
陈皮表面微生物及其转化黄酮类物质的研究进展
2022-04-28 09:02:10傅曼琴陈玉婷吴继军余元善温靖徐玉娟
现代食品科技 2022年4期
傅曼琴,陈玉婷,2,吴继军,余元善,温靖,徐玉娟*
(1.广东省农业科学院蚕业与农产品加工研究所,农业部功能食品重点实验室,广东省农产品加工重点实验室,广东广州 510610)(2.广东海洋大学食品科技学院,广东湛江 524088)
陈皮(Citri Reticulatae Pericarpium,CRP),是植物橘及其栽培品种的果皮经干燥制成[1],陈皮中含有黄酮类、柠檬苦素类、挥发油类、生物碱类等物质,主要以黄酮类化合物为主[1]。大量研究结果表明,陈皮在陈化期间黄酮类化合物逐渐积累,大幅度地提高了陈皮的药用功效。黄酮类化合物有抗炎[2]、抗氧化[3]、抗肿瘤[4]、抗菌[5]、抗病毒[6]、降血脂[7]、抗过敏[8]、抗肥胖[9,10]、促消化[11]、祛痰平喘、解痉[12]、改善学习记忆能力[13]等生物活性。但传统的自然陈化,时间漫长,效率低且容易受到污染导致品质不稳定。另外,陈皮中黄酮类化合物难溶于水、生物利用率低,极大程度地限制了黄酮类化合物的临床应用,微生物转化黄酮类化合物的反应周期短、条件温和,可产生一些新颖活性物质用于新药的开发以及临床的应用。
1 陈皮微生物
陈皮是传统特色中药材,现有研究报道多集中在陈皮化学成分尤其是黄酮类化合物及其生物活性,另有报道在陈皮贮藏放置过程中微生物对其中的活性物质含量有显著影响。阳洁等[14]从陈皮中分离鉴定出的细菌均属于芽胞杆菌属Bacillus或者类芽孢杆菌属Paenibacillus,未分离出放线菌和真菌。王福等[15]从不同批次的陈皮样品中分离得到青霉、桔青霉、黑曲霉和黄曲霉等25株真菌。刘素娟等[16]也从陈皮样品中共分离鉴定出子囊菌类曲霉属和青霉属真菌等10株真菌,黑曲霉和黄曲霉为优势菌群。陈聪聪[17]对广陈皮进行了研究,结果表明,广陈皮样品中菌属有假单胞菌属Pseudomonas、乳球菌属Lactococcus、肠球菌属Enterococcus和节细菌属Arthrobacter。刘丽娜[18]从不同年份陈皮中分离微生物,其中细菌属有假单胞菌属Pseudomonas、鞘脂单胞菌属Sphingomonas、甲基杆菌属Methylobacterium、乳杆菌属Lactobacillus等,检测到的真菌属有耐干霉属Xeromyces、酵母菌属Symmetrospora、蜡菊属Xerochrysium、枝孢霉属Cladosporium、被孢霉属Mortierella、菇属Pleurotus、毛壳菌属Chaetomium、赤霉属Gibberella。张鑫等[19]分析了不同产地陈皮表面真菌群落多样性,主要发现青霉属Penicillium、曲霉属Aspergillus、毛霉属Mucor、枝孢霉属Cladosporium、镰刀菌属Fusarium等。

表1 陈皮微生物种属来源Table 1 Origin of microbial species of citri reticulatae pericarpium
可见,陈皮表面含有大量微生物,且可能参与陈皮活性物质的转化,对陈皮陈化品质的形成产生影响。陈化时间不同,其微生物种类存在差异,年份越久的陈皮中的微生物多样性也越丰富。陈皮中的细菌主要是芽孢杆菌属,真菌主要是曲霉属和青霉属,其中黑曲霉因其安全性和强大的酶系成为微生物转化的主要菌种之一,黑曲霉在食药材陈化过程中参与了部分次生代谢产物的形成。陈皮微生物转化其活性成分的机制亟待进一步研究。
2 陈皮黄酮类活性物质
黄酮(flavonoid,又称类黄酮)是以2-苯基色原酮-4-酮骨架的黄酮类化合物[8],根据化学结构的不同可将黄酮类化合物分为黄酮类(flavones),黄酮醇类(flavonol),二氢黄酮类(flavonones),二氢黄酮醇类(flavanonol),花色素类(anthocyanidins),双苯吡酮类(xanthones),查尔酮(chalcones)和双黄酮类(biflavonoids)等十几种[8]。陈皮中的黄酮类化合物主要有两种,一种是黄酮苷类,如橙皮苷(hesperidin)、柚皮苷(naringin)等,另一种是多甲氧基黄酮类,主要有川陈皮素(nobiletin)、橘皮素(tangeretin)[20]。
2.1 黄酮苷类
陈皮中含有的黄酮苷类化合物主要是橙皮苷、新橙皮苷、柚皮苷和芸香柚皮苷(图1)[21,22],橙皮苷由一个黄酮苷元和两个单糖组成,是陈皮中含量最高的黄酮苷[23],2015版药典中定义其为陈皮的指标性成分,其含量不得低于3.50%[24]。
随着贮藏时间的延长,陈皮中橙皮苷的含量呈升高趋势[2,23,24],刘丽娜等[25]研究发现不同贮藏年份陈皮橙皮苷含量在15年间增加了2.61倍。杨放晴等研究发现广陈皮中黄酮类化合物的含量随陈化时间增加总体呈增加趋势,其中橙皮苷含量显著增加[26]。品种和产地的不一,也可导致橙皮苷的含量变化不一。张鑫等[27]研究发现陈皮中橙皮苷的含量先增加后减少,Fu等[28]发现陈皮在0~3陈化期橙皮苷的含量有所下降。魏莹等[29]发现随着陈化期的延长陈皮中橙皮苷含量呈下降趋势。陈皮中橙皮苷含量变化可能主要与陈皮中的微生物有关,刘丽娜[18]从不同年份陈皮微生物中筛选出8株具有将橙皮苷转化为橙皮素的真菌,分别为巴西曲霉Aspergillus brasiliensis、黑曲霉Aspergillus niger、塔宾曲霉Aspergillus tubingensis、棘孢曲霉Aspergillus aculeatus、 简 青霉Penicillium simplicissimum、互生交链孢霉Alternariaalternata、出芽短梗霉Aureobasidium pullulans和葡萄牙棒孢酵母Clavispora lusitania。由此说明陈皮陈化过程中橙皮苷含量的变化可能与其微生物有关。此外,在不同的陈化阶段,陈皮微生物多样性及其代谢物成分变化亦有显著差异[17,18],表明微生物转化对于陈皮黄酮类活性物质的积累起到重要作用,可能是陈皮“越陈越佳”的关键因素。
2.2 多甲氧基黄酮类
多甲氧基黄酮是一类高度甲基化的黄酮类化合物,在柑桔属植物中广泛存在,尤其是柑桔果皮中,是柑橘属植物独有的一类天然黄酮化合物[30]。迄今为止,陈皮中鉴定出的多甲氧基黄酮类化合物有五十多种[30],其中陈皮中含量最高的是川陈皮素[31],其次是橘皮素、5-羟基-6,7,8,3',4'-五甲氧基黄酮和3,5,6,7,8,3',4'-七甲氧基黄酮[32]。张珂等从陈皮中初步鉴定了61个化学成分,主要以黄酮类化合物为主,而从陈皮中首次发现10个含有3-羟基-3-甲基戊二酸取代基的多甲氧基黄酮氧苷成分[33]。童超英等首次在陈皮中分离出新圣草次苷、柠檬黄素-3-O-(3-羟基-3-甲基戊二酸)葡萄糖苷及其异构体[34]。对于不同陈化年限陈皮中多甲氧基黄酮类化合物也有研究,杨放晴[26]等用超高效液相色谱-四极杆-静电场轨道阱高分辨质谱法技术分析不同陈化时间广陈皮中的黄酮类化合物,分析结果表明6-去甲氧基橘皮素、川陈皮素、橘皮素的相对含量增加明显。盛钊君等[35]通过高效液相色谱与分光光度法两种方法,测定新会柑胎仔、1年青皮、5年陈皮和10年陈皮中多甲氧基黄酮类化合物的含量,两种方法检测结果均表明柑胎仔中多甲氧基黄酮含量远远高于青皮或陈皮。Fu等[28]研究陈化时间与黄酮含量是否具有相关性时发现川陈皮素、橘皮素、5-羟基-6,7,8,3',4'-五甲氧基黄酮和3,5,6,7,8,3',4'-七甲氧基黄酮呈增加趋势,与刘丽娜等[25]研究1、5和15年新会陈皮时的结论相一致。相反,杨宜婷等[36]研究发现随着贮藏时间的延长,陈皮中多甲氧基黄酮的含量降低,魏莹等[29]发现川陈皮素和橘皮素含量随陈化年限的增加(1~19年)而增加,之后出现反复,研究者陈彤[37]也进行了验证并且发现七甲氧基黄酮的含量随贮藏时间的延长呈波动性变化,规律性不明显。张怀等[38]研究发现陈化年份和川陈皮素没有相关性,陈化年份和橘皮素呈负相关,陈化时间对陈皮橘皮素和川陈皮素的影响也与陈皮的具体产地、品种和采收期有关。在陈皮长期陈化过程中,植物细胞逐渐失去活性,植物自身的酶影响多甲氧基黄酮类成分变化的可能性不大,那么是否与其所含微生物有关?在前文推测陈皮中真菌将橙皮苷脱糖基化生成橙皮素的基础上,进一步假设黄酮苷类脱糖基化生成羟基黄酮,然后通过氧甲基化生成多甲氧基黄酮(图2),具体机制有待进一步验证。
3 微生物转化陈皮黄酮类化合物
微生物对陈皮黄酮类化合物的转化主要表现在陈化过程中,陈皮微生物代谢所产生的酶使其黄酮类化合物的组成及含量产生显著变化。王福等[15]通过将陈皮微生物接入无菌陈皮中,结果发现陈皮霉变后,橙皮苷的含量呈增加趋势。刘丽娜等[18]发现8株真菌对橙皮苷有转化作用,但对其转化机制并未研究。Wang等[39]研究发现将黑曲霉接种陈皮后,川陈皮素的含量呈增加趋势,橙皮苷的含量先增加后降低。杨丹等[40]研究黑曲霉发酵对陈皮黄酮类化合物的影响,结果表明与未发酵陈皮相比,发酵陈皮黄酮含量较高,随着发酵时间的延长,橙皮苷含量和川陈皮素含量逐渐升高,总黄酮和橘皮素的含量先增加后降低。张鑫等[41]研究发现,将黑曲霉接种陈皮样品后,黄酮成分含量均高于接种前,表明黑曲霉的转化对陈皮活性化合物的积累发挥了重要作用。微生物转化黄酮类物质发生的主要反应有甲氧基化、糖基化、羟基化等,真菌脱糖基化、羟基化、在羟基上进行甲基化等。
3.1 糖基化反应
糖基化反应是活性化合物结构修饰或合成的重要反应,黄酮类化合物化合物的理化性质可极大程度的被改变。Dymarska等[42]利用昆虫病原真菌Isaria fumosoroseaKCH J2作为生物催化剂,发现糖基化主要发生在6-甲基黄酮的8位和4'位上(图3),同时糖基的引入提高了6-甲基黄酮的水溶性、稳定性及其生物利用率。Dymarska等[43]利用I. fumosorosea(J1.4和J1.6)和KCH J2对槲皮素、黄芩素、3-羟基黄酮和3-甲氧基黄酮(图3)四种黄酮类化合物进行糖基化反应并获得了相应的吡喃葡萄糖苷,糖基化后的产物水溶性都显著提高。Chiang等[44]通过Bacillus subtilisATCC 6633的糖基转移酶BsGT110将8-羟基黄豆苷元转化成了2种新的异黄酮苷类化合物:8-羟基黄豆苷元-7-O-β-葡萄糖苷和8-羟基黄豆苷元-8-O-β-葡萄糖苷,新的异黄酮糖苷的水溶性分别是8-羟基黄豆苷元的9倍和4.9倍,且其稳定性较8-羟基黄豆苷元显著提高,这两种新的异黄酮苷类化合物在医药和化妆品行业具有潜在的应用前景。
3.2 脱糖基化反应
微生物脱糖基化黄酮糖苷以提高苷元的含量是非常普遍的,Da silva等[44]用橙皮苷酶和柚皮苷酶分别处理橙皮苷和柚皮苷发生脱糖基化反应,脱掉鼠李糖转变成为橙皮素-7-O-葡萄糖苷和柚皮素-7-O-葡萄糖苷,发现脱糖基化后的产物比橙皮苷和柚皮苷的抗氧化活性强。郭瑞等[45]研究发现有8种益生菌可使2种主要异黄酮苷类成分转化为各自苷元,其中最佳菌株为植物乳杆菌,随着植物乳杆菌发酵黄芪时间的延长,2种异黄酮苷的转化率也随之增加,最终可达80.00%以上。Hsiao等[46]研究了浸泡和加热黑豆奶加工过程中花青素和异黄酮的转化,以β-葡萄糖苷酶为生物催化剂,飞燕草苷-3-O-葡萄糖苷和花青素-3-O-葡萄糖苷通过脱糖基化反应生成飞燕草苷和花青素。黄玉军等[47]从乳酸菌中筛选出产β-葡萄糖苷酶的菌株,豆乳经产酶目标菌株发酵后大豆异黄酮苷元总量均有显著提高。Gaya等[48]发现乳酸杆菌和双歧杆菌能催化大豆异黄酮、葛根素和染料木素进行脱糖基化反应,转化为相应的黄酮苷元。You等[49]利用黑曲霉的α-L-鼠李糖酶为生物催化剂,催化芦丁(图4)进行脱糖基化反应转化为槲皮素-3-葡萄糖苷。
3.3 羟基化反应
羟基化反应是微生物转化中常见的结构修饰反应,是在化合物的特定位置引入羟基,激活不活泼的C-H键(表2)。利用A. nigerMB和切美辛青霉113(Penicillium chermesinum113)发酵,黄烷酮羟基化为6-羟基黄烷酮和4'-羟基黄烷酮,6-羟基黄烷酮羟基化为6,4'二羟基黄烷酮[50]。在菌株A. nigerNBRC 4414作用下7,4'-二甲氧基异黄酮羟基化为6-羟基-7,4'-二甲氧基异黄酮,转化率为36.00%[51]。柚皮素在菌株A.nigerCGMCC 3. 4628作用下,C-6,C-8位羟基化分别生成6-羟基柚皮素,8-羟基柚皮素,转化率分别为38.00%和43.00%[52]。白曲霉A. candidus生物转化异雄酮可生成10种羟基化衍生物,羟基化主要发生在C-1α,C-11α和C-15β位[53]。利用链霉菌MA-4680发酵,大豆苷元和染料木素发生羟基化反应生成3',4',7-三羟基异黄酮,3',4',5,7-四羟基异黄酮,羟基化后的产物抗氧化活性显著增强[54]。
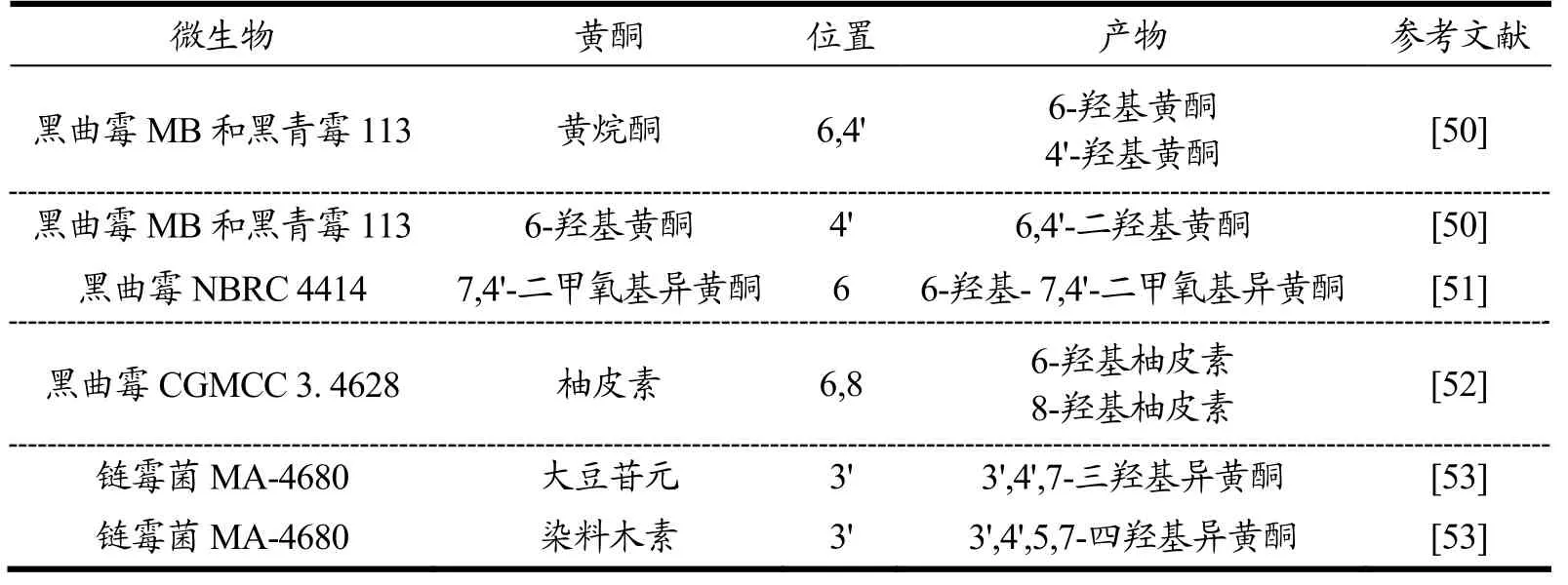
表2 微生物对黄酮类化合物的羟基化作用Table 2 Hydroxylation of flavonoids by microbial

表3 微生物对黄酮类化合物的O-去甲基化作用Table 3 O-demethylation of flavonoids by microbes
3.4 甲基化反应
甲基化反应是黄酮类化合物重要的修饰反应,甲基化修饰后可以增强天然活性物质的活性,从Streptomycessp.KCTC 0041BP菌株中获得的甲基转移酶能够以区域选择性方式向4'-OH添加甲基[55],通常在4'-OH处具有甲基化的类黄酮趋向于生物有效且化学稳定,因此该转移酶可用于生产多种生物活性分子及其衍生物。灰色链霉菌(S. griseus)对黄酮醇类(如槲皮素和漆黄素)的O-甲基化大多发生在C-3'和C-4'位,对黄酮类(如木犀草素和黄芩素)的O-甲基化大多发生在C-6(OH)位[56]。在球孢白僵菌ATCC 7159的发酵作用下槲皮素甲基化为3'-甲氧基槲皮素[57]。Kostrzewa-Suslow等[58]筛选出曲霉和青霉将7-羟基黄烷酮邻甲基化为7-甲氧基黄烷酮和3',4'-二羟基-7-甲氧基黄烷酮。Kostrzewa-Suslow等利用Chaetomiumsp菌株催化黄芩苷生成5,7-二羟基-6-甲氧基黄酮[59]。
3.5 脱甲基化反应
微生物对多甲氧基黄酮类化合物的去甲基化倾向于发生在B环的C-3'和C-4'位置(表2),5,6,7,3',4'-五甲氧基黄酮在菌株A. nigerNBRC4414作用下脱甲基生成4'-甲基-5,6,7,3'-四甲氧基黄酮[51],两种黄酮类化合物对化学诱变剂均具有显著的抗诱变活性,且4'-甲基-5,6,7,3'-四甲氧基黄酮表现出比底物更高的抗诱变活性。在菌株A. niger的作用下塞尼汀、6,7,4'三甲氧基异黄酮和5,7,4'-三甲氧基异黄酮脱甲基化分别生成4'羟基-5,6,7,3'-四甲氧基黄酮、4'羟基-6,7-二甲氧基异黄酮和4'羟基-5,7-二甲氧基异黄酮[51],7-甲氧基黄酮与之相似,在C-7位脱甲基,生成相应转化产物[51]。5,6,7,8,4'-五甲氧基黄酮在菌株A. nigerATCC 984199作用下C-4'位脱甲基生成4'-羟基-5,6,7,8-四甲氧基黄酮,转化率为71%。3-羟基橘红素在菌株A.nigerATCC 9142作用下C-4'位脱甲基生成3,4'-二羟基-5,6,7,8-四甲氧基黄酮,转化率为60.00%,而川陈皮素在这两种菌株的发酵作用下转化慢且代谢物含量少,证实了微生物转化具有特定选择性[60]。
3.6 脱氢反应
Kostrzewa-Suslow等[61]通过曲霉属菌株对黄烷酮脱氢反应进行了一些研究。7-羟基二氢黄酮在菌株A.nigerKB作用下发生羰基还原反应,在菌株A. niger13/5作用下C-2,C-3位脱氢化,其中产量最高的产物分别是7-羟基黄烷-4-醇,7-羟基黄酮。在菌株A.ochraceus 456作用下发生脱氢反应,随后羟基化形成4'-羟基-7-甲氧基黄酮。A. niger6/2也选择性地将6-羟基黄烷酮的C2-C3处脱氢生成6-羟基黄酮作为单一产物[62]。
4 展望
陈皮为传统中药材,对于陈皮的生物活性、活性物质以及陈皮中微生物已经进行了深入系统的研究。陈皮中活性物质主要以黄酮类化合物为主,黄酮类化合物存在溶解度差,生物利用度低、含量低、难纯化等问题,很难被开发利用。微生物转化因其具有反应温和,安全性高,产生新颖化合物等优点,从而可以提高了化合物的生物利用率。陈皮为久贮药材的代表,其陈化过程中伴随着微生物的生长,已报道的有黑曲霉、朱黄青霉和桔青霉等。将真菌反接陈皮培养,发现黑曲霉对黄酮类成分的影响较大,将黑曲霉反接陈皮培养,动态监测黄酮含量的变化,以黄酮含量上升为指标筛选具有转化功能菌株,下一步研究应在于结合代谢组学和转录组学技术明确其促进黄酮形成的反应途径,以及在代谢途径中相关基因的表达和功能酶活,阐明在陈皮陈化过程中,微生物转化陈皮黄酮的代谢机制,进而为科学仓储、调控陈化过程中,提升陈皮品质提供理论依据。
猜你喜欢
Acta Mathematica Scientia(English Series)(2021年1期)2021-04-08 12:52:22
中国茶叶加工(2020年3期)2020-10-21 08:06:48
铜仁学院学报(2018年6期)2018-07-05 09:47:36
中成药(2017年4期)2017-05-17 06:09:50
益寿宝典(2017年2期)2017-02-26 21:27:52
吉林大学学报(医学版)(2015年3期)2015-12-17 07:47:51
中国当代医药(2015年24期)2015-03-01 02:06:02
中国当代医药(2015年9期)2015-03-01 02:02:08
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:32
江西理工大学学报(2013年1期)2013-03-20 14:57:07
