
酵母培养物对初产母猪哺乳期血浆生化指标和代谢组的影响
2022-05-12 10:13孟庆维马智卓李建平
动物营养学报 2022年4期
吴 泽 陈 鹏 孟庆维 马智卓 李建平*
(1.东北农业大学动物科学技术学院,哈尔滨 150030;2.北京英惠尔生物技术有限公司,北京 101109)
在猪生产体系中,初产母猪的营养调控是十分关键的环节,初产母猪在哺乳期的营养水平和健康状态对母猪的繁殖性能、利用年限及其后代的生长性能有很深远的影响[1-2]。如果初产母猪哺乳期的营养摄入不能满足其营养需求,初产母猪则会在哺乳期间分解身体储存以补充其营养消耗,而身体储存损失会导致母猪下次受孕时卵泡营养不良和黄体发育受阻,从而降低胚胎存活率[3]。因此,初产母猪在哺乳期间的代谢水平将影响其未来的健康和繁殖性能。
母猪机体代谢是一个复杂的过程,除了需要体内多个器官协同发挥作用,体内的糖、脂类和蛋白质也相互影响,并在动物体内分解成小分子物质共同维持机体正常的生命活动[4]。母猪分娩后由于泌乳需要、机体生理变化和应激反应容易发生能量负平衡[5]。因此,调节哺乳母猪能量代谢平衡对母猪健康水平至关重要。酵母培养物(yeast culture,YC)是一种在特定的温度条件下以酵母为原料的干发酵产品,它是在未经修饰的酿酒酵母菌株发酵过程中产生的[6]。YC已广泛应用于调控猪的营养与健康,在促进家畜的健康和生产性能方面是较理想的抗生素替代品[7]。研究显示,YC可以降低母猪哺乳期血浆中尿素氮和胆固醇的含量,减少脂类分解,提高氮的利用率,表明YC有改善哺乳母猪代谢平衡的作用[8]。但目前关于YC对初产母猪哺乳期标志代谢物及其参与代谢通路的研究鲜有报道。因此,本试验利用代谢组学手段,通过有监督的主成分分析(PCA)和偏最小二乘法判别分析(PLS-DA)等方法,筛选、鉴定初产母猪哺乳期血浆中受到YC影响的代谢物及其代谢途径,以期为更深入了解YC调节初产母猪哺乳期代谢的主要标志代谢物变化和生理机制提供理论依据。
1 材料与方法
1.1 试验材料
试验用YC粗蛋白质含量≥15%、甘露聚糖含量≥1%。
1.2 试验设计
选用健康的初产母猪(长白)60头,随机分为2组,每组30个重复,每个重复1头猪。对照组饲喂基础饲粮,试验组在基础饲粮中添加YC(妊娠期添加0.5%,哺乳期添加0.8%)。饲养试验从母猪妊娠第30天开始至分娩后第28天(仔猪断奶)结束,试验在黑龙江省巴彦县大东北牧业集团进行。母猪妊娠第30~109天饲喂妊娠期饲粮,从妊娠第110天开始至试验结束饲喂哺乳期饲粮。基础饲粮参照NRC(2012)母猪营养需要量进行配制,其组成及营养水平见表1。

表1 基础饲粮组成及营养水平(风干基础)
1.3 饲养管理
试验母猪在妊娠第30~109天在妊娠舍饲养,每天08:00和14:00各饲喂1次妊娠期饲粮。妊娠第110天,母猪被转移至哺乳舍待产并开始饲喂哺乳期饲粮。分娩后,母猪每天饲喂4次(05:00、10:00、16:00、20:00)哺乳期饲粮,饲喂初始量为每日1 kg,以后每天增加1 kg直至达到母猪最大采食量后自由采食。妊娠期和哺乳期母猪均在限位栏饲养且不限饮水。妊娠舍温度保持在18~20 ℃,哺乳舍温度保持在20~23 ℃。试验期间保持舍内通风、干燥,消毒和防疫严格按照猪场防疫措施和饲养管理制度进行。
1.4 样品采集
母猪分娩后第28天(仔猪断奶),采集耳缘静脉血10 mL于抗凝的采血管中,3 000 r/min离心15 min后收集上层血浆,置于1.5 mL EP管中,液氮速冻,-80 ℃保存待测。
1.5 测定指标与方法
1.5.1 血浆生化指标
采用全自动生化分析仪(DS-161V,英诺华公司)检测母猪血浆中谷丙转氨酶(GPT)、谷草转氨酶(GOT)活性以及总蛋白(TP)、白蛋白(ALB)、尿素氮(UN)、葡萄糖(GLU)、甘油三酯(TG)、总胆固醇(TC)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)含量。
1.5.2 血浆代谢组学分析
血浆样本在4 ℃环境下缓慢解冻后,取适量样本加入预冷甲醇/乙腈/水溶液(2∶2∶1,体积比),涡旋混合,低温超声30 min,-20 ℃静置10 min,14 000×g4 ℃离心20 min,取上清真空干燥,质谱分析时加入100 μL乙腈水溶液(乙腈∶水=1∶1,体积比)复溶,涡旋,14 000×g4 ℃离心15 min,取上清液转入自动进样瓶。采用Agilent 1290 Infinity LC超高效液相色谱系统(UHPLC)HILIC色谱柱进行分离;为避免仪器检测信号波动而造成的影响,采用随机顺序进行样本的连续分析。样本队列中插入QC样品(取等量制备好的试验样本混合而成),用于监测和评价系统的稳定性及数据的可靠性。采用AB Triple TOF 6600质谱仪进行样本一级、二级谱图的采集。采用XCMS软件进行峰对齐、保留时间校正和提取峰面积。对XCMS提取得到的数据首先进行代谢物结构鉴定、数据预处理,然后进行数据质量评价,最后再进行数据分析。采用有监督的PLS-DA方法对样品进行模式识别分析。潜在标志物根据变量投影重要度(VIP)、置信区间得分图(scoring plot)和载荷图(loading plot)进行筛选,用SPSS 25.0软件进行独立样品t检验分析,再根据VIP和统计结果进行判别分析。
1.6 数据处理与统计分析
血浆生化指标等数据用Excel 2 010进行初步处理后再采用SPSS 25.0软件通过独立样品t检验方法分析组间差异显著性,P<0.05表示差异显著。
2 结果与分析
2.1 饲粮中添加YC对初产母猪哺乳期血浆生化指标的影响
由表2可知,与对照组相比,饲粮中添加YC显著降低了母猪血浆中葡萄糖、尿素氮、总胆固醇和低密度脂蛋白的含量(P<0.05),对其他血浆生化指标无显著影响(P>0.05)。
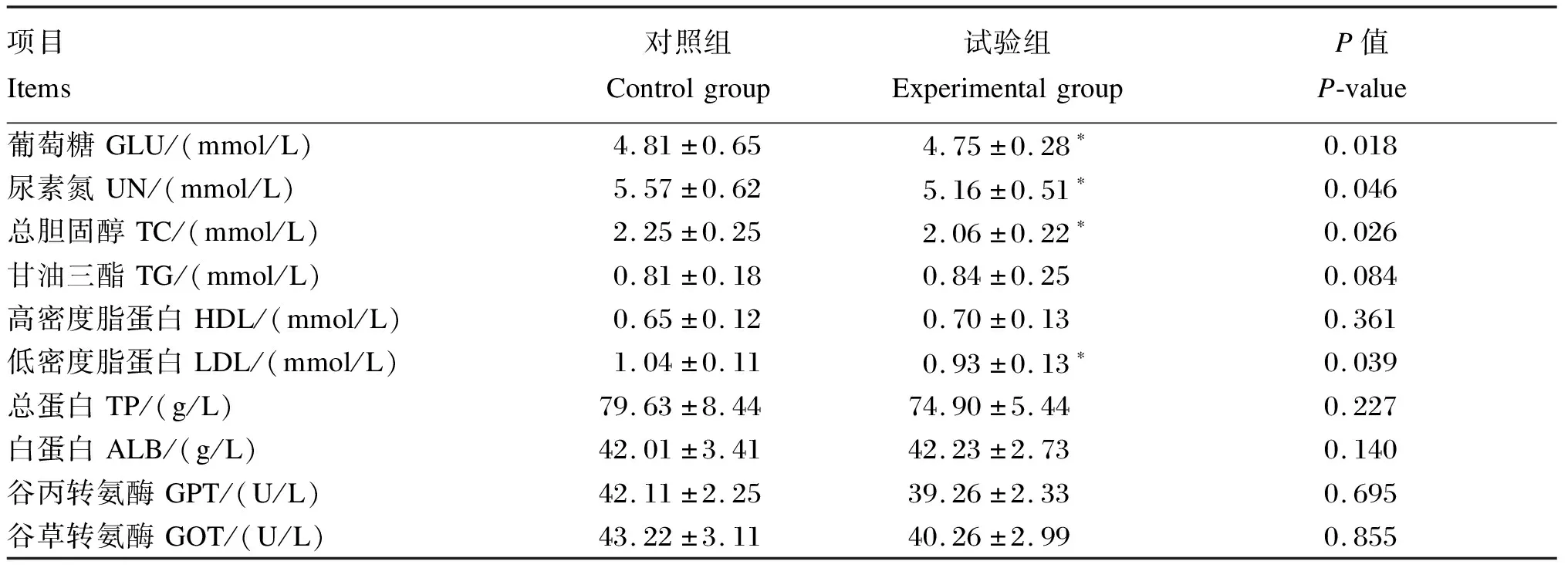
表2 饲粮中添加YC对初产母猪哺乳期血浆生化指标的影响
2.2 饲粮中添加YC对初产母猪哺乳期血浆代谢组的影响
血浆代谢组学数据分析结果表明,正离子模式下在母猪血浆中检测到142种可鉴定出的代谢物,负离子模式下共鉴定出141种代谢物。
2.2.1 总体样本PCA
将所有样本和QC样本提取得到的峰进行PCA,如图1-a和图1-b所示。结果显示,正、负离子模式下QC样本紧密聚集在一起,说明本次研究使用的仪器稳定,重复性好。
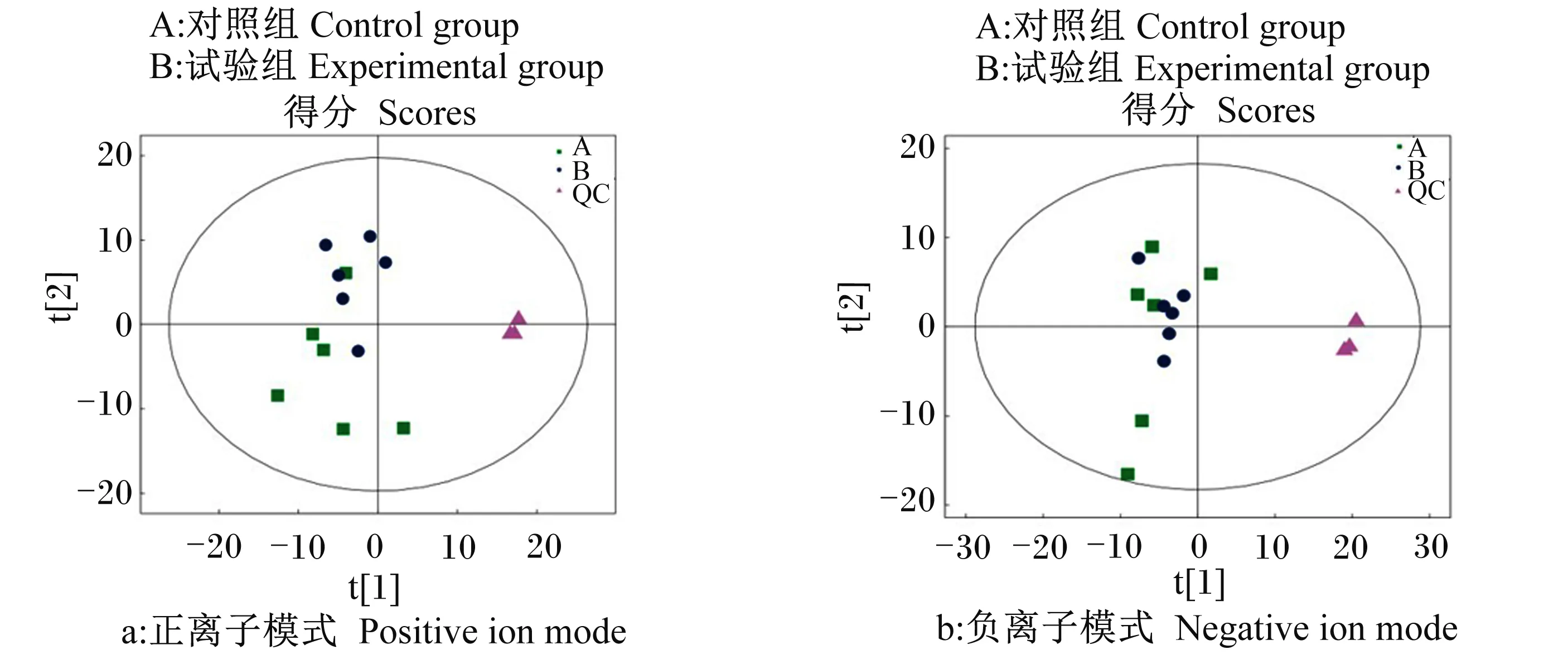
图中t[1]代表主成分1,t[2]代表主成分2,椭圆代表95%置信区间。同一颜色的点表示组内的各个生物学重复,点的分布状态反映出的是组间、组内的差异度。图2同。
2.2.2 PLS-DA
使用PLS-DA模式分析对照组与试验组之间代谢物的差异(图2),结果显示,2组代谢物的分离度好,进行7次循环交互验证得到模型评价参数在正离子模式下模型对Y变量解释率R2Y为0.995,模型预测能力Q2为0.467;在负离子模式下R2Y为0.992,Q2为0.430,表明该模式下得到的差异代谢物准确、可靠。
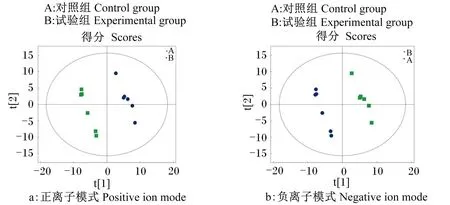
图2 初产母猪血浆代谢物偏最小二乘法判别分析
2.2.3 显著差异代谢物筛选
本试验以OPLS-DA VIP>1和P<0.05为标准筛选出差异代谢物(如表3所示)。YC显著提高了初产母猪血浆中十六烷二酸、肌酸、苯乳酸、D-脯氨酸、L-亮氨酸、L-色氨酸、L-苯丙氨酸、L-丙氨酸、L-丝氨酸、L-鸟氨酸的含量(P<0.05),显著降低了1-甲基烟酰胺和胆固醇的含量(P<0.05)。

表3 母猪血浆差异代谢物及其主要参与的代谢通路
3 讨 论
3.1 饲粮中添加YC对初产母猪哺乳期血浆生化指标的影响
动物血浆生化指标可以在一定程度上反映动物的生理代谢和营养水平。尿素氮是蛋白质和氨基酸的最终代谢产物,其含量与动物体内蛋白质或氨基酸利用率呈负相关,可以准确地表明动物体内氨基酸和蛋白质之间的代谢平衡[9]。本试验中,YC显著降低了初产母猪血浆中尿素氮的含量,这与曹迪[8]用YC在母猪上进行的研究结果一致。因此,我们推测YC可能参与调控了初产母猪哺乳期蛋白质的合成,提高了氨基酸的利用率。
动物机体血浆中葡萄糖含量一般处于一个相对稳定的状态,其升高或降低可以反映机体代谢能力的增强或减弱[10]。本研究发现,试验组初产母猪血浆中葡萄糖的含量显著降低,表明饲粮中添加YC可以减少初产母猪哺乳期机体新陈代谢,为调节哺乳母猪能量平衡发挥重要作用。
研究表明,初产母猪哺乳期脂肪动员会导致下次受孕时卵泡营养不良和黄体发育受阻,降低胚胎存活率,减少母猪利用年限[11]。母猪脂肪代谢过程十分复杂,受到体内多个器官和酶的共同调节,而总胆固醇含量是反映动物体内脂类代谢的重要指标。据报道,总胆固醇含量与脂质沉积呈正相关[12]。低密度脂蛋白和高密度脂蛋白含量可以反映动物体内总胆固醇的代谢水平,通过低密度脂蛋白可以转运肝脏合成的内源胆固醇,高密度脂蛋白可以将外周组织中的胆固醇运送到肝脏中进行代谢,产生胆汁酸排出体外,从而减少肝外组织细胞中游离胆固醇的沉积[8,13]。本试验中,YC显著降低了初产母猪血浆中总胆固醇和低密度脂蛋白含量,表明YC可能通过调节体内低密度脂蛋白含量,改善肝脏内源胆固醇合成,进而改善母猪哺乳期脂类代谢过程,减少体脂动员,提高利用年限。
3.2 饲粮中添加YC对初产母猪哺乳期血浆代谢组的影响
研究表明,初产母猪在哺乳期对营养物质的需求会不断增加,由于需要通过泌乳维持后代生长,因此容易出现能量负平衡,影响自身健康与繁殖性能[14-15]。而脂肪、蛋白质和碳水化合物等营养物质都需要在体内代谢成小分子物质才能发挥其生理功能,提供动物生命活动所需要的能量[16-17]。因此,本研究通过代谢组学技术对初产母猪血浆中的小分子代谢物进行定性和定量分析,结果共检测出12种标志性差异代谢物,揭示了YC调控初产母猪哺乳期代谢变化过程。
动物活动靠机体内的三磷酸腺苷(ATP)提供能量。烟酰胺是维生素B3的酰胺形式,也是烟酰胺腺嘌呤二核苷酸(NAD+)的前体,NAD+是氧化还原反应中ATP生产的重要辅酶,可通过无氧糖酵解和线粒体氧化磷酸化产生ATP提供能量[18-19]。肌酸在生物体内由精氨酸、蛋氨酸和甘氨酸合成,作为细胞质中的高能磷酸储备,肌酸可以快速有效地为机体提供能量[20-22]。本试验中,饲粮中添加YC显著增加了初产母猪血浆中烟酰胺和肌酸的含量,通过合成ATP为机体提供能量,为维持能量稳定提供保障[22]。
蛋白质代谢的最终途径是三羧酸循环,完全氧化并释放能量用于生产。本试验中,饲粮中添加YC显著提高了初产母猪血浆中L-苯丙氨酸、L-色氨酸、L-亮氨酸和D-脯氨酸、苯乳酸的含量。L-亮氨酸分解代谢的终产物是乙酰乙酸和乙酰辅酶A,可进入三羧酸循环合成酮,是肌肉骨骼段能量代谢的重要底物[23]。L-苯丙氨酸、苯乳酸、L-色氨酸的最终产物进入三羧酸循环生成酮和糖原,脯氨酸可以直接转化为丙酮酸作为外周组织的能量来源,保证能量稳态[24-26]。此外,L-亮氨酸的含量又与低密度脂蛋白和总胆固醇的含量呈负相关,本试验中,饲粮中添加YC降低了母猪血浆中低密度脂蛋白和总胆固醇的含量,减少了母猪体内脂肪分解[27-28]。鸟氨酸和脯氨酸是精氨酸的代谢产物,同时也是胎盘和胎液合成多胺的主要前体物[29-30],可促进卵巢固醇类物质生成,对提高动物繁殖性能具有重要作用[31]。此外,鸟氨酸的积累也可促进鸟氨酸转氨酶诱导δ’-吡咯-5-羧酸(P5C)从线粒体转运到细胞质并被P5C受体还原为脯氨酸[32]。P5C可激活红细胞衍生核因子2(Nrf2)以及相关脂肪氧化基因,促进脂肪动员[33]。因此P5C可以视作一个连接氨基酸代谢和脂肪代谢的重要桥梁。我们推测YC加速了母猪体内精氨酸代谢,进而提高鸟氨酸和脯氨酸的含量并加速了P5C向脯氨酸的还原,提高母猪繁殖性能并减少哺乳期脂肪动员。这可能是本试验中试验组初产母猪血浆中十六烷二酸含量升高的原因。而降低初产母猪哺乳期脂肪动员或脂类分解,可以促进卵泡发育并提高母猪繁殖性能[34]。因此,试验组血浆中升高的L-亮氨酸、肌酸和D-脯氨酸可能参与了抑制初产母猪部分脂肪分解,并改善了繁殖性能。
4 结 论
饲粮中添加YC可以引起初产母猪哺乳期血浆小分子代谢物含量发生显著变化,这些代谢物主要参与亮氨酸、脯氨酸等氨基酸代谢通路和脂肪酸代谢通路,其中,L-亮氨酸、D-脯氨酸、肌酸可能是YC参与调控初产母猪哺乳期代谢的重要生物标志物。
猜你喜欢
中老年保健(2022年3期)2022-11-21
草业科学(2022年9期)2022-10-21
动物营养学报(2022年9期)2022-10-12
中山大学学报(自然科学版)(中英文)(2022年4期)2022-08-05
中国饲料(2022年3期)2022-03-19
现代农村科技(2022年1期)2022-01-21
安徽化工(2021年3期)2021-05-29
三农资讯半月报(2020年19期)2020-10-27
现代农业科技(2020年10期)2020-06-04
科学与财富(2019年15期)2019-10-21
