 Da,等电点为4.62。保守域分析发现该蛋白有bZIP 结构域(见图1)。Swiss Model 同源建模预测的BpbZIP1 蛋白三维结构是典型的亮氨酸拉链结构(见图2:A~B),BpbZIP1 二级结构的主要构成元件为无规则卷曲和螺旋,占总氨基酸的48.95% 和39.12%,其余延伸链和转角分别占8.25%、3.68%(见图2C)。经同源搜索并进行进化分析,显示BpbZIP1蛋白与拟南芥()ZIP同源,且与AtbZIP1 的进化关系较近(见图3)。将BpbZIP1蛋白与拟南芥同源蛋白进行多序列对比,显示出高度的相似性(见图4A),同时BpbZIP1 蛋白与拟南芥同源蛋白都具有相同的保守结构域(见图4B)。对BpbZIP1 蛋白进行亚细胞定位预测,显示BpbZIP1蛋白为核蛋白。

图1 BpbZIP1蛋白保守结构域Fig.1 The conserved domain of BpbZIP1 protein
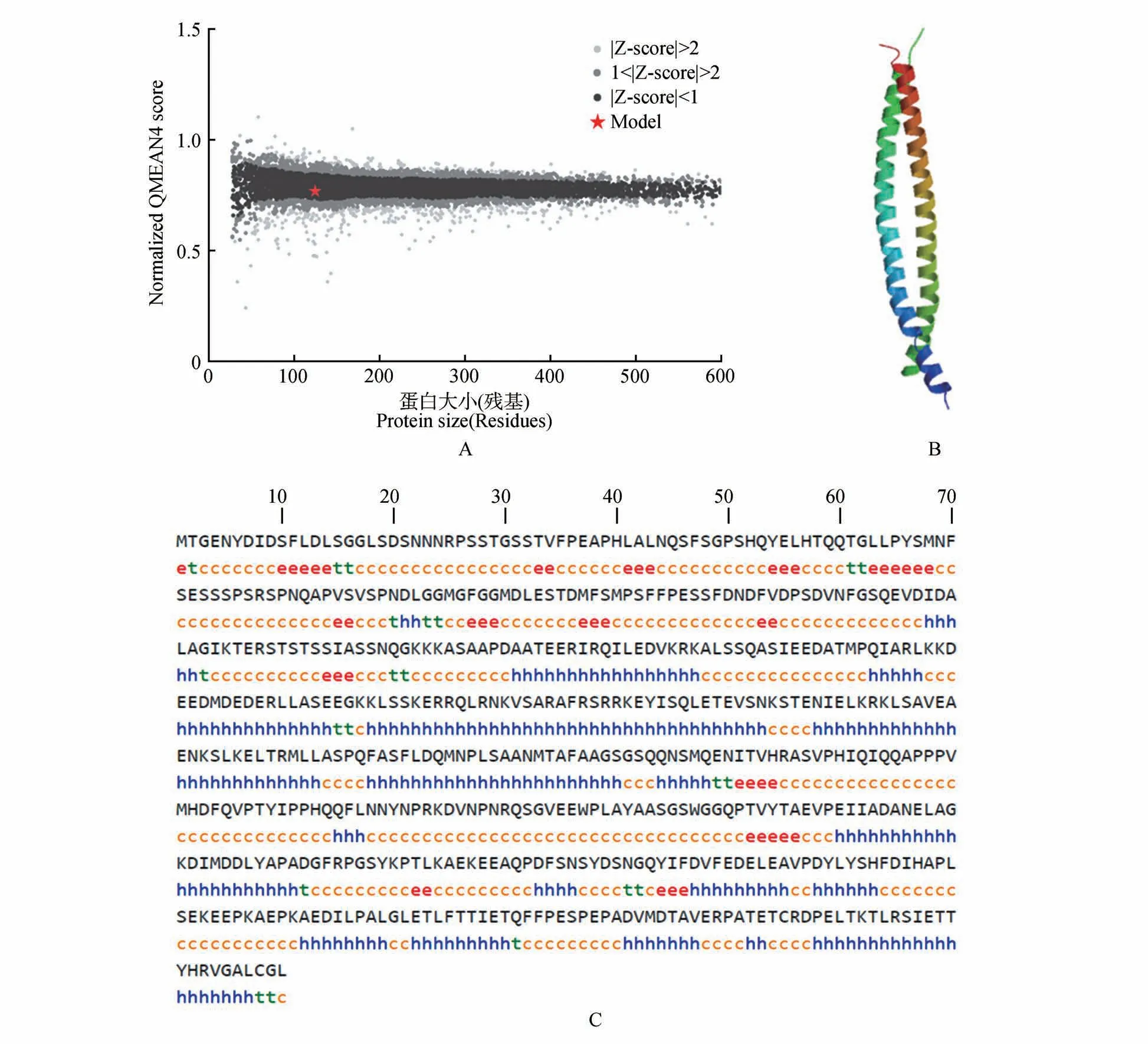
图2 BpbZIP1蛋白结构预测A.与非冗余PDB(Protein Data Bank)结构集比较;B.三维结构模型;C.二级结构;h.α螺旋;e.延伸链;t.β转角;c.无规则卷曲Fig.2 Predicted structure of BpbZIP1 protein A.Comparison with non-redundant set of PDB(Protein Data Bank)structures;B.3D structure model;C.Secondary structure;h. α helix;e.Extended strand;t.Beta turn;c.Random coil

图3 BpbZIP1与拟南芥相似蛋白的进化树括号内为登录号;Bb.构树;At.拟南芥Fig.3 Phylogenetic tree of BpbZIP1 protein GeneBank accession number in brackets;Bb.B.papyrifera;At.A.thaliana
2.2 BpbZIP1基因在Cd胁迫下的表达
提取CdCl胁迫不同时间点的根、茎、叶的总RNA 反转录为cDNA 用作qRT-PCR 分析。结果发现基因能被Cd 胁迫诱导表达,且在根、茎、叶中具有一定差异(见图4)。在根中,基因受胁迫后迅速被诱导,在3 h 达到最高水平,为对照组的17.4 倍;之后表达水平逐渐下降,但72 h 仍处于上调表达。在茎中,基因随时间延长的表达变化趋势同根中表现相似,均为倒‘V’字形,但整体上根中的表达要高于茎中的表达,且茎中出现的峰值在12 h,为对照的5.6 倍。在叶中,基因的转录水平随胁迫时间延长不断升高,在72 h为对照的5.1倍(见图5)。

图4 BpbZIP1与拟南芥同源蛋白多序列对比(A)及其motif(B)Fig.4 Multiple sequence alignment of BpbZIP1 and A.thaliana homologous proteins(A)and its motif logos(B)

图5 BpbZIP1基因在Cd处理下的表达水平**,*分别表示同一组织在不同时间点与0 h 之间的差异极显著(P<0.001)和显著(P<0.05)Fig.5 The expression level of BpbZIP1 gene under different time of CdCl2 treatment**,* mean that the difference between the same tissue at different time points and 0 h was extremely significan(tP<0.001)and signifi⁃can(tP<0.05)
2.3 BpbZIP1基因表达对酵母抗Cd性的影响
分 别 挑 取INVSC1(pYES2-BpbZIP1)和IN⁃VSC1(pYES2)单克隆在相同条件下培养并进行不同CdCl浓度处理,比较二者在Cd 胁迫下的生长活性。结果显示,在非胁迫条件下,INVSC1(pY⁃ES2-BpbZIP1)和INVSC1(pYES2)的生长没有明显区别,但在CdCl胁迫处理后,INVSC1(pYES2-Bp⁃bZIP1)和INVSC1(pYES2)的OD明显下降,且随着CdCl浓度增加,INVSC1(pYES2-BpbZIP1)和INVSC1(pYES2)的OD逐渐变小,但INVSC1(pY⁃ES2-BpbZIP1)一直高于INVSC1(pYES2),且差异显著(见图6)。表明基因的表达显著改善了酵母的CdCl胁迫性能。

图6 转BpbZIP1基因酵母在镉胁迫下的OD600**表示相同处理条件下INVSC1(pYES2-BpbZIP1)和INVSC1(pYES2)之间的差异性显著性(P<0.01)Fig.6 The OD600 of BpbZIP1 transgenic yeasts under different CdCl2 concentrations** indicates the significant difference between INVSC1(pYES2-BpbZIP1)and INVSC1(pYES2)under the same treatment conditions(P<0.01)
3 讨论
植物修复(Phytoremediation)是一种利用自然生长植物或者遗传工程培育植物修复金属污染土壤环境的技术,是一种经济、高效的方法。超级积累植物可以从土壤中吸取金属污染物以达到降低或去除土壤重金属污染的目的。目前发现的重金属超积累植物已经多达700 多种,如,我国东南景天()为Cd 和Zn 超级积累植物;土荆芥()为Pb 超级积累植物;商陆()能大量富集Cd;荨麻()对Zn有较强富集能力。可见,植物在重金属治理过程中可以发挥重要作用,发展植物修复方法可行性强。构树以其突出的经济价值和优良的生态适应性受到广泛关注。商侃侃等分析了54 种木本植物对土壤Cu、Pb、Zn 的提取能力,结果表明构树具有较强的金属离子综合提取能力。金裕华等研究表明构树在重金属污染土壤中适应性较强、生长良好。曾鹏等研究显示芦竹()与构树、桑树()间种可有效用于重金属污染土壤修复。可见,充分掌握构树的重金属响应分子机制,能为更好地应用构树于重金属污染修复提供依据。
本研究鉴定获得的基因与拟南芥bZIP的较多成员具有较近的亲缘关系(见图3),其中与的进化关系较近;是一个糖调控基因,介导糖信号,影响基因的表达和植物的生长发育。且Sun 等研究证明参与植物生物调控,发现其为植物耐盐、耐渗透和耐盐性的正向调节因子。参与植物中活性氧(ROS)的产生,而ROS 作为信号分子调控植物对生物和非生物胁迫的响应。是葡萄糖-ABA 相互作用网络的重要节点,根据糖的供应情况参与ABA介导的非生物胁迫响应调节。所以,我们初步认为基因可能与植物非生物胁迫响应相关。
为进一步明确基因是否能响应Cd胁迫,对构树进行Cd 处理,分析不同处理时间点该基因在根、茎、叶中的表达。结果发现,基因在构树根、茎、叶中都表现出对Cd 胁迫的积极响应,且具有一定的组织特异性(见图5),这与其他基因在逆境中的表达具有相似性。如,耿芳等通过实时RT-PCR 分析a 基因在烟草中的表达,在干旱、高盐的诱导下烟草()中基因表达量明显上调,后证明基因参与转基因拟南芥耐旱和耐盐的调控。同样,才华等发现在大豆的根和叶中基因的表达均受盐胁迫诱导,过表达基因的拟南芥对盐胁迫的敏感性提高。王荣凯使用NaCl 胁迫嘎啦生根组培苗,组培苗根中表达受到明显诱导,苹果()转基因研究表明是苹果抗逆的正调控因子。由此可见,基因可以响应镉胁迫,具有镉胁迫耐受调控潜力。
为进一步快速确定基因对Cd的响应功能,本研究将转入酵母,通过与对照酵母比较,分析其抗Cd 能力。结果发现,转入基因的酵母INVSC1(pYES2-BpbZIP1)在CdCl胁迫下的生长活性显著高于对照INVSC1(pYES2)(见图6)。由于酵母表达系统被广泛应用于快速检测基因的抗逆功能,如通过转入和基因提高酵母对盐、渗透及热胁迫的耐受性,证明和能够积极响应非生物胁迫;用同样的方法验证了和在非生物胁迫反应中的积极作用;Yang等通过将柽柳()金属硫蛋白在酵母中过表达,发现其能增强酵母的抗Cd、Zn、Cu和NaCl 能力,证明具有增强重金属耐受性的作用;Jiang 等研究表明基因导入酵母后,转基因酵母在热、盐和氧化应激条件下显示出更高的生存力,证明基因的抗逆能力。本研究Cd 胁迫下的酵母生活力被基因的表达而显著提高,表明的表达能有效改善转基因酵母的抗Cd性,可以作为Cd 响应的候选基因。在后续研究中,我们将通过植物表达系统对基因调控Cd胁迫的分子机制进行全面解析。
猜你喜欢
伴侣(2022年3期)2022-04-08
中国军转民(2019年9期)2019-09-10
名人传记·财富人物(2017年9期)2017-11-02
名人传记·财富人物(2017年9期)2017-11-02
中国扶贫(2017年16期)2017-08-16
中国扶贫(2017年5期)2017-03-10
中国扶贫(2016年16期)2016-12-09
Coco薇(2016年8期)2016-10-09
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
