
元阳梯田不同植被类型土壤养分及酶活性特征
2022-04-30 04:02吴流通张智浩解国玲邸雪嫣苏友波
广东农业科学 2022年3期
吴流通,张智浩,解国玲,邸雪嫣,石 浩,苏友波
(云南农业大学资源与环境学院,云南 昆明 650102)
【研究意义】土壤酶是土壤中具有生物活性的蛋白质。一方面,土壤酶是土壤有机质分解、腐殖质形成、养分转化和循环的动力,在土壤物质循环和能量流动等过程中起着至关重要的作用[1-2];另一方面,土壤酶可以反映土壤微生物在物质转化过程中的所表现的活性,是评价土壤综合肥力的重要生物学指标[3]。土壤和植被有着紧密的关系,土壤在为植被生长提供必需养分的同时,植被生长又可改善土壤结构和养分状况。因此研究不同植被类型对土壤养分和酶活性的影响至关重要。【前人研究进展】土壤酶活性主要受土壤理化性质[4]、耕种方式[5]、植被类型[6]和微生物活性[7]等的影响。Chen 等[8]研究发现土地利用方式、气候环境因素、地形地貌都与土壤酶活性的变化有关。土壤养分在植物生长和有机物的腐解等过程中起着重要作用,影响着土壤生态系统中能量流动和物质循环,是反映土壤质量和土壤健康的重要指标[9]。不同植被类型不仅影响生态系统的组成方式和功能,还与植物群落组成、土壤养分库的稳定密切相关[10-11]。王雅等[12]在对黄土高原不同植被类型的研究中发现土壤养分含量和土壤酶活性均随土层深度增加而降低,且发现土壤养分间、土壤酶活性间存在显著相关性。杨秉珣等[13]在对嘉陵江流域不同植被类型的研究中还发现不同植被多样性指数与土壤酶活性、土壤养分含量呈正相关关系。Yang 等[14]研究表明土壤含水量、植物覆盖率、粘土、淤泥和废弃物养分含量与土壤养分含量呈正相关。【本研究切入点】有着1 300 多年历史的元阳梯田,是一个人与自然高度和谐的良性循环生态系统。千百年来不施用化肥却仍然有着旺盛的生命力,它不仅是传统梯田生态系统的典型代表,更是深入认识农业土壤肥力可持续发展的天然窗口。目前关于元阳梯田生态系统的研究多集中在土壤养分分布特征[15-17]、土壤微生物特征[18-19]和土壤碳氮转化[20]等,而关于不同植被类型对元阳梯田土壤养分和土壤酶活性影响的研究还较少。因此本试验以元阳梯田不同植被类型为研究对象,分析不同植被类型对土壤养分和土壤酶活性的影响。【拟解决的关键问题】通过对比不同植被类型土壤养分和土壤酶活性的差异,评价不同植被类型对土壤养分和土壤酶活性的影响,以期探明元阳梯田土壤肥力自我维持机制,为元阳梯田生态系统的保护提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于云南省红河哈尼彝族自治州元阳梯田核心区(102°43′~102°47′ E,23°5~23°8′ N),为亚热带山地季风气候。年平均气温为20.5℃,年降雨量1 397.6 mm,全年大约179.5 d 有雾,363 d 无霜,年均日照1 820.8 h。1 200 m 海拔以下为河谷区,常年无霜,降雨充足,气候炎热;在1 200~1 700 m 海拔段之间为中低山区,雨量充沛,气候温和;1 700 m以上为中高山区,多雾多雨。
1.2 样品采集
土壤样品于2021 年6 月在元阳梯田采集,采样地点设置10 m×10 m 的样方,按照“S”型5 点采样法采集0~20 cm 和20~40 cm 土层土壤,共获得土壤样品36 份。测定土壤酶活性的土壤样品置于4 ℃冰箱保存,测定土壤养分含量的样品自然风干后研磨,分别过孔径0.25 mm 和1 mm 筛,去除土壤中的根茬、动植物残体和石块等杂物。
1.3 土壤理化性质的测定
土壤pH 值采用电极电位法(2.5 ∶1 水土比浸提液)测定,有机质(OM)采用重铬酸钾氧化-外加热法测定,土壤全氮(TP)采用自动定氮仪法(NY/T 1121.24-2012)测定,土壤全磷(TP)采用碱熔-钼锑抗分光光度法(HJ632-2011)测定,土壤全钾(TK)采用NaOH 熔融-火焰光度法测定,碱解氮(AN)采用碱解扩散法测定,有效磷(AP)采用0.5 mol/L NaHCO3溶液浸提-钼锑抗比色法测定,速效钾(AK)采用醋酸铵-火焰光度计法测定。
1.4 土壤酶活性的测定
过氧化氢酶活性采用高锰酸钾(0.1 mol/L)滴定法测定,蔗糖酶活性采用3,5-二硝基水杨酸比色法测定,酸性磷酸酶活性采用对磷酸苯二钠比色法(缓冲液pH=5)测定,脲酶活性采用靛酚蓝比色法测定[15]。
试验数据采用Excel 2016 软件进行整理和绘图,采用SPSS 23 软件进行单因素方差分析和差异显著性检验,检验方法采用ANOVA 法,相关分析采用Pearson 相关。
2 结果与分析
2.1 不同植被类型土壤理化性质的变化
不同植被类型在不同采样深度的土壤养分和化学性质如表1 所示。
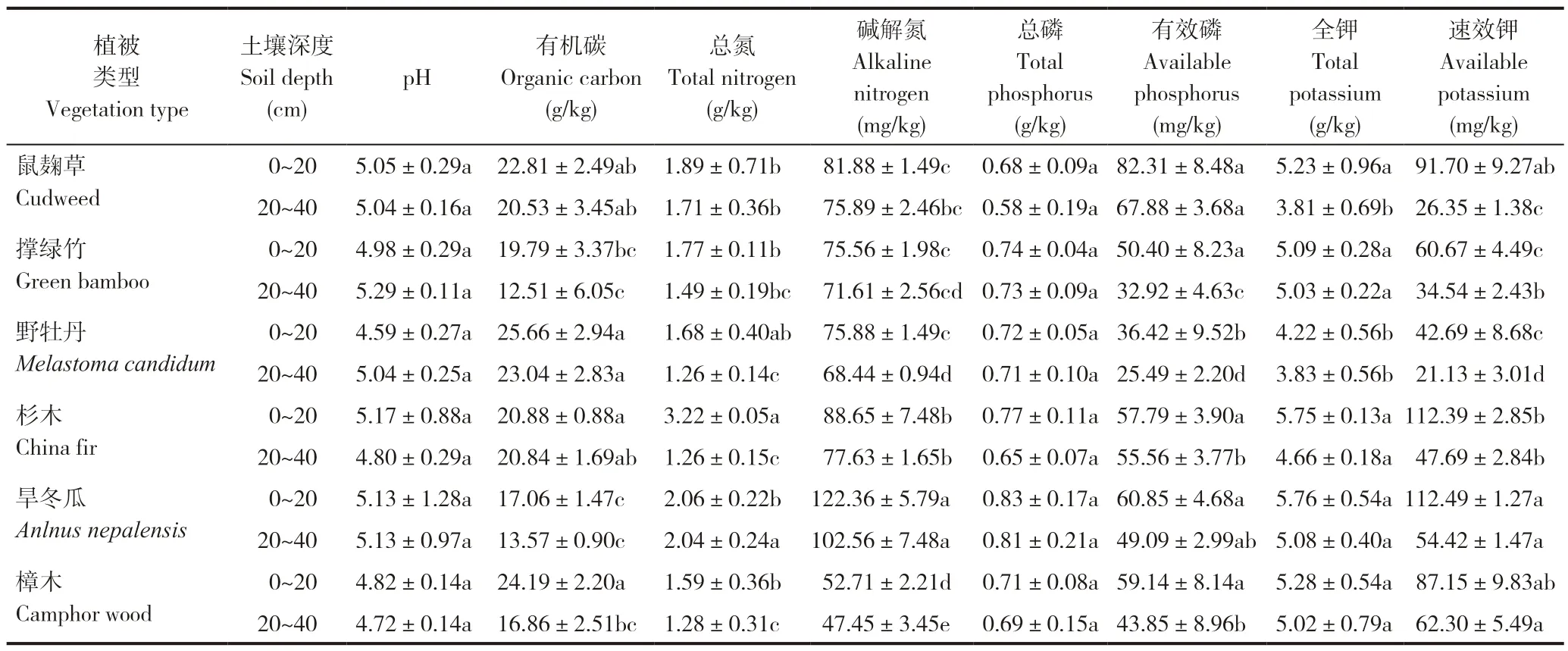
表1 不同植被类型土壤基本理化性质Table 1 Basic physicochemical properties of soils in different vegetation types
2.1.1 pH 值 不同植被类型土壤pH 在4.59~5.29之间,属于酸性土壤,不同植被类型土壤pH 差异不大。
2.1.2 有机碳 土壤有机碳是反映土壤质量的重要指标,不同植被类型间土壤有机碳含量差异显著。土壤机碳含量在0~20 cm 土层高低依次为:野牡丹>樟木>鼠麹草>杉木>撑绿竹>旱地,在20~40 cm 土层高低依次为:野牡丹>杉木>鼠麹草>樟木>旱冬瓜>撑绿竹,各植被类型土壤有机碳含量均表现为0~20 cm >20~40 cm 土层,野牡丹土壤有机碳含量在0~20 cm 土层和20~40 cm 土层均显著高于撑绿竹和旱冬瓜。6 种植被类型0~40 cm 土层土壤有机碳含量平均值为:鼠麹草21.67 g/kg,撑绿竹16.15 g/kg,野牡丹24.35 g/kg,杉木20.86 g/kg,旱冬瓜15.32 g/kg,樟木20.53 g/kg,对照全国土壤养分分级标准,野牡丹处于一级水平,鼠麹草、杉木和樟木处于二级水平,旱冬瓜和撑绿竹处于三级水平。
2.1.3 全氮和碱解氮 土壤全氮含量在0~20 cm土层变化范围在1.59~3.22 g/kg 之间,杉木土壤全氮含量显著高于鼠麹草、撑绿竹、樟木和旱冬瓜,20~40 cm 土层土壤全氮变化范围在1.26~2.54 g/kg之间,旱冬瓜土壤全氮含量显著高于鼠麹草、撑绿竹、野牡丹、杉木和樟木,各植被类型土壤全氮含量均表现为0~20 cm >20~40 cm 土层。土壤碱解氮含量在0~20 cm 土层和20~40 cm 土层均表现为旱冬瓜显著高于鼠麹草、撑绿竹、野牡丹、杉木和樟木,各植被类型土壤碱解氮含量均表现为0~20 cm >20~40 cm 土层。6 种植被类型0~40 cm 土壤全氮含量平均值为:鼠麹草1.8 g/kg,撑绿竹1.63 g/kg,野牡丹1.47 g/kg,杉木2.24 g/kg,旱冬瓜2.05 g/kg,樟木1.44 g/kg,对照全国土壤养分分级标准,旱冬瓜和杉木处于一级水平,鼠麹草和撑绿竹处于二级水平,野牡丹和樟木处于三级水平。6 种植被类型0~40 cm 土层土壤碱解含量平均值为:鼠麹草78.89 mg/kg,撑绿竹73.59 mg/kg,野牡丹72.16 mg/kg,杉木83.14 mg/kg,旱冬瓜112.46 mg/kg,樟木50.08 mg/kg,对照全国土壤养分分级标准,旱冬瓜处于三级水平,鼠麹草、撑绿竹、野牡丹、杉木处于四级水平,樟木处于五级水平。
2.1.4 全磷和有效磷 土壤全磷含量在0~20 cm土层表现为旱冬瓜>撑绿竹>杉木>野牡丹>樟木>鼠麹草,在20~40 cm 土层表现为旱冬瓜>撑绿竹>野牡丹>樟木>杉木>鼠麹草。6 种植被类型在0~20 cm 土层和20~40 cm 土层土壤全磷含量差异不显著。这可能是因为土壤全磷的分布趋势主要与根系分布和不同植被对土壤剖面磷的吸收利用有关。有效磷含量在0~20cm 土层表现为鼠麹草>旱冬瓜>樟木>杉木>撑绿竹>野牡丹,野牡丹土壤有效磷含量显著低于鼠麹草、旱冬瓜、樟木、杉木和撑绿竹。在20~40 cm 土层土壤有效磷含量表现为鼠麹草>杉木>旱冬瓜>樟木>撑绿竹野牡丹,且鼠麹草中的有效磷含量显著高于杉木、野牡丹和撑绿竹中的有效磷含量。6 种植被类型0~40 cm 土层土壤全磷含量平均值为:鼠麹草0.63 g/kg,撑绿竹0.74 g/kg,野牡丹0.71 g/kg,杉木0.71 g/kg,旱冬瓜0.81 g/kg,樟木0.7 g/kg,对照全国土壤养分分级标准,旱冬瓜处于二级水平,鼠麹草、撑绿竹、野牡丹、杉木和樟木处于三级水平。6 种植被类型0~40 cm 土层土壤有效磷含量平均值为:鼠麹草72.10 mg/kg,撑绿竹41.66 mg/kg,野牡丹30.96 mg/kg,杉木56.68 mg/kg,旱冬瓜54.97 mg/kg,樟木51.50 mg/kg,对照全国土壤养分分级标准,野牡丹处于一级水平,鼠麹草、撑绿竹、杉木、旱冬瓜樟木处于二级水平。
2.1.5 全钾和速效钾 土壤全钾含量在0~20 cm土层表现为旱冬瓜>杉木>樟木>鼠麹草>撑绿竹>野牡丹,在20~40 cm 土层表现为旱冬瓜>撑绿竹>樟木>杉木>野牡丹>鼠麹草。各植被类型土壤全钾含量均表现为0~20 cm >20~40 cm土层。土壤速效钾含量在0~20 cm 土层表现为旱冬瓜>杉木>鼠麹草>樟木>撑绿竹>野牡丹,在20~40 cm 土层表现为樟木>旱冬瓜>杉木>撑绿竹>鼠麹草(26.35 mg/kg)>野牡丹。且随土层深度的增加,土壤速效钾含量降低。各植被类型土壤速效钾含量均表现为0~20 cm >20~40 cm土层。6 种植被类型0~40 cm 土层土壤全钾含量平均值为:鼠麹草4.52 g/kg,撑绿竹5.06 g/kg,野牡丹4.03 g/kg,杉木5.21 g/kg,旱冬瓜5.42 g/kg,樟木5.15 g/kg,对照全国土壤养分分级标准,鼠麹草、撑绿竹、杉木、旱冬瓜和樟木处于五级水平,野牡丹处于六级水平。6 种植被类型0~40 cm 土层土壤速效钾含量平均值为:鼠麹草59.01 g/kg,撑绿竹47.61 g/kg,野牡丹31.91 g/kg,杉木80.04 g/kg,旱冬瓜83.46 g/kg,樟木74.73 g/kg,对照全国土壤养分分级标准,鼠麹草、杉木、旱冬瓜和樟木处于四级水平,撑绿竹和野牡丹处于五级水平。
2.2 不同植被类型土壤酶活性的变化
不同植被类型0~40 cm 土层土壤酶活性变化如表2 所示。

表2 不同植被类型0~40 cm 土层土壤酶活性测定结果Table 2 Determination results of soil enzyme activities at 0-40 cm of different vegetation types
2.2.1 土壤蔗糖酶 土壤蔗糖酶可以增加土壤中易溶性物质,其活性与土壤有机质的转化和呼吸强度密切相关。如图1A 所示,不同植被类型土壤蔗糖酶活性变化范围在0.70~81.74 mg/g·24h之间,随着土层深度的增加,土壤蔗糖酶活性均呈现出降低的趋势。0~20 cm 土层土壤蔗糖酶活性表现为:杉木>撑绿竹>野牡丹>旱冬瓜>鼠麹草>樟木,撑绿竹和杉木显著高于鼠麹草和樟木。20~40 cm 土层土壤蔗糖酶活性表现为:撑绿竹>杉木>野牡丹>旱冬瓜>鼠麹草>樟木,撑绿竹和山木显著高于其他植被类型。
2.2.2 土壤脲酶 土壤脲酶是催化土壤尿素的唯一酶类,其酶促水解产物为氨气和二氧化碳,土壤脲酶活性的高低常被用来表征土壤氮素的供应程度大小。如图1B 所示,不同植被类型土壤脲酶活性变化范围在10.56~106.73 mg/g·24 之间,随着土层深度的增加,土壤脲酶活性均呈现出降低的趋势。0~20 cm 土层土壤脲酶活性表现为:杉木>旱冬瓜>撑绿竹>野牡丹>鼠麹草>樟木,杉木显著高于其他植被类型。20~40 cm 土层土壤脲酶活性表现为:撑绿竹>旱冬瓜>杉木>野牡丹>鼠麹草>樟木,撑绿竹和旱冬瓜显著高于其他植被类型。

图1 不同植被类型对土壤蔗糖酶、脲酶、蛋白酶、过氧化氢酶活性的影响Fig.1 Effects of different vegetation types on soil sucrase,urease,protease and catalase activities
2.2.3 土壤蛋白酶 土壤蛋白酶能水解各种蛋白质以及肽类等化合物为氨基酸,因此土壤蛋白酶活性与土壤中氮素营养的转化状况有及其重要的关系。如图1C 所示,不同植被类型土壤蛋白酶活性变化范围在4.59~356.41 mg/g·h 之间,随着土层深度的增加,土壤蛋白酶活性均呈现出降低的趋势。0~20 cm 土层蛋白酶活性表现为:旱冬瓜>杉木>撑绿竹>鼠麹草>野牡丹>樟木,旱冬瓜显著高于其他植被类型。20~40 cm 土层土壤蛋白酶活性表现为:撑绿竹>旱冬瓜>杉木>野牡丹>鼠麹草>樟木。
2.2.4 土壤过氧化氢酶 土壤过氧化氢酶广泛存在于土壤和生物体内中,其活性与土壤有机质含量和微生物数量有关。如图1D 所示,不同植被类型土壤过氧化氢酶活性变化范围在0.14~0.57 mg/g·24h 之间,随着土层深度的增加,土壤过氧化氢酶活性均亦呈降低的趋势。0~20 cm土层土壤过氧化氢酶活性表现为:杉木>撑绿竹>鼠麹草>旱冬瓜>野牡丹>樟木,杉木显著高于鼠麹草、野牡丹、旱冬瓜、和樟木。20~40 cm土层土壤过氧化氢酶活性表现为:杉木>撑绿竹>旱冬瓜>鼠麹草>樟木>野牡丹,野牡丹和樟木显著低于其他植被类型。
2.3 土壤理化性质与土壤酶活性的相关分析
为了进一步研究元阳梯田土壤理化性质与土壤酶活性之间的关系,将土壤理化性质与土壤酶活性进行Pearson 相关分析,结果见表3。4 种酶的活性存在较高的相关性,其中蛋白酶与过氧化氢酶、蔗糖酶和脲酶呈极显著正相关(r=0.654,P<0.01;r=0.667,P<0.01;r=0.584,P<0.01)。从酶活性与土壤理化性质的相关性来看,过氧化氢酶与总氮和pH 呈显著正相关(r=0.324,P<0.05;r=0.356,P<0.05),与碱解氮呈极显著正相关(r=0.390,P<0.01)。蛋白酶与总氮、碱解氮呈极显著正相关(r=0.548,P<0.01;r=0.694,P<0.01),与有效磷呈显著正相关(r=0.317,P<0.05)。蔗糖酶与碱解氮呈极显著正相关(r=0.413,P<0.01),脲酶与碱解氮呈显著正相关(r=0.340,P<0.05)。
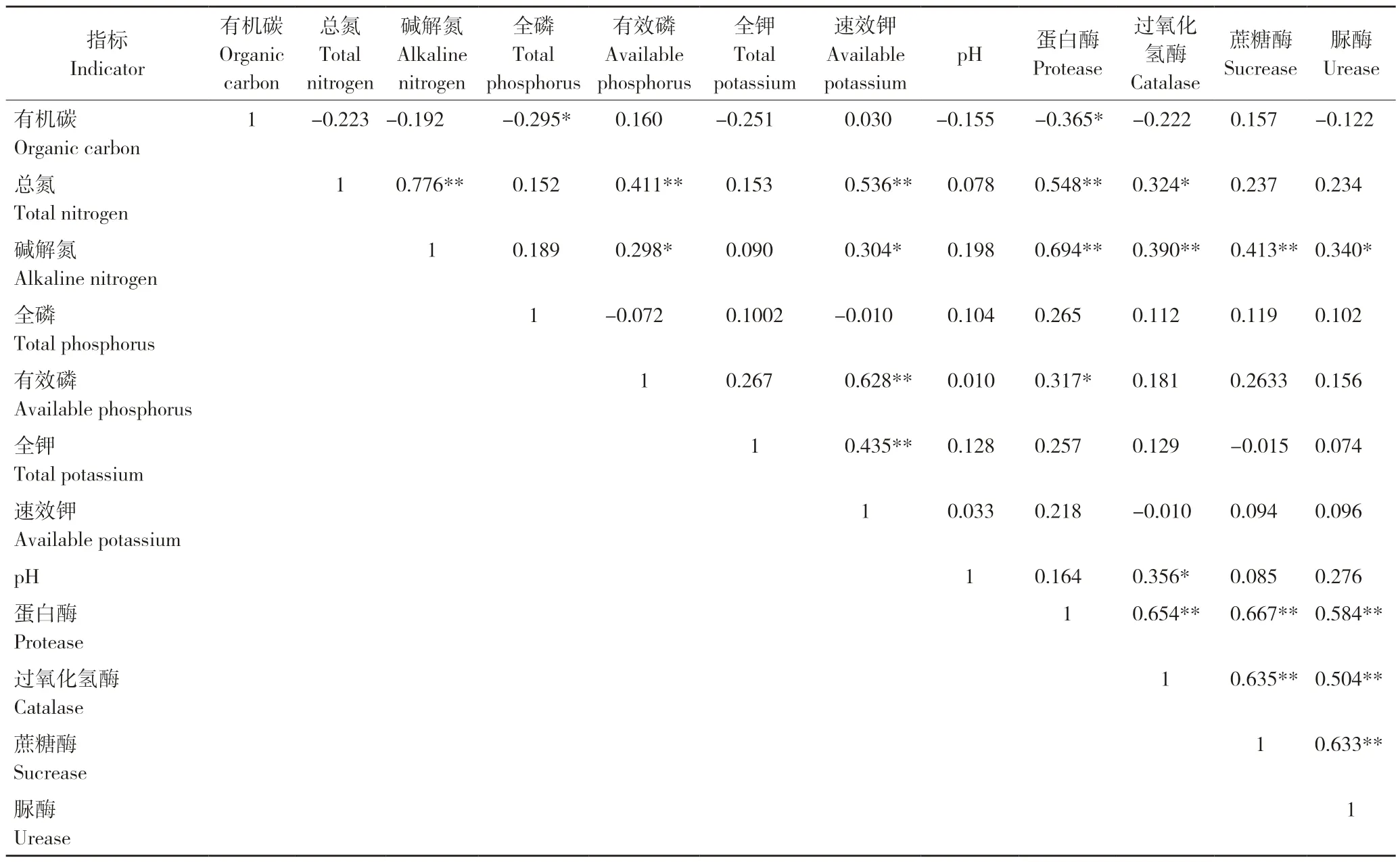
表3 土壤理化性质与土壤酶活性的相关分析Table 3 Correlation analysis between soil physicochemical properties and soil enzyme activities
3 讨论
3.1 不同植被类型对土壤理化性质的影响
有研究表明,由于不同植被类型林下凋落物的组成和分解状况不同,土壤养分含量也会存在较大差异[21-22]。本研究发现不同植被类型土壤养分含量在垂直空间分布上具有显著差异,具体表现为不同植被类型0~20 cm 土壤养分含量高于20~40 cm 土壤,这与前人的研究结果一致[23-26]。这可能是因为植物根系影响了土壤养分在垂直空间的分布[27-28]。郝宝宝等[25]认为这是因为凋落物在土壤表层分解形成的有机质在降雨、地表径流等的作用下向深层土壤缓慢运移所导致的。本试验发现,所测养分随土层深度的增加,养分含量均呈现降低的趋势。除有机碳和有效磷分别在野牡丹和鼠麹草的含量最高外,总氮、碱解氮、总磷、全钾、速效钾均表现为在旱冬瓜的含量最高,一方面说明旱冬瓜提高了土壤养分质量分数,并且对改良土壤和防止地力衰退具有重要作用,另一方面也表明植被类型与土壤养分有着密切的联系。
有机碳含量的高低主要取决于有机物质的输入和输出[29-30]。王雅等[12]研究表明,中国黄土高原4 种植被类型土壤有机碳含量范围在2.44~11.73 g/kg,而本研究中不同植被类型土壤有机碳含量普遍较高,0~20 cm 土壤有机碳平均含量为22.40 g/kg,20~40 cm 土壤有机碳平均含量为18.31 g/kg。与撑绿竹和旱冬瓜相比,野牡丹土壤有机碳含量更高,这可能是因为不同植被类型的生物量和凋落物数量不同,使得输入土壤的有机碳含量差异较大,从而使不同植被类型的有机碳含量差异显著[19,31]。
3.2 不同植被类型对土壤酶活性的影响
土壤酶是土壤中有机体代谢的主要动力,是反映生态系统土壤肥力和养分转化过程的重要指标[32-33]。土壤脲酶、土壤蔗糖酶在土壤碳、氮循环中起着重要的作用。土壤脲酶不仅是土壤中氮转化的重要催化剂,而且还对提高土壤氮的利用效率有至关重要的作用[34]。土壤蛋白酶参与着土壤中氨基酸、蛋白质的转化过程[35]。土壤过氧化氢酶直接影响着土壤氧化还原,参与微生物呼吸过程的物质代谢作用,可表示土壤氧化还原过程的强度。元阳梯田不同植被类型下土壤酶活性研究结果显示,不同植被类型土壤酶活性高低均存在一定的差异。从酶活性的空间分布来看,土壤蔗糖酶、土壤脲酶、土壤蛋白酶和土壤过氧化氢酶均呈现明显的垂直分布现象,具体表现为各植被类型土壤酶活性在0~20 cm 土层土壤高于20~40 cm 土层土壤,这与前人的研究结果一致[36]。土壤蔗糖酶、脲酶、蛋白酶和过氧化氢酶活性间呈现出极显著正相关关系,表明这3 种酶在进行反应时,一方面具有特定的专一性,另一方面说明其存在一定的共性[37]。
本研究中,6 种植被类型土壤蛋白酶活性存在较大差异,撑绿竹、杉木和旱冬瓜土壤蛋白酶活性显著高于其他3 种植被类型,表明撑绿竹、杉木和旱冬瓜土壤氮转化速率高于鼠麹草、野牡丹和樟木。撑绿竹和杉木土壤过氧化氢酶活性显著高于其他4 种植被类型,说明杉木和撑绿竹土壤的氧化还原程度高于鼠麹草、野牡丹旱冬瓜和樟木。本研究还发现蔗糖酶和脲酶在杉木土壤中的活性最高,且杉木土壤中的总氮含量显著高于其他植被类型土壤,表明杉木土壤中的氮素转化速率高于其他5 种植被类型土壤,这与Gong 等[38]的研究结果一致。
3.3 土壤肥力和酶活性相关性分析
郭春秀等[39]研究表明,各类土壤酶在土壤养分循环过程中扮演着重要角色。土壤养分含量通过影响植物和微生物的生长,从而使土壤酶活性发生变化,最终导致土壤酶活性与土壤养分之间的关系更加密切。Xu 等[41]研究表明土壤养分含量的增加或减少会导致土壤酶活性的变化,这可能是由于养分需求和养分供应共同调节了土壤酶活性。本研究中土壤总氮含量和土壤蛋白酶活性呈极显著正相关,与土壤过氧化亲酶活性呈显著正相关,这可能是因为土壤中氮素为土壤酶提供了能量来源,从而促使了土壤蛋白酶和土壤过氧化氢酶活性的提高[42-43]。土壤碱解氮含量与土壤蛋白酶活性、蔗糖酶活性呈极显著相关正相关,与土壤脲酶活性呈显著正相关,这表明土壤蛋白酶、蔗糖酶、过氧化氢酶与碱解氮存在明显的协同工作用,促进了土壤中氮的转化[44]。
4 结论
元阳梯田不同植被类型对土壤理化性质和土壤酶活性有不同程度的影响。不同植被类型土壤pH 无显著差异,土壤养分随着土壤深度的增加均呈现出降低的趋势,垂直分布现象明显。总体来看,土壤蔗糖酶、脲酶、蛋白酶和过氧化氢酶活性均表现为0~20 cm 土层高于20~40 cm 土层。旱冬瓜和杉木在元阳梯田土壤养分持续供给、土壤氮素转化过程中具有一定的优势。
猜你喜欢
防护林科技(2022年2期)2022-04-25
大众文艺(2020年22期)2020-12-13
创作(2020年6期)2020-11-17
农民致富之友(2020年8期)2020-05-11
农家科技下旬刊(2019年12期)2019-01-13
文苑·经典美文(2018年6期)2018-06-23
散文百家(2017年10期)2017-10-10
现代养生·上半月(2017年9期)2017-09-05
现代装饰·理论(2016年9期)2016-12-26
传奇故事(破茧成蝶)(2015年7期)2015-02-28
