
调节膀胱逼尿肌舒缩相关信号通路的研究
2022-05-09 01:57:50孙红伟常晓丽陈少宗
当代医学 2022年13期
孙红伟,常晓丽,陈少宗
(山东中医药大学针灸研究所,山东 济南 250355)
膀胱功能障碍可分为高张力性膀胱和低(失)张力性膀胱两大类。患者不仅会出现尿频、尿急、排尿费力、排尿踌躇等症状,严重者还会引发上尿路、肾脏病变,导致肾功能衰竭,甚至危及生命。因此,治疗膀胱功能障碍已成为临床急需解决的问题。目前,膀胱功能障碍的相关研究较多,但机制尚不清晰。本文阐述的膀胱功能障碍主要包括膀胱过度活动症(overactive bladder syndrome,OAB)和急性尿潴留(Acute urinary retention,AUR)两种常见疾病。OAB 由于膀胱逼尿肌过度收缩引起的。而AUR 与OAB 相反,AUR 会造成膀胱逼尿肌无力,收缩功能异常,两者均与膀胱逼尿肌收缩性相关。本文对OAB与AUR后的膀胱逼尿肌收缩相关的信号通路Ca2+敏感Ras 同族体基因家族成员A(Ras homolog gene family member A,RhoA)/Rho 激酶(Rho-associated kinase,ROCK)途径和p38丝裂原活化蛋白激酶(p38 mitogen-activated protein kinases,p38 MAPK)/丝裂原活化蛋白激酶激活蛋白激酶2(mitogen-activated protein kinase activated protein kinase 2,MAPKAPK2/MK2)/热休克蛋白 27(heat shock protein 27,HSP27)信号通路进行综述,以期为临床治疗膀胱功能障碍寻找新方向。
1 OAB和AUR与膀胱逼尿肌收缩性
OAB表现为膀胱在控制排尿方面存在问题,如出现尿急、尿频、夜尿或急迫性尿失禁。尤其是尿急,是膀胱过度活动症的核心症状[1],OAB主要是膀胱逼尿肌过度活动引起的。由此可知,OAB的发生与膀胱逼尿肌能否正常舒缩密切相关,已有研究表明,Ca2+敏感的RhoA/Rho激酶信号通路在调节大鼠和人类逼尿肌舒缩中起显著作用[2,3]。
AUR是泌尿外科最常见的急症之一,会造成膀胱功能的损害,导致膀胱收缩无力(detrusor underactivity,DU)发病率高,治疗效果较差[4]。尿潴留很大程度上取决于膀胱平滑肌能否正常收缩,平滑肌不能收缩,尿液则无法排出。因此,研究膀胱逼尿肌的收缩与舒张对于AUR很有必要。已有文献[5]报道,HSP27能调节AUR后的膀胱平滑肌收缩,HSP27通过p38 MAPK/MK2/HSP27 信号通路在膀胱平滑肌舒缩中起作用。
2 RhoA/Rho激酶信号通路调节膀胱平滑肌收缩机制
已有文献[4,5]报道,Ca2+敏感的RhoA/Rho 激酶信号通路在调节膀胱平滑肌舒缩中起显著作用。RhoA/Rho 激酶途径在人体内广泛存在,通过调节肌动蛋白骨架的解离及聚合状态,参与多种细胞的功能[5]。
2.1 RhoA/Rho 激酶 RhoA 是一种小分子鸟甘酸结合蛋白,可调节应激纤维形成中肌动蛋白细胞骨架,属于Ras蛋白超家族。RhoA在进化过程中非常保守,是一种胞内蛋白,通过胞外信号的刺激传递到胞内来发挥其功能,RhoA作为分子开关,可关闭和开启各种细胞内信号传导途径,如激活Cdc42 相关酪氨酸激酶(activated Cdc42-associated tyrosine kinase,ACK)、丝氨酸/苏氨酸蛋白激酶(p21-activated protein kinase,PAK)、ROCK。RhoA 绑定三磷酸鸟苷(guanosine triphosphate,GTP)时处于活化状态,也就是所谓的“开启”状态,而RhoA 绑定二磷酸鸟苷(guanosine diphosphate,GDP)时则处于非活化状态,即所谓的“关闭”状态。RhoA通过与GTP和GDP结合时所呈现的状态来调控Rho 激酶的活性。RhoA和GTP结合时与Rho激酶结合并且使其活化[6,7]。
ROCK也称Rho 相关激酶可提升肌球蛋白轻链(myosin light chain,MLC)的磷酸化水平,增加肌动-肌球蛋白的收缩力,促使细胞在细胞外基质(extracellular matrix,ECM)中迁移。丝氨酸/苏氨酸(serine/threoine,Ser/Thr)蛋白激酶与RhoA-GTP结合使其活化。ROCK是目前功能研究最为详细的Rho下游靶效应分子,包括ROCK1和ROCK2两个成员[8],且其不仅氨基酸序列的同源性很高,而且激酶区域有高达92%的同源性[9],因此,称为RhoA/Rho激酶。
2.2 肌球蛋白轻链磷酸酶 肌球蛋白轻链磷酸酶(myosin light chain phosphatase,MLCP)是 RhoA/Rho激酶下游最重要的效应物之一,包含3个亚基:催化亚基、结合亚基、功能未知的小亚基[9]。MLCP的结合亚基能与催化亚基或肌球蛋白轻链(myosin light chain,MLC)的结合域直接结合,并且能引导催化亚基和 MLC 连接,使 MLC 脱磷酸化[10]。RhoA/Rho激酶对MLCP的结合亚基进行磷酸化,使MLCP失去活性。进而抑制胞浆内MLC 脱磷酸化,提高MLC的磷酸化水平,从而增加与肌动蛋白丝交联产生的收缩力,最终引起平滑肌的收缩[11]。
2.3 MLC 与膀胱平滑肌的舒缩 已有研究证明,平滑肌收缩程度取决于MLC 的磷酸化水平,而MLC磷酸化受Ca2+依赖的肌球蛋白轻链激酶(myosin light chain kinase,MLCK)和Ca2+非依赖的MLCP双重调节[12]。MLCK能使下游效应物磷酸化,而MLCP 能使下游效应物多磷酸化。因此,由MLCK引起的膀胱平滑肌舒缩是随着细胞质中Ca2+浓度不断升高,Ca2+能与钙调蛋白(calmodulin,CaM)结合,进而形成Ca2+-CaM 复合物,激活MLCK,被活化的MLCK使MLC磷酸化,从而增加与肌动蛋白丝交联产生的收缩力,最终使平滑肌收缩。由MLCP 引起的膀胱平滑肌舒缩是RhoA/Rho激酶对MLCP的结合亚基进行磷酸化,使MLCP 失去活性,进而抑制胞浆内MLC脱磷酸化,提高MLC磷酸化水平,引起平滑肌收缩[13]。
2.4 RhoA/Rho激酶信号通路在膀胱平滑肌舒缩机制RhoA/Rho激酶信号通路虽只是一个通用的信号通路,但其中还有很多可探究的地方,如经研究发现,RhoA/Rho激酶磷酸化和磷酸酶的脱磷酸化也会受其他激酶的驱动[14],如蛋白激酶G(protein kinase G,PKG)也能对RhoA 活性进行调控,而PKG 又能被环磷酸腺苷(cyclic adenosine monophosphate,cAMP)激活,因此其可能是环鸟苷酸(cyclic guanosine monophosphate,cGMP)-蛋白激酶G(protein kinase G ,PKG)-RhoA/Rho激酶途径[15]。此信号通路只研究了一个框架,还有很多细节需要深入研究。基于OAB产生的主要原因以及RhoA/Rho激酶信号通路调节膀胱逼尿肌舒缩的研究,Y27632[16](RhoA/Rho激酶抑制剂)能抑制RhoA/Rho激酶信号通路,从而使MLCP对MLC进行脱磷酸化,减少膀胱逼尿肌的收缩,且能缓解OAB的症状[17]。Ca2+敏感RhoA/Rho激酶信号通路图,见图1。
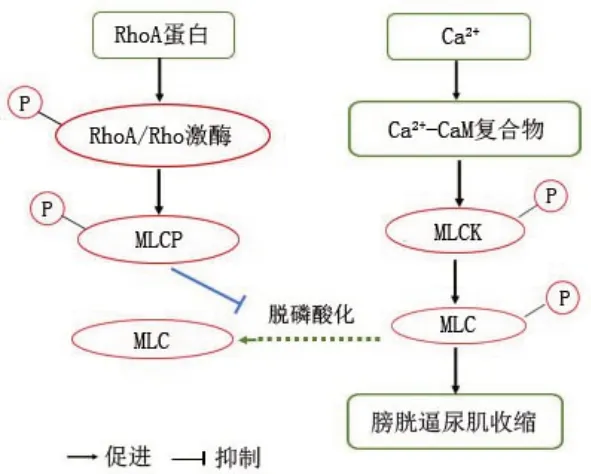
图1 RhoA/Rho激酶信号通路调节膀胱平滑肌收缩机制
3 p38 MAPK/MK2/HSP27信号通路调节膀胱平滑肌收缩机制
已有研究表明,HSP27 直接参与调控膀胱平滑肌细胞的收缩,磷酸化的HSP27 是其活化形式,并且可能通过p38 MAPK/MK2/HSP27 信号通路调节膀胱逼尿肌的收缩功能[4]。
3.1 MAPK 级联信号通路 丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPK)是普遍存在于真核生物中一类保守的丝氨酸/苏氨酸蛋白激酶,并参与许多细胞表达程序,如细胞增殖、细胞分化、细胞运动和细胞死亡。MAPK信号级联被分为三层模块。MAPK 被MAPK 激酶(MAPK-kinases,MAPKKs/MKK/MEK)磷酸化并激活,而MAPK 激酶又被MAPKK激酶(MAPKK-kinases,MAPKKKs/MKKK)磷酸化并激活。在哺乳动物中已有14 种MKKK 被鉴定 ,其中发现有 12 种 MAPK 以及 7 种MKK[18]。经分析发现,这些激酶属于不同亚族[19]。MAPK 有4 个亚族:细胞外信号调节激酶(extracellular signal-regulated kinase,ERK)、p38、c-Jun 氨基末端激酶(c-Jun N-terminal kinase,JNK)和细胞外信号调节激酶5(extracellular signal-regulated kinase 5,ERK5)。MKK 也有 4 个亚族包括 :MAPK 激酶 1(mitogen extracellular kinase 1,MEK1)、MEK2、MKK3、MKK,此级联信号通路被激活后,最后活化的MAPK会进入细胞核中,从而使许多底物蛋白的丝氨酸/苏氨酸残基磷酸化,从而修饰其活性。
3.2 MK2的激活 MK2是人体中由MK2基因编码的酶。MK2是由MAPK激活的新型蛋白激酶,并且能选择性地被p38 MAPK磷酸化[20]。
3.3 HSP27与肌动蛋白 HSP27是小分子热休克蛋白家族的成员。已有文献[21]报道,在平滑肌的收缩中,HSP27起非常重要的调节作用。人体HSP27由205个氨基酸组成,包括基因在启动子区和氨基酸序列。蛋白质磷酸化是蛋白质的一种主要修饰方式,HSP27的磷酸化位点有3个:Ser-15、Ser-78、Ser-82[22],当以上3个位点磷酸化后,大的聚合体会解聚形成小的四聚体,能够与肌动蛋白(Actin)结合,从而促进球状肌动蛋白(globular actin,G-Actin)聚合,抑制丝状肌动蛋白(filament actin,F-Actin)的解聚,进而增强平滑肌的收缩[23-26]。
3.4 p38 MAPK/MK2/HSP27 信号通路调节膀胱平滑肌收缩机制 细胞在未受到胁迫时,HSP27 表达量较低,而且主要呈大聚合体,这种聚合体下的HSP27 一般无活性。当细胞在受到胁迫时,通过G蛋白偶联受体激活MKKK,使其结合并磷酸化MKK3/6,磷酸化的MKK3/6 进一步结合并磷酸化p38 MAPK[27],磷酸化p38 MAPK进一步磷酸化下游MK2,磷酸化的MK2 进一步磷酸化HSP27,使其表达量增加以及大分子聚合体解离,使HSP27具有活性。被激活的HSP27与肌动蛋白结合,可增强平滑肌细胞收缩,从而引起膀胱逼尿肌收缩[28]。
有文献报道,SB203580 作为常用的p38 MAPK抑制剂能够通过抑制p38 MAPK/MK2/HSP27 信号通路降低平滑肌的收缩力[29-31]。膀胱逼尿肌是一种特殊的平滑肌,AUR后导致膀胱组织的过度充盈而损伤,造成逼尿肌不能正常收缩。因此,AUR 后可通过激活p38 MAPK/MK2/HSP27 信号通路来参与逼尿肌收缩和结构重建,见图2。

图2 p38 MAPK/MK2/HSP27信号通路调节膀胱平滑肌收缩机制
4 小结
膀胱功能障碍已严重影响人们的日常生活,AUR和OAB的发病率不断升高,AUR和OAB均与膀胱逼尿肌收缩性相关,因此,对于膀胱逼尿肌收缩相关的信号通路从细胞、生化与分子水平上研究很有必要。虽然临床对OAB与AUR后的膀胱逼尿肌收缩相关信号通路Ca2+敏感RhoA/Rho 激酶途径和p38 MAPK/MK2/HSP27 信号通路进行了广泛的研究,已知OAB可通过抑制Ca2+敏感hoA/Rho激酶途径,使磷酸化MLC 脱磷酸化,抑制膀胱逼尿肌收缩,促进其舒张进行缓解OAB 症状。但是,AUR后,可通过激活p38MAPK/MK2/HSP27信号通路参与逼尿肌收缩和结构重建,而此信号通路较复杂,仍存在许多未知蛋白相互作用,很多细节需需进一步深入研究。
猜你喜欢
科学(2020年2期)2020-08-24 07:57:00
中国畜牧杂志(2020年1期)2020-01-16 04:09:54
中国临床医学影像杂志(2019年4期)2019-06-18 10:55:06
浙江中西医结合杂志(2018年12期)2018-12-27 01:51:10
罕少疾病杂志(2016年5期)2016-03-11 16:34:45
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:17
医学研究杂志(2015年3期)2015-06-10 06:41:52
中国医科大学学报(2015年10期)2015-03-01 02:09:59
郑州大学学报(医学版)(2015年1期)2015-02-27 14:50:36
中国医学科学院学报(2013年6期)2013-03-11 20:26:00
