
确定力量和随机性在生境可入侵性中的重要性
2022-04-27 14:03唐龙刘秋月周庆诗高扬
地球环境学报 2022年1期
唐龙,刘秋月,周庆诗,高扬
1.西安交通大学 人居环境与建筑工程学院,西安 710049
2.西安理工大学 水利水电学院,西安 710048
随着全球化的加速,越来越多的区域被外来植物占领(宋兴江等,2021;王友国等,2021;Cavieres,2021)。农田、牧场以及森林常常大量生长着外来种加拿大一枝黄花(Solidago canadensis)、土荆芥(Dysphania ambrosioides)、藜(Chenopodium album)、野燕麦(Avena fatua)、刺苋(Amaranthus spinosus)的个体,这些入侵植物实质性地降低了农作物、牧草和林木的生存空间、营养、水分以及光的水平,严重影响农业、牧业以及林业的发展;一些河道、池塘等水生生境则分布着外来种凤眼蓝(Eichhornia crassipes)、大薸(Pistia stratiotes)的个体,而这些个体能够使水生生物窒息,堵塞航道。根据我国已公布的四批入侵种名单*国家环保总局.2003-01-10.关于发布中国第一批外来入侵物种名单的通知.索引号:000014672/2003-00014.https://www.mee.gov.cn/gkml/zj/wj/200910/t20091022_172155.htm.环境保护部.2010-01-07.关于发布中国第二批外来入侵物种名单的通知.索引号:000014672/2010-00017.https://www.mee.gov.cn/gkml/hbb/bwj/201001/t20100126_184831.htm.环境保护部,中国科学院.2014-08-20.关于发布中国外来入侵物种名单(第三批)的公告.索引号:000014672/2014-00832.https://www.mee.gov.cn/gkml/hbb/bgg/201408/t20140828_288367.htm.环境保护部,中国科学院.2016-12-20.关于发布《中国自然生态系统外来入侵物种名单(第四批)》的公告.索引号:000014672/2016-01463.https://www.mee.gov.cn/gkml/hbb/bgg/201612/t20161226_373636.htm.,入侵植物占多数(56.3%)。入侵植物扩散快速,其个体一旦大量增加,难以根除。因此,入侵生态学家的重要任务是:阐明土著生境被外来植物种群占据的过程与原因,即生境可入侵性(或称为土著群落可入侵性,invasibility),进而为预警、预防、控制外来植物入侵提供理论依据。
一个物种的个体数量,即该物种种群在特定时空范围的尺寸(population size),取决于该物种个体的出生率、死亡率、迁入率和迁出率。当出生率和迁入率之和大于死亡率和迁出率之和,这个物种的个体数量是增加的,种群尺寸是扩大的。由于土著植物个体数量减少、外来植物个体数量增加,土著植物个体被外来植物个体取代,进而导致土著生境中植物物种组成发生显著变化,是土著生境被外来种入侵的生态学实质(Cavieres,2021)。因而,阐明生境可入侵性的研究需要以物种组合理论为依据。
生态位理论和中性理论是物种组合、群落构建的两个主要理论,它们分别强调确定过程和随机过程在物种组合中的效应。这两种理论存在显著的矛盾,但都可以解释、预测一些类型的土著生境的可入侵性。本文将总结、对照两种理论在剖析生境可入侵性中的主要应用,在此基础上,提出了一个观点,即综合确定与随机过程来理解和预测群落可入侵性,并探讨了综合两种过程的方法,以期为未来的研究提供一个方向,为进一步明确可入侵性理论框架提供参考。
1 确定过程与生态位理论
确定力量是指那些可以影响个体的出生率、死亡率、迁入率和迁出率的环境因素,这些因素包括非生物因素和生物因素。非生物因素一般指营养、水、光等资源型因素,以及土壤pH、土壤盐度、大气温度等非资源型因素(周淑荣和张大勇,2006;牛克昌等,2009;Tang et al,2014;D’Andrea and Ostling,2017;Cavieres,2021)。生物因素是与一个个体发生相互作用的其他物种个体。例如:共享资源的竞争物种个体、取食生物量的消费物种个体、吸取体内营养的寄生物种个体等(周淑荣和张大勇,2006;牛克昌等,2009;Tang et al,2014;D’Andrea and Ostling,2017;Cavieres,2021)。这些生物因素大多数都不能被生物个体吸收并转化成生物量或新个体,因此是非资源型环境因子。
生态位是一个抽象的概念。经过长期的进化,所有植物种对每个环境因素的水平都有一定的耐受范围。一个植物种耐受所有环境因素范围的组合称之为该物种的生态位。这是经典的生态位定义,即超维生态位(n-dimensional niche),每个维代表一个环境因素的水平。生态位新定义是物种对环境水平组合的响应以及影响。响应是指存活、生长等表现,而影响是指对资源的消耗、对天敌的支持和对空间的占据等(Cavieres,2021;Zhou et al,2021)。
生态位理论并非一个具体的理论,自从生态位概念被提出后,其思想体现在生态学很多理论中,尤其是植物种分布理论和群落物种组成理论。这些理论指出,由于进化,一个植物种对每一个环境因素都有一个最偏好的水平范围。极端的例子是,竞争物种的个体数量为零。当一个环境因素的水平超出了偏好范围,植物个体的生长与繁殖将受到限制而减少,个体增殖速度从而放缓。基于此,生态位理论强调环境因素对植物表现的影响是确定的,即支持的正效应或限制的负效应。因此,确定力量及其相关理论可被简明地总结为:根据植物种的环境偏好和耐受性,以及环境因素的范围,一个物种个体在一个时空范围中的出生率、死亡率、迁入率和迁出率是确定的,因而是可预测的。
2 随机性与中性理论
中性理论是解释物种分布和群落物种组成的一个新理论(周淑荣和张大勇,2006;牛克昌等,2009;Tang et al,2014;D’Andrea and Ostling,2017)。该理论的核心是:同营养级生命体的出生、死亡、迁入和迁出是随机的,不受环境因素的影响;并且,出生率、死亡率、迁入率和迁出率具有物种等价性,即这些决定种群尺寸的行为在物种间没有差异。某一物种多度的偶然上升是其他物种个体数量减少的结果,因此一定时空植物个体总数基本稳定。
Hubbell(2005,2006)是中性理论的主要提出者,通过观察热带雨林中植物种群的动态发现:一个物种个体的出生、死亡、迁入和迁出在区域间没有显著差异,并且,一个区域中,各物种的出生率、死亡率、迁入率和迁出率也没有显著差异。据此,这些学者认为,环境因素对于植物种群大小和动态的影响是很小的,物种的生态位是不存在的,各物种的个体数量随机波动。
3 主要可入侵性理论
目前,揭示群落可入侵性的理论有七项(表1),这些理论的原理都是生态位这一概念(Beaury et al,2020;Cavieres,2021)。在这些理论中,非资源环境因素,例如:土壤pH、盐度以及气温、风速等被称之为“过滤器”,其对外来种造成的生理胁迫水平,是群落可入侵性高低的基础。另一类非资源因素,即生物环境因素则被认为是各种土著生物,这些生物通过对外来物种的竞争、捕食、寄生、共生等种间作用影响外来种的种群大小。资源环境因素更多强调外来种的资源有效性,即外来种可获得的资源量。非生物环境与生物环境间这些环境因素往往是通过相互作用而影响外来种种群扩大的速度,进而影响生境的可入侵性。
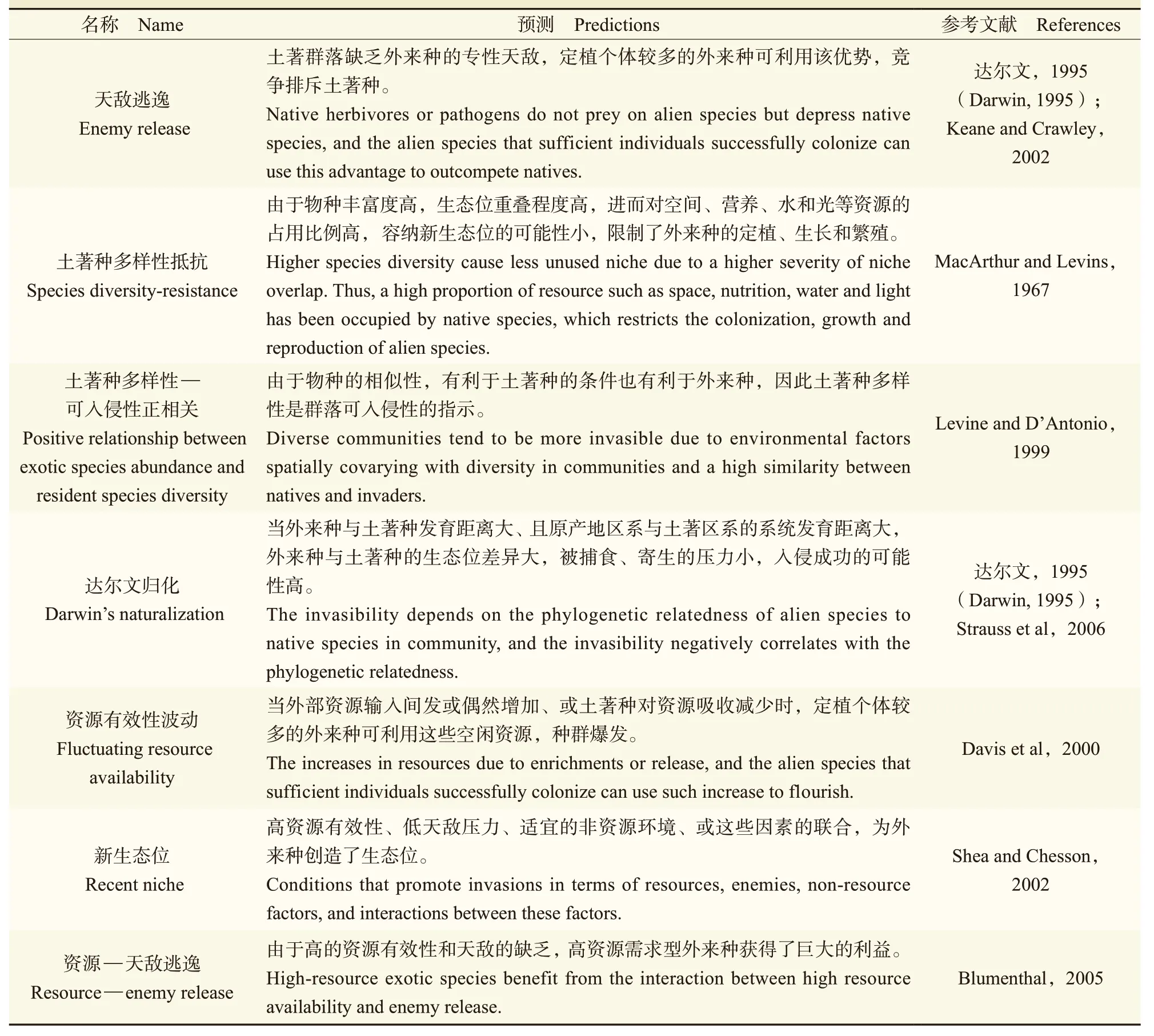
表1 七项群落可入侵性理论Tab.1 Seven theories describing the mechanism of community invasibility
3.1 天敌逃逸(enemy escape)
天敌逃逸理论又称为食草动物逃逸、捕食者逃逸或生态释放(herbivore escape,predator escape或ecological release),由达尔文(1995)提出,Keane and Crawley(2002)系统发展。该理论的核心是:天敌是植物种群大小重要的调节因素,而外来种的专性天敌在土著群落中是缺乏的,当广性天敌对土著种的影响比外来种的影响大时,该土著群落易于被入侵。这个理论成立的条件可以概括为:(1)外来种在入侵地缺乏其专性天敌;(2)土著种的专性天敌没有发生寄主转变;(3)广性天敌对土著种具有更大的影响;(4)外来种繁殖体定殖的多度大,从而可以利用天敌缺乏这一优势。这些条件意味着,假如外来种是低天敌防御型物种,它就需要具备快速的生长能力和高的繁殖能力;假如外来种是高天敌防御型物种,影响土著群落构建的主要天敌类型应该是广性的。如此一来,竞争的不对称性偏向外来物种,使其形成高密度的种群进而入侵成功。
根据Keane and Crawley(2002)的论述,天敌影响了竞争优势是天敌逃逸理论的核心。因此,依赖于天敌逃逸理论建立的生物控制实践需要慎之又慎。引入天敌有成为新入侵种的风险;同时,由于入侵种分布广泛,能否在一个大的生境范围内降低入侵种的竞争能力,则是天敌引入成功与否的另一项标准。
3.2 物种多样性抵抗(species diversity resistance)
物种多样性抵抗理论又称多样性抵抗理论(diversity resistance),植根于生态位极限相似理论中。生态位极限相似性理论指出:可以共存的物种,它们的生态位相似程度,也被称为生态位重叠程度,必须低于一个水平,否则将发生竞争排斥,该水平称之为极限相似性(Abrams,1983)。据此,物种多样性抵抗理论的核心是:物种丰富度高的群落,其物种之间生态位的重叠程度高,相似性接近极限,群落容纳新生态位的可能性小,群落的可入侵性低(MacArthur and Levins,1967)。
两个著名的实验研究支持了这个理论,并将理论中的物种多样性明确为“邻域尺度物种多样性(neighborhood-scale diversity)”(Levine,2000;Kennedy et al,2002)。Levine(2000)发现,假如土著物种的丢失降低了领域尺度的物种多样性,将提高群落的可入侵性。Kennedy et al(2002)的研究结果支持了这个观点,即当相邻物种丰富度、密度和拥挤程度较高时,留给外来种定殖和个体数量增加的资源是少的。由此可见,物种多样性抵抗理论的实质是:土著种对空间、营养、水和光的占用而引起外来种定植、生长的困难。
3.3 土著种多样性—可入侵性正相关(positive relationship between exotic species abundance and resident species diversity)
土著种多样性—可入侵性正相关这个理论预测,由于入侵种和土著种的相似性,能够影响土著种表现的生物和非生物因素,可能同样影响着外来种的表现,因此土著种的物种多样性和群落的可入侵性呈正相关(Levine and D’Antonio,1999);该理论另一表述为:除非土著种和外来种差异显著,否则调控土著种多样性的那些因素对外来种的生长有同样的调控作用。因此,土著群落物种多样性水平简洁地指示着群落可入侵性水平(Levine and D’Antonio,1999)。
物种多样性水平与可入侵性水平的正相关关系,不能被简单地理解为土著种多样性支持入侵。相反,该理论应该被理解为:支持高土著种多样性的生长条件往往是良好的,因而也有利于外来种的生长。物种间的相互作用也被认为支持多样性—入侵性正相关。由于相似的生境需求,在小空间负相互作用的物种,其多度常常在大空间正相关。这样,土著种多样性是一个有利条件的指示,土著多样性越高的群落,环境条件越有利于生长,入侵越容易。因此,定量化评估影响土著种物种多样性的因素,如干扰、营养、消费者等在可入侵性中的重要性是未来需要关注的工作。
3.4 达尔文归化(Darwin’s naturalization)
达尔文归化理论由达尔文(1995)提出。Strauss et al(2006)研究了美国加利福尼亚草本植物的系统发育,并依据结果系统表述了这个理论:外来种与土著种系统发育的相关性,以及外来种群系与土著种群系系统发育的相关性是决定群落可入侵性的重要力量。该理论强调,当一个土著群落面临不同区系的外来物种时,其可入侵性是高的。
达尔文归化假说有三个相关性的度量(Strauss et al,2006;Trotta et al,2021):(1)外来种与最近亲缘土著种的距离。物种间的亲缘距离反映了种间相互作用的强度,与土著种相近的外来种,由于与土著种资源偏好重合程度高而不能定殖成功,这类似于生态位极限相似性的限制。(2)外来种与土著群落多数物种的亲缘相似性。外来种与土著群落多数物种的亲缘相似性潜在地指示着外来种与多个土著种的相互作用。天敌通过进化,更易将其捕食对象或寄主转变到近源种而非远源种。因此,和土著种差异越大的外来种,从天敌压力中释放的程度越高。(3)外来种原产地群系与土著群系的差异。原产地群系与土著群系之间的距离反映了资源偏好和利用方式的差异,以及抵御天敌能力的差异。入侵成功的外来种更有可能来自与土著群系差异大的群系。根据这些分析,土著种与外来种亲缘差异是群落可入侵性重要的影响因素,差异越显著,群落的可入侵性越高。
3.5 资源有效性波动(fluctuating resource availability)
Davis et al(2000)提出了资源有效性波动理论并指出:资源间歇性富集或释放与入侵繁殖体多度的耦合,决定了群落可入侵性。该理论的核心是:各种原因引起的资源间歇性富集或者释放,同时与入侵繁殖体有效性的巧合决定了群落的可入侵性。该理论的简洁表述是:资源有效性的提高将增加群落的可入侵性(Davis et al,2000;Akin-Fajiye et al,2021)。资源有效性提高的原因包括:土著植被对资源的利用程度降低,资源的供应速度大于土著植被吸收的速度。干扰能降低土著植被对资源的利用,因而提高了外来种资源有效性,这样的干扰包括强的啃食、疾病以及火等(Hobbs and Huenneke,1992)。
该理论潜在的预测:如果没有土著种高强度地竞争资源,一个外来种在入侵一个群落过程中将取得更大的成功,而这样的竞争在干扰发生不久的环境中是不重要的,这类环境中,土著植被已被破坏,进而不可能吸收大量的可利用资源,因而新物种的定植是普遍的。因此,该理论需要验证的外延性预测是:(1)相对于资源输入速率以及利用速率稳定的群落,资源输入速率间歇增加或资源利用率间歇降低的群落,其可入侵性更高。(2)当资源供应和土著植被吸收之间的间隔延长,可入侵性增加。(3)群落可入侵性与物种多样性以及生产力没有必然关系。
该理论强调生境的可入侵性是变化的。Davis et al(2000)指出可入侵性不是静态的或永久的属性,而是一个可以随时间波动的属性。群落可入侵性的变化可在年际甚至年内发生,原因是未被利用资源有效性的变化,因此,土著群落被入侵很可能只是偶然事件。
3.6 新生态位(recent niche)
新生态位理论被表述为:生态位机会是土著群落提供给外来种个体数量增长的可能性。因此,理解生态位机会形成的条件是理解该理论的关键(Shea and Chesson,2002)。生态位机会是在资源、天敌、生理环境的时空变化之上形成的,这些机会在群落类型间变化并将促进群落可入侵性。这样,新生态位理论强调时间、物种库、物种数量的效应,特别是天敌随时间的积累、特化以及与资源、非资源因素的相互作用。
形成生态位机会的事件主要有四类:(1)根据 Tilman 的资源—比率理论(resource ratio hypothesis),外来种应具有更高的资源获取速度、更低的资源需求或更高的资源转化能力(Tilman,1985;Shea and Chesson,2002)。(2)资源有效性的波动可以是确定的(季节)或随机的(年际间气候变化的响应),关键是外来种响应这些波动的时间模式必须与土著种不同。这样,资源在高值和低值之间的波动是入侵者的机会,这被称之为“竞争的相对非线性”机制。(3)外来种具有特殊的局部效应,例如利用化感作用降低邻居密度而增加资源有效性,或改变干扰体系而释放资源。(4)资源的空间变化可通过储存效应以及竞争的相对非线性发生。
这些形成资源机会的事件可以是独立的也可以是交互的(Shea and Chesson,2002),交互的具体形式包括:(1)胁迫程度越高,维持生长所需要的资源越多。在胁迫环境中,当外来种维持生长的资源需求比土著种的低,或者外来种对增加的资源响应更强,是外来种入侵成功的机会。(2)高密度的专性天敌可导致土著种的低密度,进而增加了外来种的资源有效性,也是新生态位形成的机会。(3)一个逃脱了天敌的外来种维持需求低,它可能因此成为强的竞争者。由此可见,生物、非生物因素的综合是创造外来种新生态位的重要力量。(4)低成熟度群落中的机会:形成时间短的群落,其构成物种对于局部条件适应时间短,具有宽生态位的物种少。相反,来自形成时间长的群落的物种,其竞争能力可能更强。
3.7 资源—天敌逃逸(resource—enemy release)
Blumenthal(2005)指出,高资源需求的物种,将在高资源区域从天敌缺失下获利。这个综合性的理论可以理解为:天敌逃逸有利于高资源需求物种在高资源水平区域入侵成功,或者,天敌逃逸将促进高资源需求物种对高资源有效性土著群落的可入侵性。
该理论依据的生态学原理是:资源需求量、生长速度和天敌敏感性之间的权衡。资源稀缺,生长缓慢,进而产生新组织的代谢成本高昂;因此,低资源区域的植物种进化形成了防御系统,这类植物种称之为低资源需求型物种。相反,高资源区域的物种生长快速,生产组织的代谢成本低,进而在防御中的投资小,这类植物种被称为高资源需求型物种。高资源需求物种,其防御性结构物质少、组织营养含量高,更易吸引消费者,因而受到天敌影响更强。这样,当天敌缺失、且资源有效性高时,高资源需求物种将获利更大,即天敌逃逸和资源有效性增加的交互效应将提高可入侵性。
值的注意的是,一些研究发现外来种既可入侵资源丰富的区域,也可在贫瘠区域成为群落优势种。例如:外来互花米草对土著种的竞争排斥优势可以发生在贫瘠的沙子中,也可以发生在营养水平比较高的湿地(Tang et al,2014;Tang et al,2018)。根据这些研究结果,资源—天敌逃逸理论应该存在条件性,其预测入侵成功的范围,只是针对高资源需求型外来种。
4 当前可入侵性理论对确定力量的发展及对随机性的忽视
天敌逃逸理论支持达尔文归化理论、新生态位理论和资源—天敌逃逸理论,因为这些理论都关注了天敌的作用(MacDougall et al,2009;Pinto-Ledezma et al,2020;Cavieres,2021)。达尔文在天敌逃逸理论中强调天敌的直接作用,尤其是专性消费物种的作用(达尔文,1995;Keane and Crawley,2002);并且,达尔文(1995)和Strauss et al(2006)在达尔文归化理论中解释了这些作用缺失的原因,即外来种的专性天敌留在了原产地,以及新区域中专性天敌没有发生变异而取食外来种。新生态位理论和资源—天敌逃逸理论进一步指出了天敌逃逸作用的生态学后果,包括形成了可被外来种利用的生态位(新生态位理论),以及帮助外来种排斥土著种的竞争优势(资源—天敌逃逸理论)(Shea and Chesson,2002;Blumenthal,2005)。这些理论不仅强调了外来种缺乏专性天敌,同时强调了土著种具有专性天敌,这种差别是群落可入侵性的重要组成。
物种丰富度理论和资源有效性波动理论围绕群落资源分析群落可入侵性(MacArthur and Levins,1967;Davis et al,2000)。这里,资源包括土壤营养、土壤水分、光以及空间。两者的区别是:物种丰富度理论强调土著种留给外来种剩余资源的量,而资源有效性波动理论强调资源的输入量以及土著种的利用程度对外来种资源有效性的影响。因此,资源有效性理论的预测潜在地包含着物种丰富度理论的预测。资源波动理论和新生态位理论强调了群落可入侵性是随时间变化的,且这种变化并非单向地加强或减弱,而是波动的。
当前有关可入侵的七项理论,都以天敌、资源或两者的联合作用为基础。在没有消费者取食或病原体限制的条件下,外来种器官表面积单位时间内增大速度更高,这样,外来种后续资源吸收的速度高且吸收量大。据此,外来种在与土著种竞争排斥中占据优势,并且这种优势将被高的资源有效性放大,无论这个高的资源水平是来自外部输入的增加还是土著种吸收量的减小,都会导致高资源群落可入侵性高。资源—天敌逃逸理论将这种优势限定于高资源物种才能具有(Blumenthal,2005;Beaury et al,2020;Beaury et al,2021),达尔文归化理论则将这种优势限定于与土著种差异大的外来种。因此,天敌(尤其专性)缺乏和高资源的联合可以被总结为促进群落可入侵性的确定力量,而外来种的个体数量或者性状特征有可能放大了这些力量的效应。
这七项理论没有涉及中性理论强调的随机性。群落的可入侵性反映在外来种个体数量的增加和土著种个体数量的减少。个体数量的变化不能够排除偶然性和随机性。因此,尽管一些现有可入侵性理论内涵宽泛,可以解释比较多的现象,但依然是不全面的。生态位理论和中性理论在揭示、预测群落物种组成和结构中有各自的优势,也都有缺陷。将这两种理论结合起来构架可入侵性理论的方法,被越来越多的生态学家所接受。
5 生态位理论和中性理论的核心矛盾
生态位理论预测:在既定时空,每个物种的个体数量受到其偏好资源出现频率以及有效性的严格限制,新到达物种也不例外。中性理论预测:群落中各个物种的相对多度是随机的,而总多度基本稳定,新到达物种的定植成功率和多度增加速度是随机的(Hubbell,2005,2006;Holt,2006;Saavedra et al,2017)。
中性理论和生态位理论各有缺点。在预测群落结构动态时,强调确定性因素的生态位理论常常是低效率的,因为这些理论涉及过多的环境因子,不得不引入超维生态位来预测植物可能的分布范围,而如此复杂的计算很难实现。对照地,中性理论没有考虑共存植物种间的相互作用。大量的实验研究发现,邻居存在时一个植物种生物量的下降或增加,证明了种间相互作用的存在(Ricciardi et al,2013;Tang et al,2014;Tang et al,2018)。其次,中性理论没有包含环境条件及其波动。显而易见地,自然生物地理屏障、非资源胁迫因素、资源因素及干扰等可强烈限制植物的分布(Tang et al,2014)。
通过整合中性理论与生态位理论来完善群落构建机理的理论,是群落生态学发展的趋势。生态位理论从实验中获得了强有力的支持,因此中性理论奠基人Hubbell 和许多生态学家认为,不能由于一些缺陷就抛弃生态位理论(Hubbell,2005)。另一方面,中性理论包含了生态位理论忽视的很多方面,例如个体迁移、物种分化、群落大小等,从而强调了随机过程在群落构建中的重要性,并以新物种的迁入为基础,构建了在个体水平上探讨群落结构形成的理论框架;中性理论也获得一些实验证据的支持(Tilman,2004;Adler et al,2007;Vergnon et al,2012)。概括起来,两类理论的差异涉及物种性状、生境、随机移动、以及种间相互作用,这些是群落物种组成的基础,所以有必要统一两者的差异,以促进对植物分布一般规律的认知(Whitfield,2002;Adler et al,2007)。
6 确定力量和随机力量的整合
生态位理论和中性理论引发了入侵生物学相矛盾的预测(牛克昌等,2009;Letten et al,2017)。前者预测群落是稳定的,因而难以被入侵,并且群落的可入侵性与环境及物种的特征紧密联系(D’Andrea and Ostling,2017);后者预测物种趋向于功能等价,个体随机出现和消失,群落易被入侵,并且群落的可入侵性与环境及物种的特征没有实质性的关系(Tilman,2004)。由于群落可入侵性是入侵预警、预防的基础,因此需通过详实的研究仔细验证这些相矛盾的预测(Barabás et al,2013)。
Tilman(2004)和Gravel et al(2006)的工作是整合确定力量和随机力量研究的重要开端。Tilman(2004)发现:随机生态位组合创造了群落,在这些群落中,物种占据等宽的栖息地空间异质性范围;资源水平的轻微下降会导致繁殖体生长至成年的概率大幅下降;高物种多样性群落的低侵入性不是由多样性本身造成的,而是由随机竞争组合造成的资源低水平导致的。这一结果为入侵悖论提供了一个潜在的解决方案。Gravel et al(2006)提出了一个增长概率模型来融合两个力量。模拟结果表明:确定力量和随机力量形成了个体数量从竞争排斥到随机变化的连续体。在没有外来种的情况下,竞争排斥倾向于创造生态位的一个有规律的间隔。然而,外来种将阻碍这样的有限相似性的建立,引起入侵。
近十年来,整合确定力量和随机力量的研究不断发展。Crowder et al(2011)发现生境利用变化导致的生态位差异是影响生态位分布的主要因素,但随机过程影响了生境内稀有物种的建立,因此影响了外来种到达后群落构建模式。Du et al(2011)发现适当的负密度依赖强度可以抵消竞争不对称性对物种共存的负面影响,产生中性的物种分布。Tang and Zhou(2013)发现只有将生态位结构纳入多重离散群落模型,构建生态位—中性混合模型,才可反映物种和个体的分布模式。Pigolotti and Cencini(2013)发现生态位稳定群落支持长寿命物种的高多样性,暗示着中性力量决定短寿命物种的共存。Latombe et al(2015)提出了群落装配阶段空间概念,这个空间是由随机扩散和生态位选择过程确定的一个多维空间。该空间可以指示两类力量在群落构建中的互补程度。Souza et al(2016)指出,由于生态位重叠和聚类沿干扰梯度减少,未来的研究应包括不同的干扰条件和环境变化。这样的方法有利于评估确定力量和随机力量对群落构建过程的影响。Ruziska et al(2018)认为,随机空间构造模型可以准确模拟物种间相对优势的变化。Cao et al(2019)提出,在评价确定力量和随机力量在群落构建中的相对作用时,应考虑物种的个体尺寸,因为随着个体尺寸的增加,确定力量的作用将减弱。
这些整合确定力量和随机力量研究的结果,指示着理解群落可入侵性的重要方向应该是以中性理论为基础构建模型,之后使用确定力量逐步取代随机力量。中性理论强调繁殖体移动的随机性,这为外来种个体到达土著群落提供了必要条件,是土著群落被入侵的开端。到达的外来种个体能否存活,则使用非资源胁迫这一确定力量作为判定准则,能够耐受非资源胁迫的外来种个体才能定植。Richards et al(2006)的研究强调了表型可塑性与环境条件随机匹配在入侵性中的效应,该研究发现:当低可塑性物种随机扩散至适宜的环境,适合度高;当高可塑性物种扩散至次好的环境,适合度低。这样,环境质量和表型可塑性等确定力量,以及扩散等随机力量决定了外来种个体定植的可能性。
定植后,外来种个体数量的增加速度受到土著种个体数量减少速度的深刻影响。土著种个体数量的减少可以由个体随机死亡引起,也可以由种间竞争、土著消费者和土著病原体引起。根据Melbourne et al(2007)和Zhou et al(2021)的研究结果,结合确定力量环境异质性以及物种的扩散可以更清晰地认知外来种个体增加的可能性。环境异质性高,意味着环境差异大,外来种偏好生境的面积小,限制了种群的规模。相反,环境异质性低,即匀质性高的区域,外来种可能受到更多土著种的强烈的竞争排斥。
土著种个体死亡后,形成了空闲的资源,这些资源能否被外来种繁殖体占据进而增加个体数量,取决于繁殖体产生能力的物种间差异以及个体生长能力的物种间差异。因此,外来种个体更高的生长速度和繁殖能力,将是土著群落可入侵性重要的确定力量。
外来种个体到达、定植、个体增加伴随着土著种个体的减少,这个土著群落被入侵过程中的每一个重要环节都由随机和确定力量共同决定。通过调查、实验、模拟,整合生态位理论和中性理论,理解和预测群落的可入侵性及其控制因素,将为预测预警预防生物入侵的工作提供必要的理论基础,是降低外来种危害我国生态安全、经济社会发展的重要保障。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
落叶果树(2020年2期)2020-04-20
环球时报(2017-10-10)2017-10-10
中国新闻周刊(2015年31期)2015-08-28
岁月(2009年10期)2009-10-28
