
江仓矿区高加索三叶草引种试验
2022-04-27 04:46陈善晶谢久祥欧为友
青海草业 2022年1期
贾 萍,牟 丹,陈善晶,曾 心,王 娟,谢久祥,欧为友
(1.青海大学农牧学院,青海 西宁 810016;2.青海省草原总站,青海 西宁 810008)
青海省木里江仓矿区位于青海省海西州天峻县和海北州刚察县境内,是青海省最大的煤矿,也是优质焦煤和可燃冰资源整装勘查矿区。江仓矿区的植被类型以天然高寒草地和湿地为主,畜牧业是当地的主导产业,是青海省发展牧业生产自然环境条件较好地区之一[1]。近年来随着国民经济的快速发展,该地区地下储存的煤炭资源遭到掠夺式、粗放型开发,导致当地的原生植被遭到严重破坏,加剧恶化了当地的生态环境,同时也让被誉为“中华水塔”的青藏高原宝贵的湿地资源流失,这与国家近年来提出的“绿水青山就是金山银山”的生态文明建设不符。2014年以来国家和青海省政府加大资金扶持力度,对该地区的露天煤矿区进行大规模的渣山植被恢复,已经得到了初步成效。高寒矿区海拔高,年积温低,极端的生态环境对于植物的生长极为不利,目前有关矿区渣山植被恢复的研究报道较少[2]。
在2014年至2021年进行的对青海木里和江仓煤田渣山进行的植被修复中,所使用的草种为同德短芒披碱草(Elymusnutans)、青海草地早熟禾(Poapratensis)、青海冷地早熟禾(Poacrymophila)和青海中华羊茅(Festucasinensis)等4种禾本科牧草[3]。以垂穗披碱草、冷地早熟禾、中华羊茅等禾本科牧草为主的多年生人工草地具有1年建植多年受益的特点,是青藏高原主要的多年生人工草地推广栽培方式。但是,生产实践中发现,这种草地普遍存在建植3~5年后急速退化的现象[4~8],成为进一步示范推广的严重障碍。建植人工草地是恢复“黑土滩”草地的重要措施,但是由于管理和利用不当,3~5年后再次成为“黑土滩”[9]。原因在于垂穗披碱草等人工引进牧草,极大的消耗了土壤肥力,人工草地在退化后并没有改善“黑土滩”的土壤质量,可能造成“黑土滩”二次发生[9,10]。改建黑土滩所用草种单一,也是引起黑土滩人工草地退化的原因之一。多年生人工草地建植和管理中,豆禾混播是最理想的组合[11]。不过,青藏高原高寒牧区适宜栽培的豆科牧草极度缺乏[12],外来草种的引进又难以成功[13]。
近20年来,科学家在青海高寒地区进行了大量的豆科牧草引种试验,在海拔2500 ~3000 m的地区,引种成功的报道很多,如紫花苜蓿[14~17]、截形苜蓿和波斯三叶草[18]、黄花草木樨[11]、红豆草和甘农1号杂花苜蓿[19]等;在海拔3000~3300 m的地方,引种成功的报道很少,如甘肃红豆草和甘农1号杂花苜蓿[19,20],青大1号紫花苜蓿(王晓勇等,2012);在海拔3300 m以上的地方,虽有几次尝试,均以失败告终。2015年以来,青海大学在河南县柯生乡和优干宁镇海拔3600~3700 m的高寒草地引种高加索三叶草,发现其可在当地成功越冬并依靠地下茎实现扩繁而多年存活[21]。本研究中,我们将高加索三叶草引种到江仓矿区,并观察其越冬和扩繁情况,为高寒矿区植被恢复过程中的草种选择提供参考依据。
1 研究区概况与试验材料
1.1 研究区概况
青海木里江仓矿区地处青藏高原东北端的祁连山中段南部地区,是大通河的上游盆地,因此该地区露天煤矿的开采严重影响大通河(湟水重要支流)的水质。该地区年最高气温为19.8 ℃,最低气温可达-34 ℃,年平均气温为-4.2~-5.1 ℃。年平均降水量为314 mm,蒸发量为1 418 mm,降雨主要集中在7~9月,降雪集中在1~5月。矿区四季多风,夏季盛行东南风,冬季盛行西北风。矿区海拔高度为3800~4000 m ,属于高寒草甸低位沼泽湿地,地势起伏不大,多年冻土发育,地表冻融草沼、热融湖沼、冻胀丘等地貌形态发育。矿区天然高寒草甸和湿地保存完好,土壤肥沃,植被密度很大,植物种类较多,是青海省东北部较大的天然生态保障屏障。植被构成以寒冷中生、短根茎的嵩草属植物为主,具有植株低矮密丛、贴地面生长等耐寒特征,层次分化不明显,主要优势种有高山嵩草(Kobresiapygmaea)、线叶嵩草(Kobresiacapillifolia)、矮生嵩草(Kobresiahumilis)、垂穗披碱草(ElymusnutansHriseb.)等。
1.2 试验材料
2015年,新西兰林肯大学James Moir教授将高加索三叶草引种到江仓矿区,受鼠害破坏后,我们将残留的部分植株于2018年5月移栽至西宁市城北区。2018年,由于种苗不够,我们从内蒙古农业大学王明玖教授处采集了高加索三叶草育成品种“蒙农三叶草1号”(TrifoliumambiguumBieb. cv. Mengnong No.1)的种子和地下根茎,取50株移栽至江仓矿区。
1.3 试验方法
1.3.1 试验材料种植 2019年6月,将“蒙农三叶草1号” 高加索三叶草根茎移栽至江仓矿区试验地,进行单株移栽试验。小区移栽时,行距30 cm,株距30 cm。本研究用于引种观测的高加索三叶草种植年限为2019年至2021年。
1.3.2 引种表现观测 观测引种后的高加索三叶草的生长表现,如物候期、长势、生物量、种子产量和结瘤情况等指标。
(1)越冬率:随机挑选3个小区,分别计数越冬前后植株数,并计算越冬后(即返青期)与越冬前(即冻萎期)的比值。
(2)物候期:分为返青期、分枝期、现蕾期、开花期、结荚期和成熟期,均以50%植株进入该物候期计。
(3)冠幅:指高加索三叶草植株的在南北和东西方向宽度的平均值,用于表示其植株的大小。
(4)株高测定:当高加索三叶草进入盛花期时,随机选6株测定株高(包括自然高度和绝对高度)。
(5)分蘖数:随机选取6株并计数,早春以从根颈处萌出嫩芽数为准,盛花期以从地面长出枝条数为准。
(6)地上茎直径:用游标卡尺量取地上茎直径,随机量取6株,每株3枝。
(7)地上生物量鲜重:在高加索三叶草盛花期,随机挑选3个小区,在各小区内随机选取1株或1个1 m×1 m样方,齐地面刈割,称鲜重,分别记为单株地上生物量鲜重和单位面积地上生物量鲜重。
(8)地上生物量干重:获取地上生物量后烘干至恒重,即得干重。
(9)鲜干比:鲜重与干重的比值,即单株鲜干比=单株地上生物量鲜重/单株地上生物量干重;单位面积鲜干比=单位面积地上生物量鲜重/单位面积地上生物量干重。
(10)地下生物量干重:在高加索三叶草盛花期,随机挑选3个小区,在各小区内随机选取1个1 m×1 m样方,采挖地下部分,洗净,烘干至恒重。
(11)叶量比:指牧草或饲料作物在植物构成中,叶占地上生物量的比重。随机挑选6 株,齐地面刈割后,分离叶片,风干后分别称取叶重和总重,计算叶与总重的比值。操作时将叶柄、嫩稍均计入叶中。
(12)根茎情况:随机挖出6株,记录轴根数、侧根数、根茎数,量取每株侧根长、轴根长、根茎长、根茎直径,将地下部分洗净后,称取鲜重、干重。
(13)根瘤发育情况:随机挖出6株,观察根瘤的颜色,计数每株根瘤数,每取50粒称取根瘤鲜重,三次重复,烘干后分别称其干重。
1.4 数据分析
采用Excel 2016整理数据与绘制图形,采用SPSS 20.0进行统计分析,三年间平均值差异显著性检验用单因素方差分析方法。
2 结果与分析

表1 江仓矿区引种高加索三叶草的物候期
2.1 物候期观测
引种当年,高加索三叶草在江仓矿区不能开花,绿期129 d;引种第三年,高加索三叶草在江仓矿区不能开花,绿期164 d;引种次年,高加索三叶草少部分植株开花,但种子仍不能成熟,绿期165 d。
2.2 越冬率与无性繁殖情况观测
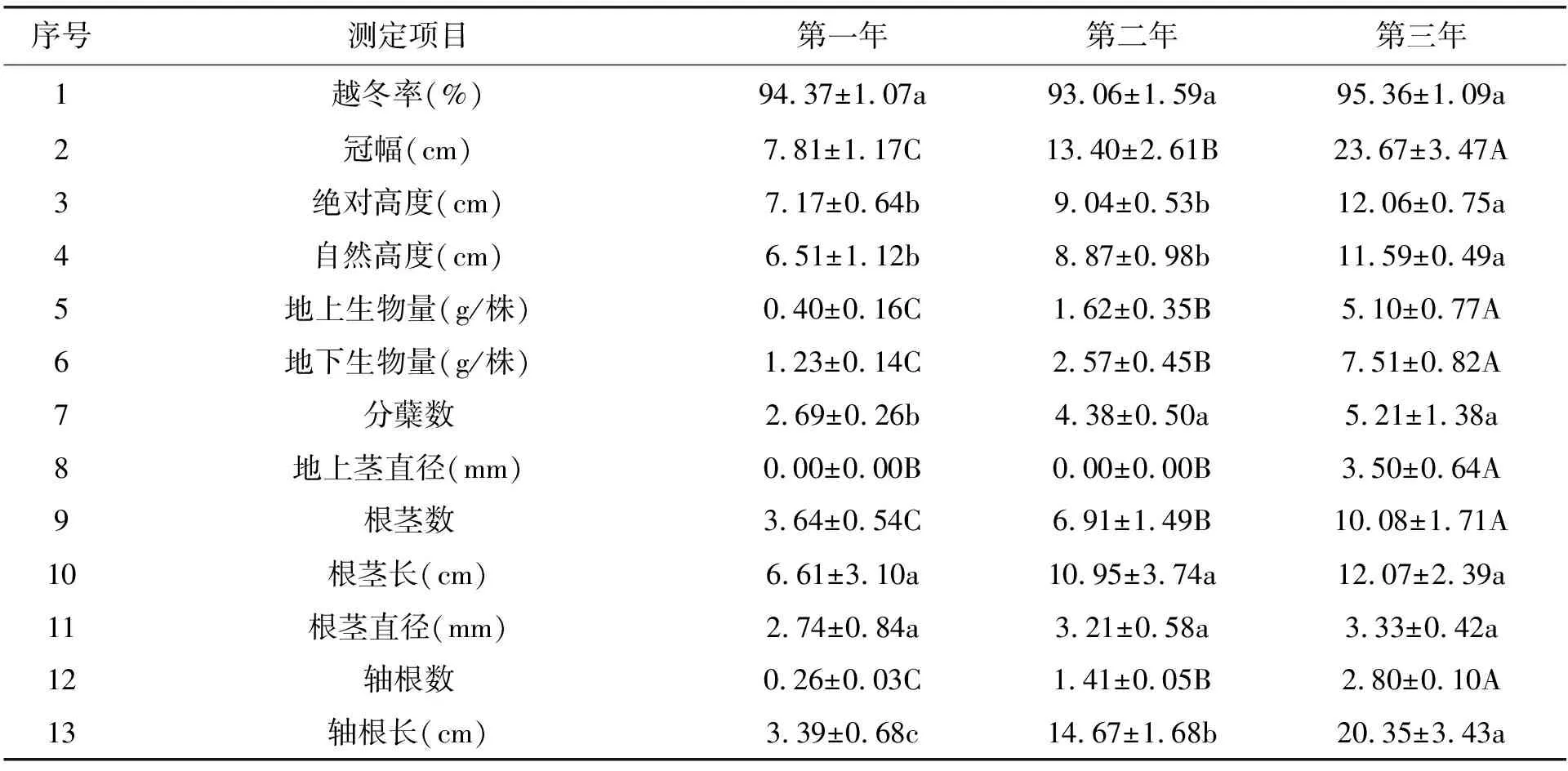
表2 江仓矿区高加索三叶草引种前三年表现
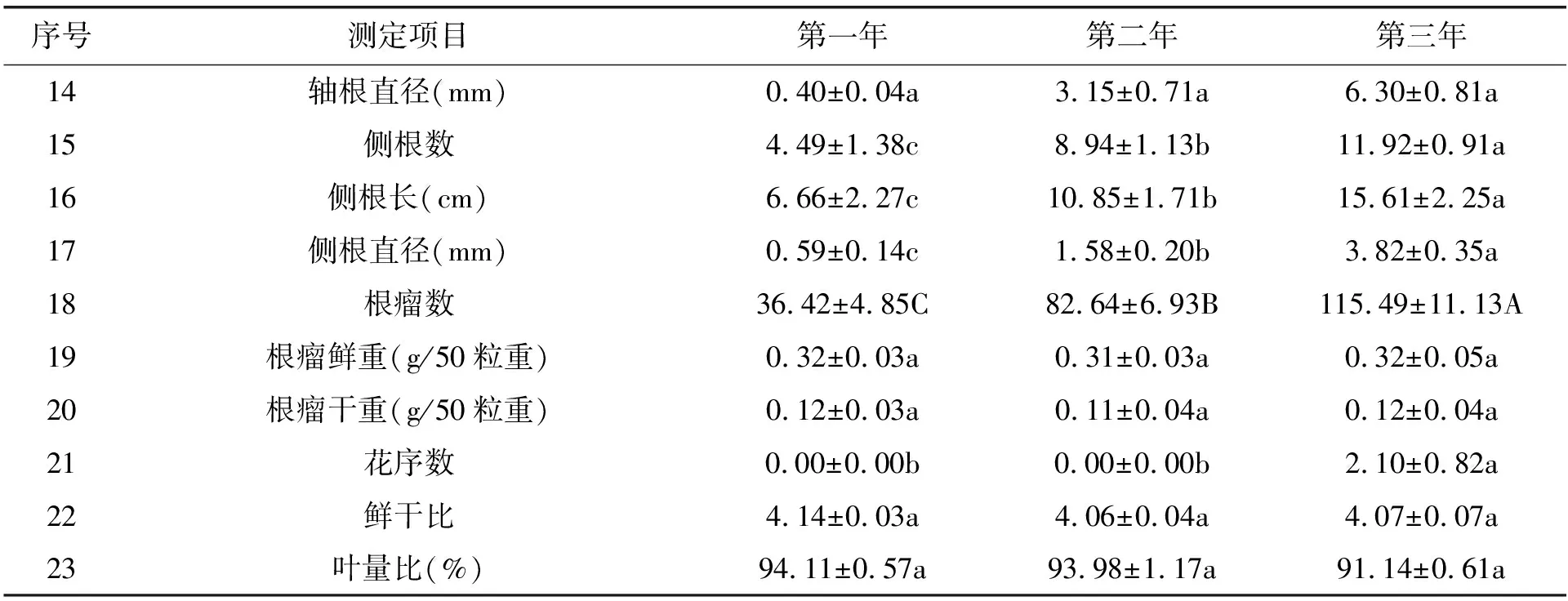
续表:
在高寒地区,影响高加索三叶草存活的主要因素为越冬困难。为此,我们测定了高加索三叶草在江仓矿区的越冬率。高加索三叶草以根茎发达著称,我们测定了根茎情况,包括根茎数、根茎长、根茎粗等指标。不论在引种当年,还是在引种第二和第三年,高加索三叶草的在江仓的越冬率均在93%以上,各年份之间差异不显著(表2,P>0.05)。说明高原冬季低温、干旱和强风对其很小影响,原因在于高加索三叶草地下根茎发达,地下根茎出芽点埋藏在地下3~4 cm深度。此外,试验地经过鼠害防控处理,免遭害鼠破坏,仅有几株死亡,也是由于偶发害鼠破坏引起的。根茎数随着引种年份的增加而显著增加,几乎呈直线增加趋势,引种各年份之间差异极显著(表2,P<0.01)。说明高加索三叶草在江仓矿区能稳定形成地下根茎,这对于高加索三叶草的无性繁殖具有重要意义。引种第一年,高加索三叶草在江仓矿区的根茎长度为6.61±3.10 cm,在引种第二和第三年,高加索三叶草的根茎长均达到10 cm以上,三年之间差异不显著(表2,P>0.05),说明高加索三叶草不仅地下根茎发达,而且地下根茎的长度在引种第二年后显著增加,第二年和第三年后长度在10 cm以上。高加索三叶草引种到江仓矿区的高加索三叶草的根茎直径在各年份之间差异不显著(表2,P>0.05)。根茎粗度发育良好,为其进一步形成的地下出芽点打下基础,有利于形成分蘖。
2.3 株形特征观测
高加索三叶草的冠幅在移栽后的三年之间差异极显著(表2,P<0.01),其冠幅依次为7.81±1.17 cm、13.40±2.61 cm、23.67±3.47 cm,年增长率约为200%,说明高加索三叶草凭借其地下根茎,能在江仓矿区快速扩繁,占领领地,形成植被。
移栽后的三年中,高加索三叶草的分蘖数在各年份之间差异显著(表2,P<0.05),移栽后的三年中,江仓矿区高加索三叶草的分蘖数逐年增加,形成了庞大的地下根系和根茎系统,储存了大量有机物,为分蘖的形成和返青做好了物质准备。
高加索三叶草的自然高度在三年之间差异显著(表2,P<0.05),第一年和第二年的自然高度平均值在5.39~9.85 cm之间,均贴地生长形成垫状植物,没有地上茎的形成;至第三年,由于有少量花序形成,地上茎长度达到11.59 cm左右,显著高于前两年。
高加索三叶草的绝对高度在三年之间差异显著(表2,P<0.05),第一年和第二年的绝对高度平均值在6.53~9.57 cm之间,均贴地生长形成垫状植物,没有地上茎的形成;至第三年,由于有少量花序形成,地上茎长度达到12.06 cm左右,显著高于前两年。
移栽后的三年中,江仓矿区高加索三叶草的地上茎直径存在极显著差异(表2,P<0.01)。原因在于,高加索三叶草至第三年才发育出了地上茎,第一年和第二年没有地上茎的形成。第三年地上茎直径为3.5 mm左右。
高加索三叶草虽然以根茎型植物著称,但仍具有轴根。实生苗具有轴根,而移栽苗在移栽成功后也会逐渐生长出轴根。轴根不仅提高其固着力,而且能帮助其吸收深层土壤水分和养分。移栽后的三年间,高加索三叶草的轴根数差异极显著(表2,P<0.01),三年的轴根数平均值分别为0.26±0.03根/株、1.41±0.05根/株、2.80±0.10根/株。移栽后的第一年,江仓矿区的高加索三叶草几乎不能形成轴根,第二年至第三年,高加索三叶草的轴根数显著增加,多者达到3根。
移栽后的第一至第三年,高加索三叶草的轴根长平均值分别为3.39±0.68 cm、14.67±1.68 cm、20.35±3.43 cm,说明高加索三叶草的轴根长度在三年内保持了很强的增长态势。
移栽后的三年内,高加索三叶草的侧根数差异显著(表2,P<0.05),第一至第三年的侧根数平均值分别为4.49±1.38根/株、8.94±1.13根/株、11.92±0.91根/株,说明高加索三叶草的侧根数在三年内保持了很强的增长态势。
移栽后的三年内,高加索三叶草的侧根长差异显著(表2,P<0.05),第一至第三年的侧根长分别为6.66±2.27 cm、10.85±1.71 cm、15.61±2.25 cm,说明高加索三叶草的侧根长在三年内保持了很强的增长态势。
2.4 单株饲草生产性能测定
移栽后的第一至第三年,高加索三叶草的单株地上生物量(干重)平均值分别为0.40±0.16 g/株、1.62±0.35 g/株、5.10±0.77 g/株,三年之间差异极显著(表2,P<0.01),说明高加索三叶草的单株地上生物量在三年内保持了很强的增长态势。
移栽后的第一至第三年,高加索三叶草的单株地下生物量(干重)差异极显著(表2,P<0.01),其平均值分别为1.23±0.14 g/株、2.57±0.45 g/株、7.51±0.82 g/株,说明高加索三叶草的单株地下生物量在三年内保持了很强的增长态势。
移栽后的第一至第三年,高加索三叶草的鲜干比没有显著性差异(表2,P>0.05),均维持在4.05左右,说明高加索三叶草的鲜干比总体稳定。
移栽后的第一至第三年,高加索三叶草的叶量比差异不显著(表2,P>0.05),三年的叶量比平均值分别为94.11±0.57%、93.98±1.17%、91.14±0.61%,原因在于三年之内,高加索三叶草在江仓矿区均为垫状植物,前两年没有生殖枝的形成,至第三年才有少量生殖枝形成。
2.5 结瘤性能观测
移栽后的第一至第三年,高加索三叶草的每株根瘤数差异极显著(表2,P<0.01),每株根瘤数平均值分别为36.42±4.85个/株,82.64±6.93个/株、115.49±11.13个/株,说明在移栽到江仓矿区之后,高加索三叶草的根瘤发育随着地下生物量的增长而增长,结瘤能力基本稳定,根瘤数目显著增长。
移栽后的第一至第三年,高加索三叶草的绝大部分(>90%)根瘤均为淡红色,掐开后断面呈粉红色,高加索三叶草的根瘤鲜重差异不显著(表2,P>0.05),均维持在0.32 g/50粒左右,其根瘤干重也无显著差异(表2,P>0.05),均维持在0.12 g/50粒左右。
3 讨论
引种到江仓矿区后,高加索三叶草在第一年和第二年不能开花,在引种后的第三年才能少量开花,但种子不能成熟。不过,凭借强大的根孽和地下根茎扩繁能力,高加索三叶草在江仓矿区可实现无性扩繁,这也在一定程度上说明,高加索三叶草在江仓矿区的引种试验是成功的。
3.1 高加索三叶草在青海各地引种的生育期和绿期
据殷秀杰[22]在内蒙多地的引种报道,高加索三叶草生育期较短,绿期长。高加索三叶草引种到江仓矿区后,返青比当地牧草早约10 d,枯黄晚约10 d,总绿期比当地牧草长约20 d。在如此漫长的绿期内,高加索三叶草在江仓矿区不能实现种子成熟的原因,在于积温不够。为此,如果在开花前对其进行覆膜处理,增加积温,形成花蕾后去掉薄膜,任由当地昆虫传粉,应当可以实现结实和成熟,具体情况有待进一步研究。在江仓矿区,高加索三叶草的绿期长于当地的无霜期,另外,据牟丹[21]报道,高加索三叶草引种到西宁市城北区后,绿期为249 d,无霜期为100 d,绿期比无霜期长149 d;引种到西宁市湟中区后,绿期为216 d,无霜期为91 d,绿期比无霜期长125 d;在河南县,绿期为166 d,无绝对无霜期,可以认为绿期比无霜期长166 d。可以认为,高加索三叶草在较长时间的霜期内,可以正常生长,保持绿色,其抗霜冻能力较强,对于青藏高原植被修复具有重要意义。
3.2 高加索三叶草在江仓矿区的生长特性和无性繁殖特性
高加索三叶草在江仓矿区的引种过程中,引种第三年才有少部分植株开花,种子不能成熟。借助其根孽和地下根茎的扩张优势,其冠幅每年可扩张10 cm以上。引种后三年内,高加索三叶草的冠幅、分蘖数、根茎数、地上生物量、地下生物量、轴根数、轴根长、轴根直径、侧根数、侧根长等表征其生长特性和无性繁殖特性的指标均显著增长。绝对高度、自然高度、地上茎直径、根茎直径、鲜干比和叶量比等指标三年内差异不显著,保持稳定,说明引种到江仓矿区的三年内,高加索三叶草在植株生长特性、无性繁殖等方面均表现良好,适合于在江仓矿区引种、扩繁。
牧草产量方面,移栽后的第一至第三年,高加索三叶草的单株地上生物量和地下生物量均极显著增长,但因为移栽株数只有50株,种植面积过小,本研究无法计算高加索三叶草在江仓地区的单位面积牧草产量和地下干物质产量。张玉发[23]在北京引种高加索三叶草后,在种植的第二年测得,亩产干草273.9 kg,这相当于牟丹[21]在西宁市城北区引种后测得的第一年的亩产干草产量(280.05 kg)。据报道,高加索三叶草的地上生物量从第二年开始增加,在7~8 年达到高峰[24]。估计在后续年份,引种在江仓矿区高加索三叶草的地上和地下产量会有更好的表现。据Strachan et al.报道[25],在种植第13年时,高加索三叶草的地下生物量为20 t/hm2,磷、氮和硫的含量分别达58 kg/hm2、455 kg/hm2和20 kg/hm2,以及数量更庞大的碳水化合物。由此看来,如果在青藏高原扩大高加索三叶草的种植范围,既可通过生物固氮改良草地,对实现碳达峰和碳中和也具有重要的意义。
3.3 高加索三叶草在青海各地的结瘤性能
一般认为,高加索三叶草不能形成根瘤进行生物固氮,这成为限制其引种的主要担忧之一[26]。以往的做法是通过专一根瘤菌接种形成根瘤,或通过和其他三叶草杂交育种,使其具备固氮能力[27];或与其他豆科牧草混播后,进行氮替换。引种到国内后,高加索三叶草在赤峰和呼和浩特等地的结瘤很少[22]。但是,据牟丹的研究[21],引种到西宁市城北区、湟中区和河南县之后,高加索三叶草的结瘤均十分丰富。本研究中,引种到江仓矿区后,第一年和第二年的株均根瘤数分别约为36.42±4.85个和82.64±6.93个,至第三年达到115.49±11.13个,结瘤量十分丰富。三年的根瘤数大体与地下生物量成正比,不存在不结瘤的植株。另外,根瘤洗净后成淡粉色,在破开后为粉红色,均为有效根瘤。因此,引种高加索三叶草,对于青藏高原高寒草地的固氮具有重要意义。
猜你喜欢
中国种业(2022年4期)2022-04-19
中国南方果树(2022年1期)2022-01-28
农技服务(2021年7期)2021-09-24
当代世界(2021年1期)2021-01-20
蔬菜(2020年8期)2020-12-16
椰城(2019年11期)2019-12-09
北方文学(2019年18期)2019-07-25
幼儿教育·父母孩子版(2017年3期)2017-06-15
文艺生活·中旬刊(2016年8期)2016-05-14
诗歌月刊(2014年7期)2015-04-21
