
甘薯响应蔓割病病原菌侵染的IbWRKY7基因克隆与表达分析
2022-05-16 13:59刘泓江陈培茹杨新笋王连军柴沙沙靳晓杰杨圆圆程贤亮焦春海张文英
中国农业大学学报 2022年6期
刘 意 刘泓江 陈培茹 杨新笋 雷 剑 王连军 柴沙沙 靳晓杰 杨圆圆 程贤亮 焦春海* 张文英
(1.长江大学 农学院,湖北 荆州 434025; 2.湖北省农业科学院 粮食作物研究所,武汉 430064)
甘薯(Ipomoea
batatas
(L.)Lam.)是我国重要的粮食、饲料、工业原料和生物质能源作物,是世界第七大粮食作物。甘薯蔓割病又称镰刀菌枯萎病(Fusarium
wilt),是一种真菌性病害,是我国南方甘薯种植区的主要病害。选育和种植抗病品种是目前最经济有效且绿色环保的防治措施。探究抗病分子机理可提高抗病种质资源的利用效率。WRKY家族作为植物中一类重要的转录因子,在植物抵御逆境胁迫中起着重要的调控作用。IbSPF1
是第1个被克隆的WRKY基因,来自于普通栽培甘薯‘高系14’。目前针对水稻(Oryza
sativa
)和拟南芥(Arabidopsis
thaliana
)中的WRKY研究较多。水稻的OsWRKY22
、OsWRKY30
和OsWRKY45
均正调控水稻对稻瘟病菌(Magnaporthe
grisea
)的应答反应。OsWRKY28
和OsWRKY76
均负调控水稻对稻瘟病菌的应答反应。AtWRKY25
在水杨酸介导的紫丁香假单胞菌(Pseudomonas
syringae
)防御反应中发挥负调控作用。同样,AtWRKY38
和AtWRKY62
被证明负向调控拟南芥对紫丁香假单胞菌的抗性。相反,AtWRKY33
超表达可提高拟南芥对腐生型病原菌灰霉病菌(Botrytis
cinerea
)和黑斑病菌(Alternaria
brassicicola
)的抗性。已有研究表明WRKY转录因子在植物抵御生物胁迫中既存在正向调控又有负向调控作用。目前已报道与甘薯抗蔓割病相关的基因有IbSWEET10
和IbBBX24
。有关在甘薯抗蔓割病病原菌侵染后甘薯的WRKY转录因子表达模式研究尚未见报道。本研究以‘鄂薯11’为试验材料,克隆1个新的WRKY家族基因IbWRKY7
,进行生物信息学分析;并对甘薯不同抗病品种在病原菌侵染后该基因的表达模式进行分析,旨在探究甘薯响应蔓割病病原菌侵染过程中IbWRKY7
基因的表达模式,以期为揭示甘薯蔓割病抗性机理研究奠定基础。1 材料与方法
1.1 试验材料
‘鄂薯11’是湖北省农业科学院粮食作物研究所自主选育的高抗蔓割病甘薯品种;‘栗子香’是甘薯蔓割病敏感品种,由中国农业科学院甘薯研究所选育。
田间剪取健康的‘鄂薯11’和‘栗子香’薯苗主茎顶部约16 cm茎段,置于装有清水的三角瓶(100 mL)中生长,选取生长状态一致的健康植株,用无菌剪刀制造新的伤口,将伤口在孢子浓度为1×10个的菌液中持续浸泡。分别在侵染0、2、4、12、24、48、72、96和120 h后取茎基部(4 cm),进行液氮速冻后保存至-80 ℃超低温冰箱,每个处理3个重复。试验在(28±1) ℃光照生长室(16 h光照/8 h黑暗,光照强度为3 000 Lx)中进行。
1.2 试验方法
1
.2
.1
甘薯总RNA提取及cDNA合成使用北京天漠科技开发有限公司总RNA提取试剂盒提取‘鄂薯11’和‘栗子香’总RNA。用超微量分光光度计检测总RNA的浓度。采用TransScriptAll-in-One First-Strand cDNA Synthesis SuperMix for qPCR(One-Step gDNA Removal)试剂盒(北京全式金生物技术有限公司,中国)反转录合成cDNA第一链,-20 ℃存放,备用。
1
.2
.2
IbWRKY7
基因克隆基于有参转录组数据中筛选基因ID g1093的WRKY家族基因,Nr数据库(ftp:∥ftp.ncbi.nlm.nih.gov/blast/db/)注释结果为WRKY
7基因。利用基因ID在六倍体甘薯参考基因组数据库(https:∥ipomoea-genome.org/)中检索获得甘薯IbWRKY7
基因的CDS序列。利用Primer 5.0软件在参考序列CDS序列两端设计特异性引物即IbWRKR7-CDS-F
: ATGGCAGTGGACCTTATGATGG;IbWRKR7-
CDS-R
: CTAAGAAGACTCTAAGATTAAACTGT。利用 PCR扩增试剂盒TaKaRa La Taq(宝日医生物技术(北京)有限公司)进行PCR扩增, PCR 反应程序如下:94 ℃预变性5 min;94 ℃变性30 s,53 ℃复性30 s,72 ℃延伸1.5 min,35个循环;72 ℃延伸10 min;4 ℃保存。PCR目的产物回收、纯化后,连接到pMD19-T 载体(宝日医生物技术(北京)有限公司),转化DH-5α大肠杆菌感受态(北京全式金生物技术有限公司),挑选单克隆摇菌后,进行菌液PCR鉴定。挑选阳性单克隆送奥科鼎盛生物技术有限公司(武汉)进行测序。
1
.2
.3
IbWRKY7
基因序列的生物信息学分析利用NCBI网站(https:∥www.ncbi.nlm.nih.gov/)Blastp程序进行氨基酸序列比对查找同源蛋白;利用DNAMAN V6.0软件进行多序列比对;采用MEGA 7.0软件的Neighbor-joining法(bootstrap为1 000)构建同源序列系统进化树;利用在线软件Expasy(https:∥web.expasy.org/protparam/)网站分析氨基酸的分子量、等点电(pI)、不稳定系数和亲水性;利用CELLO(http:∥cello.life.nctu.edu.tw/)、Cell-PLoc 2.0(http:∥www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)以及PSORT II (https:∥psort.hgc.jp/form2.html)等多种在线网站完成亚细胞定位; Plant CARE(http:∥bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线工具分析启动子的顺式作用元件,并利用GSDS 2.0(http:∥gsds.gao-lab.org/index.php)在线工具绘图。
1
.2
.4
IbWRKY7
基因表达分析利用BIO-RAD CFX96实时荧光定量PCR仪(伯乐生命医学产品(上海)有限公司)对甘薯进行实时荧光定量PCR分析。利用Primer 5.0软件设计IbWRKY7
荧光定量特异性引物,引物序列分别为IbWRKY7-F
: CCCGTCGTTTCTTCTTTGC、IbWRKY7-R
: GAGGCGGAACTTGCTGAATC,以甘薯β肌动蛋白基因为内参基因,β
-Actin
的扩增引物为β
-Actin
-F
:AGCAGCATGAAGATTAAGGTTGTAGCAC、β
-Actin
-R
:TGGAAAATTAGAAGCACTTCCTGTGAAC。利用PerfectStartGreen qPCR SuperMix试剂盒(北京全式金生物技术有限公司)进行PCR扩增,每个反应3次重复。荧光定量PCR反应程序如下:94 ℃预变性30 s,94 ℃变性5 s,58 S退火30 s,40个循环;溶解曲线设置为65 ℃ 5 s到95 ℃,增量为0.5 ℃/循环。1
.2
.5
数据分析利用2法计算IbWRKY7
基因相对表达量并使用DPS 7.5统计软件对‘鄂薯11’和‘栗子香’的9个侵染时间点的茎部IbWRKY7
基因的表达量进行双因素方差分析和显著性检验,P
<0.05表示差异显著。2 结果与分析
2.1 IbWRKY7基因克隆及序列分析
以‘鄂薯11’ cDNA作为模板,利用IbWRKR7-CDS-F
和IbWRKR7-CDS-R
进行PCR扩增。由图1可知, PCR产物经1%琼脂糖凝胶电泳检测得到约900 bp的单一条带,与预期大小一致。将测序正确的CDS序列翻译成蛋白序列,并在NCBI在线数据库中对其蛋白序列进行blastp比对,与其他植物WRKY7蛋白同源性最高,这与Nr数据库注释信息吻合,因此将其命名为IbWRKY7
。由表1可知,IbWRKY7
的CDS序列长度为945 bp,编码314个氨基酸;预测蛋白分子量为33.92 kU,pI 9.82,IbWRKY7
编码不稳定的亲水性蛋白。2.2 IbWRKY7启动子的顺式作用元件分析
利用Plant CARE启动子在线分析网站对IbWRKY7
基因上游2 000 bp区域序列进行分析,序列中含有启动子的基本作用元件外,还存在多个与抗逆相关的顺式作用元件(图2),包括3个茉莉酸甲酯应答元件(CGTCA-motif、G-box和TGACG-motif),1个脱落酸应答元件(ABRE),1个生长素应答元件(TGA-element),1个防御和应激反应元件(TC-rich),2个胁迫响应元件(MYB和MYC)等(表2),说明该基因可能与甘薯的抗逆性相关。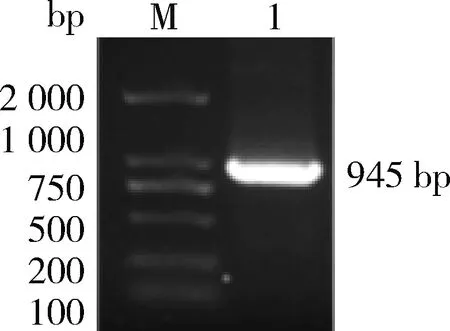
M,DNA标准DL 2 000;1,IbWRKY7基因的扩增产物。 M, DL 2 000 DNA Marker; 1, Products of IbWRKY7 gene.图1 甘薯IbWRKY7基因的克隆Fig.1 Cloning of IbWRKY7 gene from sweet potato
表1 甘薯基因序列分析
Table 1 Sequence analysis of sweet potato
基因名称Gene name长度/bpLength氨基酸数目Number ofamino acid蛋白分子量/kUProtein molecularweight等电点pI不稳定系数Instabilityindex亲水性值Grand average ofhydropathicityIbWRKY794531433.929.8246.04-0.461

图2 IbWRKY7启动子区的顺式作用元件分布Fig.2 Distribution of IbWRKY7 promoter cis-acting elements
表2 基因启动子抗逆相关顺式作用元件
Table 2 -acting elements related to stress tolerance detected in the promoter region of
编号Code元件Element核心序列Core sequence数量Number功能Function1TC-rich repeatsATTCTCTAAC2防御和胁迫应答元件Cis-acting element involved in defense and stress responsiveness2W-boxTTGACC4激素信号转导Hormone signal transduction3CGTCA-motifCGTCA1茉莉酸甲酯应答元件Cis-acting regulatory element involved in the MeJA-responsiveness4G-boxCACGTG1茉莉酸甲酯应答元件Cis-acting regulatory element involved in the MeJA-responsiveness5TGACG-motifTGACG1茉莉酸甲酯应答元件Cis-acting regulatory element involved in the MeJA-responsiveness6TGA-elementAACGAC1生长素应答元件Cis-acting regulatory element involved in auxin responsiveness7ABREACGTG2脱落酸应答元件Cis-acting element involved in salicylic acid responsiveness8MYCCATTTG6胁迫响应元件Cis-acting element involved in stress responsiveness
表2(续)
编号Code元件Element核心序列Core sequence数量Number功能Function9MYBCAACCA5胁迫响应元件Cis-acting element involved in stress responsiveness10AREAAACCA3厌氧诱导顺式调节元件Cis-acting regulatory element essential for the anaerobic induction11as-1TGACG1植物激素应答元件Cis-acting element involved in hormone responsiveness12MybTAACTG2植物激素应答元件Cis-acting element involved in hormone responsiveness
2.3 IbWRKY7蛋白的相似性及进化分析
由图3可知,与其他植物的WRKY7蛋白进行多重序列比对表明,10个WRKY7蛋白均有WRKY家族共有特点,即包含1个WRKY七肽保守序列(WRKYGQK)和1个CHC(C-X-C-X-H-X-C)的锌指结构。IbWRKY7与其他9个WRKY7氨基酸序列具有相似性。由图4可知,甘薯IbWRKY7与日本牵牛花InWRKY7和三裂叶薯ItWRKY7亲缘关系最近,与葡萄VvWRKY7、苹果MdWRKY7以及樱桃PaWRKY7等亲缘关系较远。

Ib,甘薯; It,三裂叶薯(XP_031117517.1);St,马铃薯(XP_006339860.1);In,日本牵牛花(XP_019183704.1);Ca,辣椒(XP_016561025.1);Sp,野生种番茄(XP_015063634.1);Nt,烟草(NP_001312942.1);Pa,樱桃(XP_021802303.1);Md,苹果(XP_008384448.2); Vv,葡萄(XP_002283219.1)。下同。红色框代表锌指结构域,蓝色框代表WRKY结构域。 Ib: Ipomoea batatas (L.) Lam; It: Ipomoea triloba (XP_031117517.1); St: Solanum tuberosum (XP_006339860.1); In: Ipomoea nil (XP_019183704.1); Ca: Capsicum annuum (XP_016561025.1); Sp: Solanum pennellii (XP_015063634.1); Nt: Nicotiana tabacum (NP_001312942.1); Pa: Prunus avium (XP_021802303.1); Md: Malus domestica (XP_008384448.2); Vv: Vitis vinifera (XP_002283219.1). The same below. The red box represents the zinc finger domain. The blue box represents the WRKY domain.图3 IbWRKY7氨基酸序列与其他植物 WRKY7蛋白的多序列比对Fig.3 Multi-sequence alignment of the IbWRKY7 amino acid sequences with those in other plants

图4 不同植物WRKY7蛋白系统发育分析Fig.4 Phylogenetic analysis of WRKY7 proteins from different plants
2.4 IbWRKY7蛋白的亚细胞定位
由表3可知,利用CELLO和PSORT II预测结果均显示IbWRKY7蛋白定位于细胞核的预测分值高于其他位置,同时Cell-PLoc 2.0预测亚细胞定位结果表明IbWRKY7蛋白定位于细胞核。综合3种在线预测软件分析结果表明IbWRKY7蛋白定位于细胞核。
2.5 IbWRKY7基因的表达特性分析
由图5可知,甘薯蔓割病菌(Fob)侵染甘薯茎部后,IbWRKY7
基因表达量在抗性不同的品种之间差异显著。在Fob侵染后,‘鄂薯11’的IbWRKY7
表达量在4 h达到最高随后下降,2、4、12、24、48、72、96和120 h的表达量均显著高于未侵染(0 h)时的表达量,而‘栗子香’的IbWRKY7
表达量在侵染96 h才达到最高,除侵染24 h外其他7个时间点的表达量均显著高于0 h。处理0 h,‘鄂薯11’中IbWRKY7
表达量高于‘栗子香’但不显著。Fob侵染2、4、12、24和48 h时,‘鄂薯11’IbWRKY7
基因表达量显著高于‘栗子香’,而72和96 h则相反,120 h时,两者的IbWRKY7
基因表达量无显著性差异。表3 IbWRKY7蛋白亚细胞定位
Table 3 Subcellular location prediction of IbWRKY7 protein
软件Software细胞质Chloroplasm细胞核Nuclear线粒体Mitochondrial细胞骨架CytoskeletalCELLO0.1241.2910.8010.010PSORT II30.45%52.20%8.70%8.70%
注:数值越大代表IbWRKY7蛋白定位在该位置可能性越大。
Note: The higher the value, the more likely the IbWRKY7 protein will be located at this location.
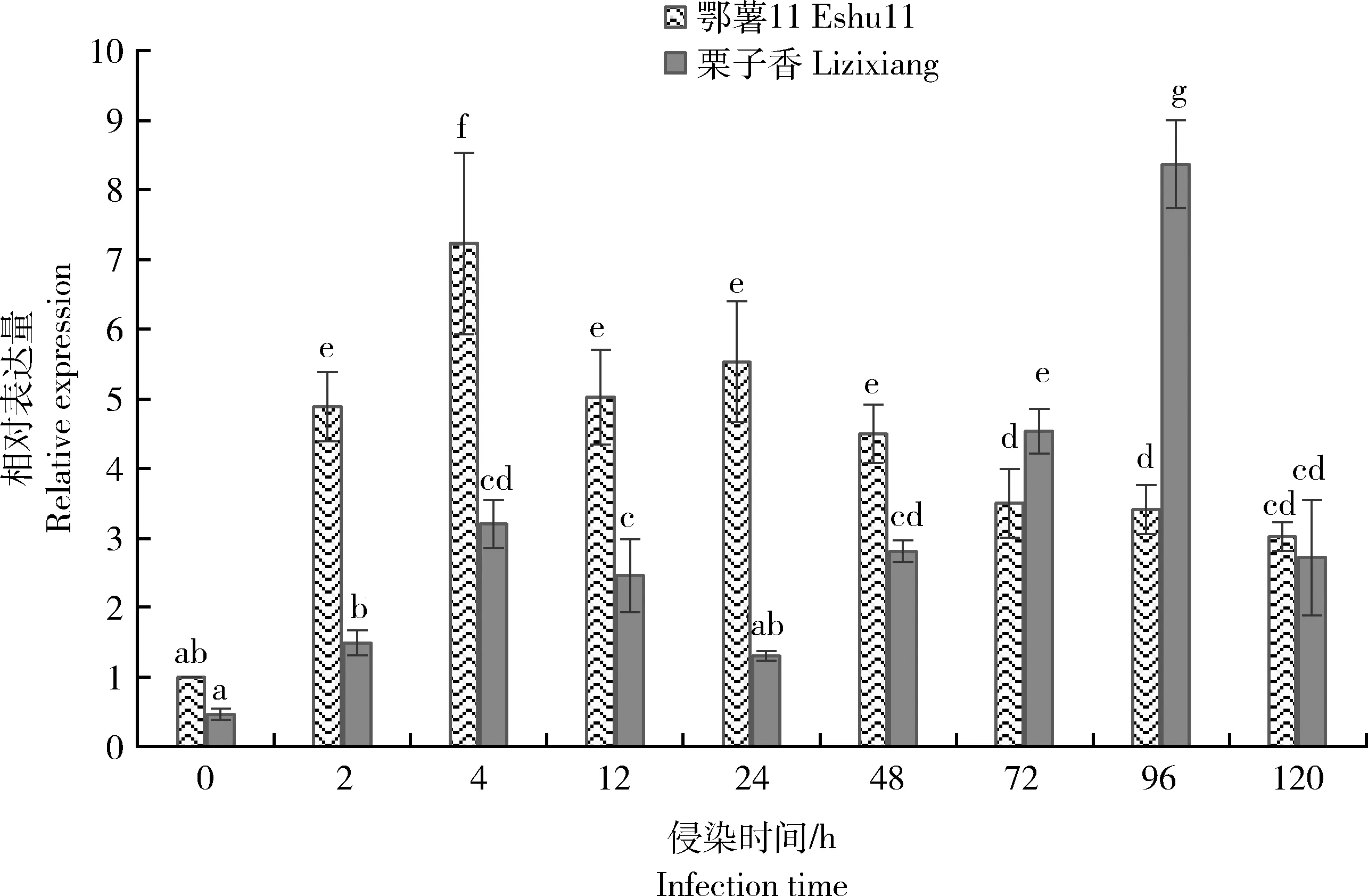
不同小写字母表示表达量差异显著(P<0.05)。 Different lowercase letters indicate significant differences (P<0.05) in IbWRKY7 expression levels.图5 蔓割病病原菌侵染后IbWRKY7的相对表达量Fig.5 Relative expression level of IbWRKY7 after Fusarium oxysporum f. sp. batatas infection
3 讨 论
WRKY家族转录因子作为植物中一类重要的转录因子,在植物抵御生物逆境胁迫与非生物逆境胁迫中起着重要的调控作用。CmWRKY15
基因通过影响水杨酸信号途径正向调控菊花(Chrysanthemum
morifolium
)对柄锈菌(Puccinia
horiana
Henn)的抗性。过量表达MdWRKY100
的转基因苹果(Malus
domestica
)植株对炭疽病菌(Colletotrichum
gloeosporioides
)的抗性增强,而RNAi植株对炭疽病菌更为敏感,结果表明MdWRKY100
正向调控苹果植株对炭疽病菌的抗性。PlWRKY65
基因通过调节茉莉酸和水杨酸信号转导正向调控芍药(Paeonia
lactiflora
)对细链孢霉(Alternaria
tenuissima
)的抗性。沉默GmWRKY40
的转基因大豆(Glycine
max
)毛状根对大豆疫霉菌(Phytophthora
sojae
)侵染的敏感性增强。Liu等研究发现RcWRKY41
基因沉默植株对灰霉病(Botrytis
cinerea
)的抗性降低。目前,关于甘薯中WRKY家族基因的研究较少。王连军等在高抗根腐病甘薯品种‘徐薯18’中克隆到15个WRKY基因家族序列。朱虹等研究表明IbWRKY2
基因正向调控转基因拟南芥的耐旱性和耐盐性。IbWRKY75
基因可能参与甘薯的抗旱、耐盐过程以及正调控甘薯叶片的衰老。张凯等利用酵母双杂交技术从甘薯文库筛选获得甘薯IbWRKY61的互作蛋白主要与生物胁迫响应相关,推测IbWRKY61
基因主要参与甘薯病原菌侵染的防御。甘薯WRKY家族基因功能研究较少,仍有待进一步挖掘和功能验证。依据WRKY七肽保守序列数量和锌指结构特点将WRKY蛋白分为3类:第Ⅰ类WRKY蛋白含有2个保守的WRKY七肽保守序列(WRKYGQK)和1个CH(C-X-C-X-H-X-H)的锌指结构;第Ⅱ类WRKY蛋白含有1个保守的WRKY七肽保守序列和1个CH的锌指结构;第Ⅲ类WRKY蛋白含有1个WRKY 七肽保守序列和1个的CHC(C-X-C-X-H-X-C)锌指结构,大部分WRKY家族成员蛋白属于第Ⅱ类WRKY蛋白。本研究在高抗蔓割病甘薯品种‘鄂薯11’中克隆了IbWRKY7
基因。对IbWRKY7蛋白序列进行生物信息学分析结果表明IbWRKY7蛋白与其他几个物种WRKY7蛋白有较高相似性,且具有相同的保守结构域(1个WRKY保守结构域和1个CHC型锌指结构域),因此IbWRKY7属于第III类WRKY蛋白。综上,本研究克隆得到的基因确为WRKY
基因家族成员IbWRKY7
。IbWRKY7
基因上游启动子区域含有多个激素应答和逆境胁迫应答相关的元件,如MYC、MYB、G-box、TGACG-motif以及ABRE等,这表明IbWRKY7
基因可能参与甘薯响应逆境胁迫的调控。沉默AtWRKY7
的拟南芥植株对灰霉病菌的敏感性显著提高,而过表达AtWRKY7
基因提高了拟南芥对灰霉病菌的抗性,表明AtWRKY7
基因正向调控拟南芥对灰霉病菌的抗性。相反,AtWRKY7
基因负向调控拟南芥对细菌类病原菌紫丁香假单胞菌抗性。郑超研究表明OsWRKY7
基因可能负向调控水稻对白叶枯病菌(Xanthomonas
oryzae
)免疫应答,降低水稻植株对白叶枯病的抗性。拟南芥中异源表达大白菜(Brassica
rapa
L. ssp.pekinensis
)BrWRKY7
基因可提高拟南芥对腐生型病原菌软腐果胶杆菌(Pectobacterium
carotovorumb
ssp.
carotovorum
)的抗性。以上研究表明WRKY7
基因在植物应答病原菌中发挥重要作用。本研究利用蔓割病病原菌侵染感病品种‘栗子香’和抗病品种‘鄂薯11’,并对IbWRKY7
基因的表达模式进行了分析。结果显示,病原菌侵染后,2个品种IbWRKY7
基因均显著上调,‘鄂薯11’中的表达量在4 h达到最大值而‘栗子香’在侵染96 h才达到最大值,并且侵染2、4、12、24和48 h,‘鄂薯11’中的IbWRKY7
基因表达量显著高于‘栗子香’。因此侵染前期IbWRKY7
基因在抗病品种中的表达量更高,表明‘鄂薯11’中IbWRKY7
基因更快速响应蔓割病病原菌的侵染。IbWRKY7
基因可能在甘薯早期响应蔓割病病原菌侵染过程中起重要作用,有待进一步利用转基因植株进行功能验证。4 结 论
本研究表明甘薯IbWRKY7
基因上游2 000 bp的区域含有12种激素应答和胁迫应答相关的顺式作用元件。IbWRKY7
的CDS序列全长为945 bp,编码314个氨基酸,预测蛋白分子量为33.92 kU,pI 9.82,编码不稳定的亲水性蛋白,具有WRKY家族的典型结构特征即1个WRKY七肽保守序列和1个CHC型锌指结构,属于第Ⅲ类WRKY蛋白,氨基酸序列与同源物种相似度高,进化上高度保守。IbWRKY7
基因不仅受Fob侵染诱导表达而且在不同蔓割病抗性品种中表达量差异显著。猜你喜欢
中国瓜菜(2019年8期)2019-09-19
农民致富之友(2019年23期)2019-08-16
湖北农业科学(2017年4期)2017-03-28
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
世界热带农业信息(2016年3期)2016-04-05
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
小学生导刊(中年级)(2009年6期)2009-11-10
