
烤烟叶片叶绿体超微结构与质体色素降解产物的关系初探
2022-04-26 06:49王玉李春光刘欢张月华冯晓民李耀光李怀奇景延秋孙觅
中国农业科技导报 2022年3期
王玉, 李春光, 刘欢, 张月华, 冯晓民, 李耀光,李怀奇, 景延秋*, 孙觅*
(1.河南农业大学烟草学院,郑州 450002;2.河南中烟工业有限责任公司技术中心,郑州 450002;3.河南中烟工业有限责任公司黄金叶生产制造中心,郑州 450002)
叶绿体普遍存在于植物细胞中,是进行光合作用的重要细胞器[1],与烟叶的生长发育和烤烟内在品质的形成关系密切[2]。质体色素降解产物是决定烟叶挥发性香味物质总量高低的关键致香成分[3-4]。烟叶叶绿体中的质体色素包括叶绿素和类胡萝卜素,主要存在于类囊体膜上,它们的合成和积累与光合作用具有紧密联系[2]。不同部位的烟叶由于生长条件不同,化学成分含量差异较大,质体色素降解产物含量的差异与其前体物质含量的差异有关,反应了叶绿体超微结构的变化。因此,研究烤烟不同部位叶片叶绿体超微结构差异对质体色素降解产物含量的影响具有重要意义。任春琼等[5]研究发现,随着叶位的升高,叶绿体中淀粉颗粒与嗜锇颗粒的积累量增多,基粒片层断裂程度加重,叶绿体结构破损更加剧烈。云菲等[6]研究表明,光氮互作效应对叶绿素及类胡萝卜素降解产物有极显著影响,适度弱光和增加施氮量有利于质体色素降解产物的合成和积累。刘典三等[7]研究发现,烟叶的类胡萝卜素降解产物含量随叶位的升高而增加,降低光照强度后,质体色素降解产物的总含量减少。张国显等[8]研究表明,随着叶位的升高,烤烟叶绿素与类胡萝卜素的降解产物含量均表现为先增加后降低趋势。目前,关于烤烟叶片叶绿体微观结构和质体色素降解产物含量的研究多集中在单一方面,而对于烤烟叶片叶绿体结构差异与质体色素降解产物含量关系的研究较少。因此,本研究以云烟87为试验材料,通过对不同部位未熟和适熟的烟叶叶绿体超微结构进行观测,进一步探讨其与质体色素降解产物含量的关系,旨在从叶片细胞超微结构角度解释不同部位间质体色素降解产物含量差异的成因,为研究烟叶香味品质的形成提供新的研究途径与理论依据。
1 材料与方法
1.1 试验材料
试验以云烟87为供试品种,于2020年在河南省三门峡市灵宝市的示范烟田(E 110°41′,N 34°21′)进行。试验地烟草生长季平均气温21.69℃,降水量329.43 mm,日照总时数817.6 h。烟苗于2020年5月10日(苗龄70 d左右)采用膜下小苗移栽方式进行移栽。试验田肥力中等,土壤基本理化性质为:pH 7.19,碱解氮70.56 mg·kg-1、速 效 磷 8.29 mg·kg-1、速 效 钾 159.47 mg·kg-1、有机质 15.82 g·kg-1。
1.2 试验设计
试验采用随机区组设计,3次重复,共6个小区,周围设置保护行,各小区均栽植烟株360株,行距120 cm,株距50 cm,按照田间优质烟叶管理方法进行管理,烟田施氮量为65 kg·hm-2。试验于2020年8—9月分别取下部叶(lower leaves,LL;叶位 5~7)、中部叶(middle leaves,ML;叶位 10~12)和上部叶(upper leaves,UL;叶位16~18)的未熟和适熟2种成熟度烟叶,不同部位不同成熟度的鲜烟叶外观判定标准[9]详见表1。对应成熟度和叶位范围在每个小区选取完整、大小一致、具有代表性的鲜烟叶5片用于透射电子显微镜观测叶绿体超微结构;并取该批次的上、中、下3个部位的初烤烟叶样品各1 kg进行质体色素降解产物的检测分析。
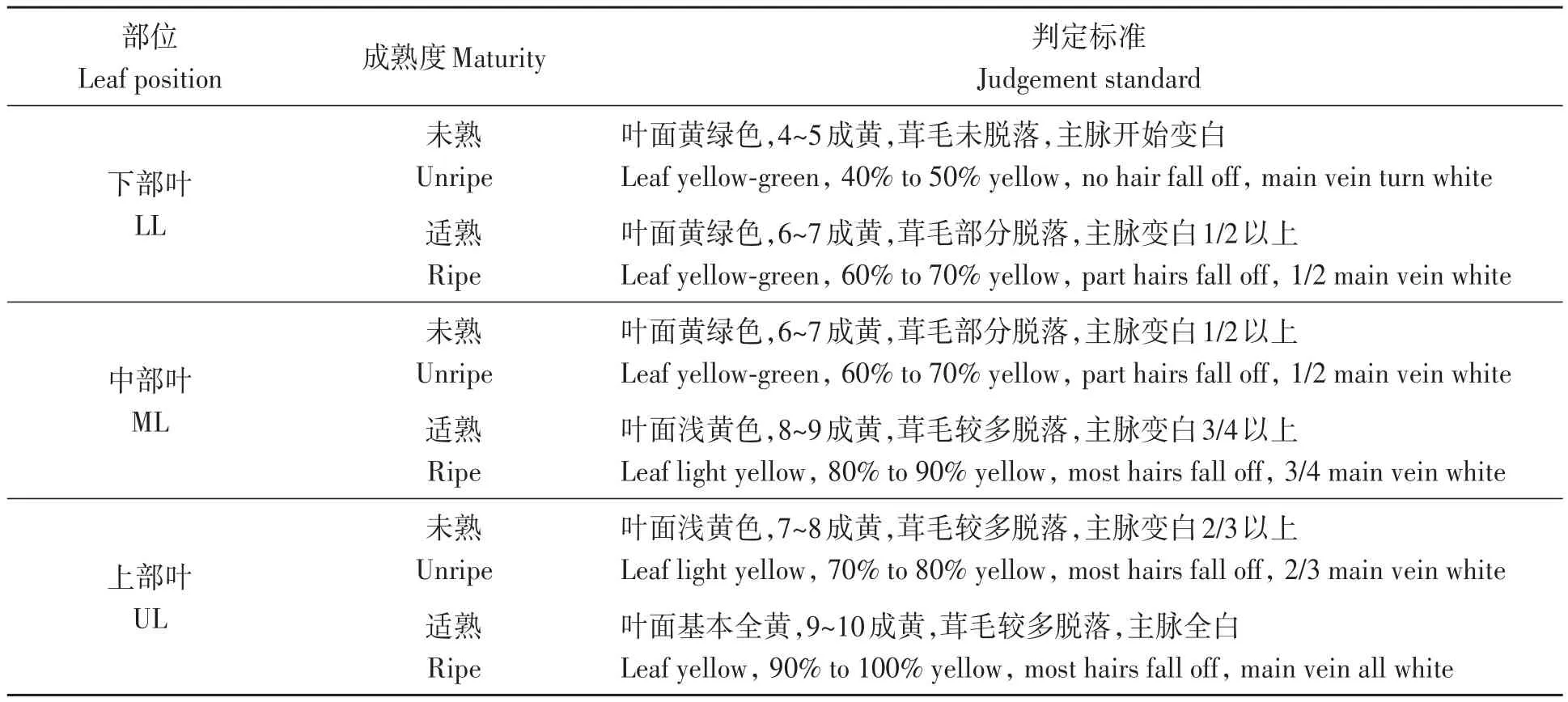
表1 不同部位烟叶成熟度标准Table 1 Criteria for maturity level of flue-cured tobacco from different leaf position
1.3 试验方法
1.3.1 烟叶透射电镜样品的制备与观测 在鲜烟叶的叶中部避开主侧脉切取1 mm×2 mm的小片,立即放入4%戊二醛固定液(pH 7.2)中,于4℃保存,之后再用1%锇酸固定。然后按照常规超薄切片法进行脱水、渗透、包埋、聚合、切片和染色(醋酸双氧铀-柠檬酸铅双重染色)[5],再用JEM-1400透射电子显微镜(日本)观测并拍照,加速电压100 kV,每个处理观察10个视野。
1.3.2 烤烟中性香气成分含量的测定 将20 g初烤烟叶的烟末样品置于蒸馏萃取装置提取香味物质,提取液经由二氯甲烷萃取后,于旋转蒸发仪上浓缩至1 mL进行气相色谱-质谱鉴定。采用HP6890-5975气质联用仪(美国)定性测定,采用乙酸苯乙酯溶液作内标进行定量测定[10]。各样品均重复3次,取平均值。
依据各中性香气成分定性和定量测定结果,统计叶绿素降解产物含量(chlorophyll degrading products content,CHLDPC)、类胡萝卜素降解产物含 量 (carotenoid degrading products content,CARDPC)、中性香气成分总量(neutral aroma component contents,NACC),其中,中性香气成分中叶绿素降解产物主要为新植二烯[6],因此,以新植二烯含量(neophytadiene content,NC)代表叶绿素降解产物含量。
采用以下公式计算叶绿素降解产物新植二烯占比(Pa)和类胡萝卜素降解产物占比(Pb)。

1.4 数据处理
运用Microsoft Excel 2010进行数据的统计整理,运用Adobe Photoshop CC 2019进行图片整理,运用SPSS 25.0进行方差分析和相关性分析。根据Goodenough等[11]改进的Teichler-Zallen法对每个样品的叶绿体基粒片层进行统计。
2 结果与分析
2.1 不同部位未熟和适熟烟叶的叶绿体超微结构特征
从图1A~C可看出,未熟的上、中、下部叶中叶绿体均具有较完整的双层膜、基粒和基质片层结构,且基粒中垛叠的类囊体个数较多。上部叶的淀粉颗粒体积最大,多数呈不规则形状,单个叶绿体中的基粒数与嗜锇颗粒数最多,垛叠层数较厚,整体结构紧贴细胞壁(图1A);中部叶的淀粉颗粒呈椭圆形,积累量和嗜锇颗粒数较上部叶少,单个叶绿体中的基粒数和类囊体垛叠程度较少(图1B);下部叶的淀粉积累量和嗜锇颗粒数最少,体积最小,淀粉颗粒呈长椭圆形(图1C)。综上所述,随着叶位的上升,未熟烟叶淀粉颗粒和嗜锇颗粒的积累量逐渐增多,叶绿体整体结构逐渐贴近细胞壁,叶绿体结构的完整程度逐渐下降。
从图1D~F可看出,适熟的上、中、下部叶中都积累了大量的淀粉颗粒和嗜锇颗粒,且数量较相同部位的未熟叶片多,而基粒和类囊体个数较未熟叶略少。上部叶的叶绿体外膜破损严重,淀粉颗粒向细胞中部游离,多数呈不规则形状,嗜锇颗粒含量最多(图1D);中部叶的叶绿体结构相对上部叶完整,淀粉颗粒呈不规则形状(图1E);下部叶的叶绿体结构最为完整,淀粉颗粒体积最小,呈长椭圆形(图1F)。中部叶和下部叶的叶绿体结构紧贴细胞壁排列。综上所述,随着叶位的升高,适熟烟叶叶绿体结构的解体程度逐渐增加;基粒片层结构松散、解体加剧,类囊体垛叠程度降低;嗜锇颗粒和淀粉颗粒数量增多,体积增大。

图1 不同部位未熟和适熟烟叶的叶绿体结构特征Fig.1 Chloroplast ultrastructure character in unripe and ripe leaves of different leaf positions
2.2 不同部位未熟和适熟烟叶的叶绿体超微结构统计
由表2可知,不同部位未熟和适熟烟叶的叶绿体中淀粉颗粒、嗜锇颗粒和基粒的含量差异显著;随着叶位的升高,叶绿体中淀粉颗粒和嗜锇颗粒的含量呈现逐渐增多的趋势。未熟烟叶叶绿体的基粒数表现为:上部叶>中部叶>下部叶;但适熟烟叶叶绿体的基粒数随着叶位的升高而减少。随着成熟度的提升,各部位烟叶叶绿体的淀粉颗粒、嗜锇颗粒含量逐渐增多;上、中部烟叶的基粒数随之减少,下部叶的基粒数随之增多。

表2 不同部位未熟和适熟烟叶单个叶绿体的超微结构统计Table 2 Statistics of chloroplasts ultrastructure in flue-cured tobacco leaves of different positions at unripe and ripe stages
由图2可知,在未熟烟叶中,上部叶中存在片层数在30以上的高基粒片层;中部叶和下部叶的高基粒片层还未出现。上部叶中基粒片层数在20~40间的占比(51.06%)显著高于中部叶和下部叶;中部叶中基粒片层数在16~30间的占比(62.42%)显著高于下部叶;下部叶中基粒片层数在15以下的占比(72.05%)较大。由此表明,未熟烟叶中高叶位叶绿体中高基粒片层数占比较高,基粒片层的垛叠程度较高。

图2 不同部位未熟烟叶叶绿体的基粒片层数Fig.2 Grana lamellae number in chloroplasts of unripe leaves at different leaf positions
由图3可知,适熟烟叶的上部叶中高基粒片层已经消失,而中部叶和下部叶出现了片层数高于30的高基粒片层。下部叶中基粒片层数在20~40间的占比(46.66%)显著高于中部叶(22.43%)和上部叶(7.43%);中部叶中基粒片层数在10~20间的占比最高(64.11%);上部叶中基粒片层数在10以下的低基粒片层占比最大(66.23%)。由此表明,随着成熟度的增加,烟叶叶绿体基粒片层的垛叠程度下降,基粒片层结构解体程度加剧。

图3 不同部位适熟烟叶叶绿体的基粒片层数Fig.3 Grana lamellae number in chloroplasts of ripe leaves at different leaf positions
2.3 不同部位适熟烟叶叶绿素降解产物含量的差异
由图4可知,成熟的烟叶烘烤后,不同部位间新植二烯(叶绿素降解产物)的含量差异显著。随着叶位的升高,新植二烯含量呈逐渐增加趋势,表现为上部叶>中部叶>下部叶。其中,上部叶的新植二烯含量占中性香气成分总量的80%以上,显著高于中部叶和下部叶;中部叶和下部叶的新植二烯占比差异不显著,均为70%左右。

图4 不同部位适熟烟叶的新植二烯含量及占比Fig.4 NC content and proportion in ripe flue-cured tobacco leaves of different leaf positions
2.4 不同部位适熟烟叶的类胡萝卜素降解产物的含量差异
由图5可知,类胡萝卜素降解产物总量随着烤烟叶位的升高而增加。不同部位间差异显著,表现为上部叶>中部叶>下部叶,与叶绿素降解产物含量变化一致。类胡萝卜素降解产物占中性香气成分总量的比例在上部叶和中部叶中为10%左右,显著低于下部叶。
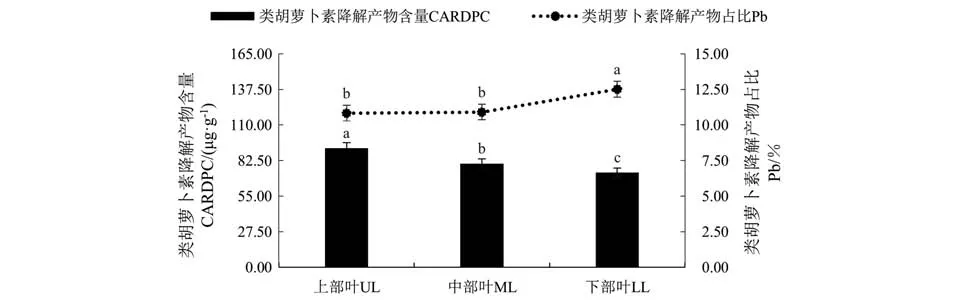
图5 不同部位适熟烟叶类胡萝卜素降解产物含量Fig.5 CARDPC content and proportion in ripe flue-cured tobacco leaves of different leaf position
由表3可知,上、中、下部叶的β-大马酮含量占类胡萝卜素降解产物总量的比例最高,为19.33%~23.19%;上部叶的6-甲基-5-庚烯-2-醇、6-甲基-5-庚烯-2-酮、β-大马酮、β-二氢大马酮、二氢猕猴桃内脂含量显著高于中部叶和下部叶;巨豆三烯酮的4个同分异构体和螺岩兰草酮的含量以中部叶最高,显著高于上部叶和下部叶;下部叶氧化异佛尔酮的含量显著高于上部叶和中部叶。

表3 不同部位适熟烟叶的香味物质含量Table 3 Contents of aroma constituents in ripe flue-cured tobacco leaves of different leaf position (μg·g-1)
2.5 烟叶的质体色素降解产物含量与叶绿体超微结构指标的相关性分析
由表4可知,未熟和适熟烟叶中单个叶绿体的淀粉颗粒、嗜锇颗粒与叶绿素和类胡萝卜素降解产物的含量均呈极显著正相关。单叶绿体中基粒数与未熟烟叶的类胡萝卜素降解产物和新植二烯含量呈极显著正相关,而与适熟烟叶中类胡萝卜素降解产物和新植二烯含量呈极显著负相关。

表4 烟叶单个叶绿体超微结构指标与质体色素降解产物含量的相关性分析Table 4 Correlation analysis between single chloroplast ultrastructure and plastid pigment degrading products content of flue-cured tobacco
由表5可知,未熟烟叶中,新植二烯和类胡萝卜素降解产物的含量与基粒片层数在30以上占比呈极显著正相关,与基粒片层数在21~30间的占比呈显著正相关,与基粒片层数在5以下的低基粒片层占比呈极显著负相关;适熟烟叶中,新植二烯和类胡萝卜素降解产物的含量与基粒片层数在10以下的占比呈极显著正相关,与基粒片层数在20以上的占比呈极显著负相关。

表5 烟叶叶绿体基粒片层数与质体色素降解产物含量的相关性分析Table 5 Correlation analysis of chloroplast granum lamella number and plastid pigment degrading products content of flue-cured tobacco
3 讨论
3.1 成熟度对烟叶叶绿体超微结构的影响
叶绿体的超微结构特征与植物的生长发育状况密切相关[12]。研究表明,随着烟叶成熟期的推进,叶绿体基粒片层结构逐渐解体,淀粉和嗜锇颗粒积累量增多[13],成熟期不同部位烟叶叶绿体的膜结构均离散降解[14]。本研究也表明,上、中、下部位适熟烟叶的淀粉颗粒、嗜锇颗粒数量和低基粒片层数的比例均高于未熟烟叶,随着成熟度的提升叶绿体结构破裂解体加剧。这主要由于烟叶在成熟过程中,叶片中相关酶活性降低,膜脂过氧化作用增强[15-16],进而导致叶绿体膜结构衰退[17]。
3.2 叶位对烟叶叶绿体超微结构的影响
叶绿体中基粒片层结构决定了叶绿体对光能的吸收、传递和转化能力[15]。本研究表明,烟叶叶位越高,叶绿体基粒片层结构解体程度越高;随着叶位的升高,未熟烟叶中高基粒片层数的比例较高,而适熟烟叶中低基粒片层数的比例较高。这可能是由于烟叶未熟时,细胞能量和物质代谢水平更强,叶绿体发育良好[18],光照通过影响光受体、光敏色素作用因子、生物合成相关基因的表达来促进叶绿素和类胡萝卜素的生成[19-21],使得高叶位烟叶叶绿体基粒片层的垛叠程度更高;而在适熟期,细胞逐渐衰老,叶绿体结构不完整[12-13,22],高叶位烟叶由于接受到较强的光照,使叶片中的酚-过氧化物酶系统加速了叶绿体膜结构衰退[17],导致其解体程度较低叶位烟叶更高。
本研究表明,未熟和适熟烟叶的淀粉颗粒与嗜锇颗粒积累量均随着叶位的升高而增多。这主要由于高叶位烟叶的基粒数与基粒片层垛叠数更多,光能转化效率更高,光合产物淀粉的积累量更多[5,23];且嗜锇颗粒是类囊体膜解体产生的脂质与锇酸结合产生[20],因此,高叶位烟叶中积累的淀粉颗粒与嗜锇颗粒更多。
3.3 叶位对烤烟烟叶质体色素降解产物含量的影响
质体色素降解产物占烟叶中性香味物质总量的85%~96%,在烟叶成熟和调制过程中,叶绿体中的酶催化叶绿素分解产生的叶绿醇大量脱水生成新植二烯[24],类胡萝卜素通过光敏氧化降解产生多种香味物质[25]。本研究表明,随着叶位的升高,叶绿素降解产物新植二烯和类胡萝卜素降解产物的含量均增多,且不同叶位间差异显著,与前人研究结果一致[7-8,26-27]。这可能是由于烤烟在大田生长期间,不同部位烟叶所受的光照条件不同,叶片接受的光照强度、日照时数和有效积温均存在差异,而成熟期的高温有助于叶绿体结构的降解以及质体色素降解产物的生成[28],致使烟叶质体色素降解产物的含量在不同部位间表现出显著差异。
3.4 鲜烟叶叶绿体超微结构与质体色素降解产物的关系
质体色素降解产物与叶绿体超微结构指标的相关分析结果表明,不同成熟度烟叶的单个叶绿体中淀粉颗粒数、嗜锇颗粒数与叶绿素降解产物新植二烯、类胡萝卜素降解产物含量均呈极显著正相关关系。由于淀粉颗粒与嗜锇颗粒的积累量主要受基粒片层结构的影响,叶绿体中高基粒片层数的占比越高,淀粉颗粒与嗜锇颗粒的含量越多[23,29],而质体色素主要存在于基粒片层结构上[2],因此该相关关系为间接正相关关系。
质体色素降解产物与叶绿体基粒片层结构的相关分析表明,各部位未熟烟叶的叶绿素降解产物新植二烯、类胡萝卜素降解产物的含量与基粒个数、高基粒片层比例均呈极显著正相关关系,与低基粒片层比例均呈极显著负相关;各部位适熟烟叶的叶绿素降解产物新植二烯、类胡萝卜素降解产物的含量与基粒数、高基粒片层数呈极显著负相关,与低基粒片层比例均呈极显著正相关。这主要是由于在未熟的高叶位烟叶叶绿体中,基粒数、高基粒片层数的占比较高,存在于基粒类囊体上的质体色素含量就较高[2];而高叶位烟叶叶绿体基粒片层的垛叠程度更高,在成熟的过程中,基粒片层结构发生破裂紊乱[13],能够离散出更多的低基粒片层;且随着叶绿体膜结构的衰退使得烤烟的质体色素降解产物含量增多[28]。因此,烟叶叶绿体中基粒数、基粒片层结构的垛叠程度以及基粒片层结构的解体程度共同影响了烤烟质体色素降解产物的含量。
猜你喜欢
现代农村科技(2022年8期)2022-11-18
消费电子(2020年12期)2020-01-19
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
小雪花·初中高分作文(2017年10期)2018-05-15
环球时报(2017-12-20)2017-12-20
今日财富(2017年32期)2017-10-19
科学种养(2017年3期)2017-03-23
江苏农业科学(2016年2期)2016-04-11
中学生数理化·高二版(2008年5期)2008-11-12
