
凋落物对落叶松林雪盖下土壤真菌多样性的影响
2022-04-25 05:26:12杨立宾江云兵崔福星朱道光
环境科学研究 2022年4期
杨立宾,江云兵,周 甜,崔福星,朱道光,徐 飞
1. 西华师范大学环境科学与工程学院,四川 南充 637002
2. 黑龙江省科学院自然与生态研究所,黑龙江 哈尔滨 150040
气候变暖是全球变化的主要表现之一[1],据IPCC评估报告预测,如果气温以目前的速度增长,到2052年,预计全球气温将会比工业化之前水平升高1.5 ℃[2]. 气候变暖引起降水格局的变化是全球气候变化的重要特征之一,随着气候变暖的加剧,降雪也将发生较大变化[3]. 在北半球的陆地生态系统中冬季有近50%的面积被积雪覆盖[4],有效厚度的雪被覆盖能够起到绝缘保温和保湿作用[5],同时也能够维持土壤微生物的活力[6-7],并使土壤微生物在冬季进行土壤呼吸[8]. 冬季土壤呼吸是重要的碳排放过程,研究[5]表明,冬季土壤呼吸主要来自微生物的异氧呼吸,其呼吸量占全年土壤呼吸总量的14%~30%. 目前对于冬季土壤呼吸的研究已有报道,但相关研究主要集中于极地苔原地区、高纬度地区和亚高山地区[9-14],而在其他地区相关的研究并不多见.
凋落物是土壤营养元素的主要补给者[13],能够为冬季土壤微生物存活提供有机物质与营养元素[9]. 真菌是凋落物分解过程中的重要分解者[14],有研究认为真菌是分解过程中的先锋群落[14],在分解过程的先驱阶段,真菌群落首先破坏木质素的保护层[14-16],为其他类群参与分解以及不同性质的物质分解创造条件.目前,对于冬季土壤呼吸的变化规律和产生原因尚存争议,有学者认为与碳亏缺限制作用(limitation of carbon)有关[9],而有些学者认为与微生物群落转化(shift of microbial populations between winter and summer )有关[10],还有学者认为与二者无关[11].
地处我国最高纬度的黑龙江省气温变暖的水平远超全国平均水平[17],大兴安岭地区是我国寒温带森林生态系统的典型代表,也是受气候变化影响较为敏感的区域. 研究表明,1961-2015年黑龙江省气温显著升高,大兴安岭降雪初日推迟、降雪终日提前、降雪期缩短、降雪日数减少、降雪强度增加、小雪次数和小雪量减少[17]. 目前针对寒温带森林冬季土壤呼吸和作用机制的研究并不广泛,相关假说的提出是否具有全球一致性还需进一步研究. 因此,为探讨寒温带森林冬季土壤呼吸的影响因素以及冬季土壤呼吸“碳亏缺”假说的全球一致性,该研究在大兴安岭落叶松林采用人工堆雪的方法设置有效雪被覆盖,添加不同质量凋落物,进行野外原位试验. 采用Illumina高通量测序技术,对土壤真菌进行测序,分析土壤真菌群落结构与多样性的差异,以期为冬季土壤呼吸及影响机制的研究提供参考.
1 研究方法
1.1 研究区概况
研究区域位于黑龙江省大兴安岭呼中国家级自然保护区内,试验样地(51°49′01″N~51°49′19″N、122°59′33″E~123°00′03″E)位于2014年建成的落叶松(Larix gmelinii)林25 hm2国际性监测样地内. 样地地形平缓,海拔847~974 m,年均气温-4 ℃,年均降水量458.3 mm,年均相对湿度71%,年均蒸发量911 mm[18].土壤为棕色针叶林土,土壤厚度为5~25 cm. 植被群落结构单一,是以落叶松为主要优势种的寒温带落叶针叶林,其中乔木4种,灌木37种,草本127种,隶属于21科39属. 乔木为落叶松和白桦(Betula platyphylla),主要灌木包括杜香(Ledum palustre)、笃斯越桔(Vaccinium uliginosum)等,主要草本包括舞鹤草(Maianthemum bifolium)、地榆(Sanguisorba officinalis)等[19].
1.2 试验设计与样品采集
2018年11月10日,在试验区内选取地势平坦、相对较为开阔、环境均质性较好的20 m×20 m的试验样地3块,在每块样地分别划出1 m×1 m的4个区域,分别对应添加不同质量的凋落物. 其中,自然对照组不做处理(CK),凋落物去除组(litter removal group,LR)将土壤表层凋落物层去除,凋落物加倍组(litter addition group, LA)使用0.147 mm孔径的凋落物袋在1 m×1 m的范围内均匀添加25 g低温烘干后的落叶松针叶,凋落物多倍组(litter double add group, LD)添加50 g凋落物. 采用人工堆雪的方法将雪均匀覆盖在布置好凋落物的样方上,雪被高度为50 cm.2019年2月10日将雪被和凋落物、腐殖质层移除,采集0~10 cm土壤,挑出凋落物、细根、小石块等杂物后低温研磨、混匀后置于-80 ℃下储存,其中一份用于土壤理化性质测定,另一份进行高通量测序,用于真菌多样性分析[18-19].
1.3 测定与分析
1.3.1土壤理化指标测定
采用电位法测定土壤pH;使用CN元素分析仪(Multi N/C 2100S,德国耶拿分析仪器股份公司)测定土壤OC和TN含量;采用碱解蒸馏法测定土壤AN含量;采用NaHCO3浸提比色法测定土壤AP含量;采用火焰光度计法测定土壤AK含量;采用氯仿熏蒸法测定土壤MBC含量[18-22].
1.3.2土壤DNA提取与真菌ITS基因扩增
称取0.5 g土壤样品,使用试剂盒(Carlsbad, CA,USA)法提取土壤总DNA. 以50 ng DNA为模版,采用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2R(5′-GCTGCGTTCTTCATCGATGC-3′)对土壤真菌ITS基因ITS1区进行PCR扩增. 扩增体系:5×FastPfu缓冲液4 μL,2.5 mmol/L的dNTP 2 μL,5 μmol/L的Primer F 0.8 μL,5 μmol/L的Primer R 0.8 μL,5 U/μL的Polymerase 0.4 μL,BSA 0.2 μL,DNA模板10 ng,补超纯水(ddH2O)至20 μL. PCR扩增后利用2%的琼脂糖电泳检测PCR产物,使用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences, Union City,CA, USA)凝胶回收试剂盒纯化PCR产物. 将纯化扩增产物等摩尔混合,构建Miseq文库,使用Illumina MiSeq平台对文库进行上机测序[18-22].
1.4 生物信息学分析和数据处理
使用Qiime软件进行数据去杂,以97%相似性为标准划分操作分类单元(operational taxonomic units,OTU),使用Usearch去除嵌合体. 采用RDP classifier贝叶斯算法对OTU代表序列进行分类学分析,置信度阈值为0.7. 物种分类采用Unite 8.0/itself_fungi(Unite Release 8.0)的真菌数据库进行比对和鉴定[19].采用Mothur软件进行Alpha多样性指数分析和Rarefaction分析,分别用覆盖度(Coverage)指数和艾斯(Ace)指数、香农-威纳(Shannon-Wiener)指数、辛普森(Simpson)指数分别表示测序深度和物种的多度、丰富度、均匀度[20-22]. Beta多样性基于Bray-Curtis距离计算并进行PCoA分析,用FastUniFrac分析得到样本间距离矩阵. 采用db-RDA分析和相关性Heatmap分析(采用Pearson法)进行环境因子关联分析. 采用FUNGuild软件对真菌的生态功能群进行分析. 使用R语言vegan包和ggplot包[18-22]制图.生物信息学分析采用上海美吉生物医药科技有限公司的微生物多样性云V4.0平台[19-20].
2 结果与分析
2.1 土壤理化性质
从表1可以看出:与对照组相比,各处理组土壤pH均下降,其中LD与其他各组均存在显著性差异;土壤OC含量随凋落物质量的增加呈显著升高趋势,LR与其他组相比土壤OC含量显著降低,各组之间存在显著性差异;TN和AN含量随凋落物质量的增加依次升高,其中LR、LD均与CK存在显著性差异;AP、AK含量随凋落物质量的增加而显著升高,各处理组与对照组相比均具有显著性差异;MBC含量随凋落物质量的增加依次升高,凋落物添加组与对照组无显著性差异,凋落物去除组与对照组存在显著性差异(P<0.05).
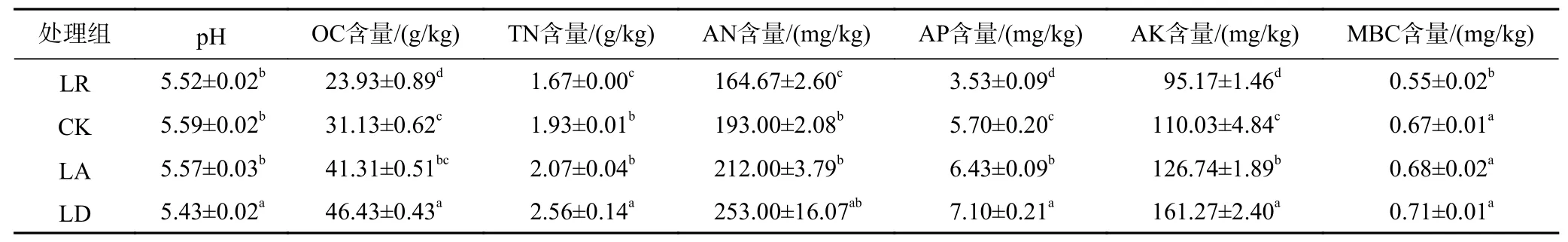
表 1 不同处理下的土壤理化性质比较Table 1 Comparison of soil physical and chemical properties under different treatments
2.2 真菌组成与群落分布特征分析
如图1所示,不同处理下土壤真菌OTUs总数排序为LA>LR>CK>LD,数量分别为797、762、733、710个. 其中,各组共有的OTUs数量为390个,特有的OTUs数量中,LR为66个,CK为52个,LA为40个,LD为31个.
在相似水平为97%的条件下,对OTUs的代表序列做分类学分析,试验测序共鉴定出11门、42纲、97目、193科、352属、534种真菌. 从图2中右侧大圆的外端半圆可以看出,在纲分类水平上按相对丰度依次为伞菌纲(Agaricomycetes)、锤舌菌纲(Leotiomycetes)、Geminibasidiomycetes、Archaeorhizomycetes、散囊均纲(Eurotiomycetes)、银耳纲(Tremellomycete)、粪壳菌纲(Sordariomycetes)、被孢霉纲(Mortierellomycetes)、茶渍纲(Lecanoromycetes)、TritirachioDothideomyc纲和麦轴梗霉纲(Tritirachiomycetes),其中伞菌纲、锤舌菌纲、Geminibasidiomycetes为各组中的优势菌纲.图2中右侧大圆的内端半圆表示某菌纲在4个组中的相对丰度排序,可以看出,各组中伞菌纲的相对丰度排序为CK>LA>LR>LD,锤舌菌纲的相对丰度排序为LD>LR>CK>LA,Geminibasidiomycetes的相对丰度排序为LD>LA>CK>LR.
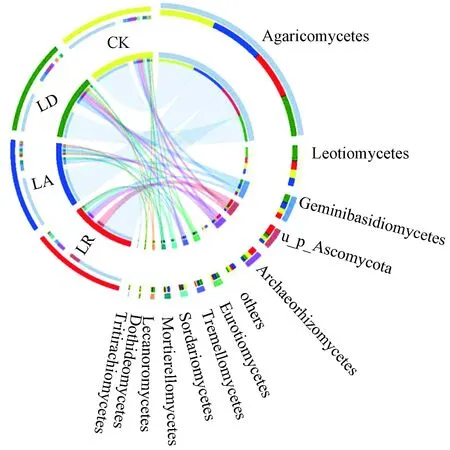
图 2 不同处理下土壤真菌纲水平上的群落组成Fig.2 Community composition of soil fungi class level under different treatments
从图2中左侧外端半圆的分析可以看出,伞菌纲在各组中的相对丰度较高,为共有优势菌纲. 随着凋落物质量的增加,伞菌纲的相对丰度先升高后降低,锤舌菌纲的相对丰度先降低后升高,Geminibasidiomycetes的相对丰度持续升高.
从图3可以看出,属分类水平上主要包括Russula、Inocybe、Geminibasidium、Archaeorhizomyces、Oidiodendron、Hygrophorus、Cortinarius、Piloderma等,其中Russula为各组中的优势菌属,各组间Russula、Inocybe、Cortinarius的相对丰度存在极显著性差异,Piloderma的相对丰度存在显著性差异.

图 3 不同处理下土壤真菌属水平上的群落组成和差异Fig.3 Community composition and difference of soil fungi at genus level under different treatments
2.3 Alpha多样性分析
对雪盖下添加不同质量凋落物的土壤真菌的Alpha多样性指数进行ANOVA分析,结果(见表2)显示,各分解等级的Coverage指数均超过99,趋近于100,且各组之间无显著性差异,说明样本中序列被测出的概率较高,表明此次测序结果能够反映样本中真菌的真实情况. 对于真菌的Ace指数,其中LR最高,LD最低,说明各组下真菌的多度有所不同,但各组之间无显著性差异. 对于Shannon-Wiener指数,其中LR最高,CK最低,说明各样品中真菌物种组成的丰富度不同,但各组无显著性差异. 对于Simpson指数,其中CK最高,LR最低,说明各组之间优势类群的优势度和整个菌群的均匀程度有所不同,但各组之间并无显著性差异.
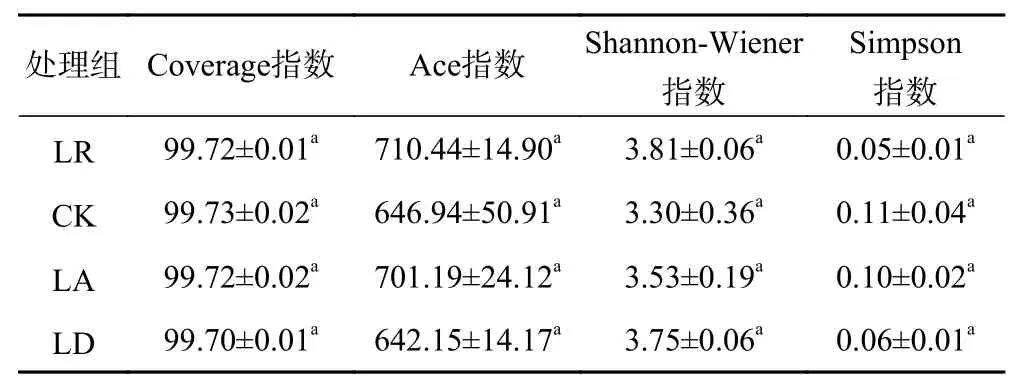
表 2 不同处理下土壤真菌多样性指数分析Table 2 Analysis of diversity index of soil fungi under different treatments
2.4 Beta多样性分析
基于Bray-Curtis距离对雪盖下添加不同质量凋落物处理的土壤真菌群落组成进行主坐标PCoA(principal co-ordinates analysis)分析. 从图4(a)可以看出:在主成分1方向上各组样品分为两大组,第一组在主成分1的负方向上(LA和LD),第二组在主成分1的正方向上(CK和LR);在主成分2方向上CK和LR被分开,分别置于第一象限和第四象限. 4个处理组的置信椭圆互有交集,其中LA和LD组椭圆面积之间的重合度较高,说明这两个处理下土壤真菌的多样性和组成更加相似. 主成分1的解释率为30.69%,主成分2的解释率为26.43%,累计解释率为57.12%.从图4(b)所示Bray-Curtis距离可以看出,LA与LD之间的距离最小,为0.611 1,说明这两组的相似性更高;而LR和LD之间的距离最大,为0.695 9,说明这两组的相似性较低.
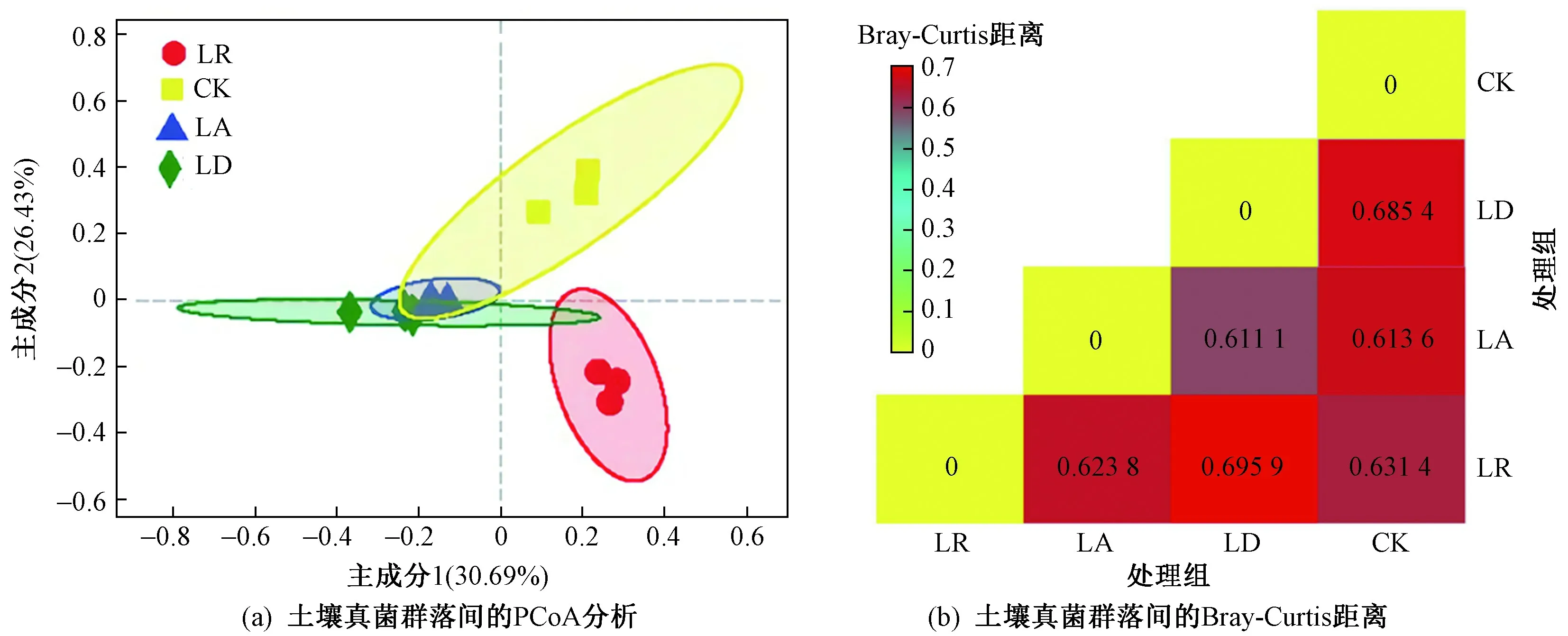
图 4 不同处理下土壤真菌群落间的PCoA分析和Bray-Curtis距离Fig.4 PCoA analysis and Bray-Curtis distance between soil fungal communities under different treatment
2.5 物种与环境因子的关联分析
基于Bray-Curtis距离,在PCoA分析的基础上加入环境因子,进行db-RDA(distance-based redundancy analysis)分析. 从图5可以看出:第一、二轴的贡献率分别为21.92%和20.17%,累计贡献率为42.09%. 与PCoA分析结果相似,全部样品被坐标轴分为3组,LA组和LD组在第一象限,LR组在第二象限,CK组在第三象限. MBC含量与土壤养分含量呈负相关,与土壤pH呈正相关.
选取属分类水平上的物种与环境因子进行相关性Heatmap分析,通过计算Pearson相关系数获得数值矩阵,以Heatmap图显示. 从图6可以看出,红菇属的相对丰度与土壤OC含量呈显著负相关,与土壤pH呈极显著正相关(P<0.01);丝盖菌属的相对丰度与TN、AN含量均呈显著负相关(P<0.05),与土壤AP含量呈极显著负相关(P<0.01);Archaeorhizomyces属的相对丰度与TN、AN、AK含量均呈显著正相关(P<0.05);丝膜菌属和子囊菌门中未定义的菌属与TN、AN、AK含量均呈极显著正相关(P<0.01).
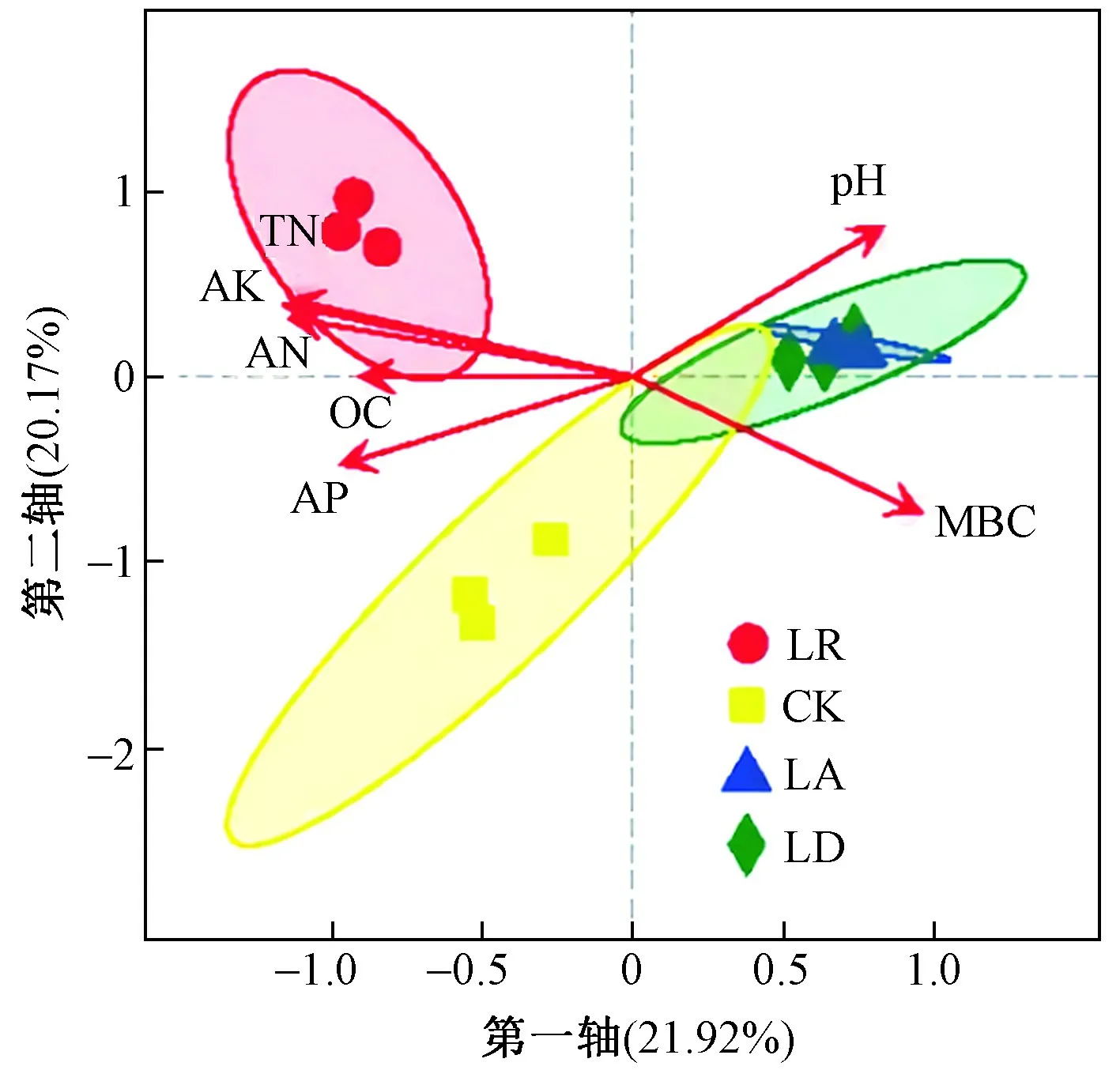
图 5 基于Bray-Curtis距离的冗余分析Fig.5 Redundancy analysis based on Bray-Curtis distance

图 6 属水平上的真菌与环境因子的相关性Heatmap分析Fig.6 Heatmap analysis of the relationship between fungi at the genus level and environmental factors
3 讨论
3.1 凋落物输入对土壤微生物生物量的影响
有效厚度的雪被(厚度大于30 cm)能够隔绝土壤与外界环境的热量交换,从而减小土壤温度的变化幅度,对土壤起到保温作用[23],而适宜的土壤温度又有利于土壤水分的保持[24-26]. 温度和水分是土壤微生物活性的重要影响因素,适宜的温度和水分改变了土壤微生物的生物量和群落组成结构[27-28]. 土壤微生物的活性影响着冬季土壤碳、氮矿化及养分利用[29],也扮演了凋落物在高寒地区森林生态系统冬季的分解主要的驱动因素;同时,凋落物的数量、成分、质地、养分均会影响土壤微生物的种类、数量及活力[11].
目前有关添加凋落物对土壤微生物生物量的影响的研究结果不尽相同,有研究发现,凋落物的添加使土壤MBC和MBN(微生物生物量氮)分别增加了13%和46%[11];但也有研究表明凋落物的影响并不明显[30],这与笔者得到的MBC含量变化结果一致. 笔者发现,有效厚度雪盖下添加不同凋落物的土壤养分发生了不同程度的变化,说明有效厚度的雪被覆盖下原有的土壤腐殖质和输入的凋落物发生了分解,同时也说明土壤微生物具有活性;这从该研究中土壤MBC含量的变化也可以看出,随着输入凋落物质量的增加,土壤微生物的生物量也升高,但与对照组相比并无显著性差异;而去除凋落物组与其他3组相比却具有显著差异. 究其原因可能是,凋落物的输入为土壤微生物提供了可利用的资源,使得土壤微生物中对于养分变化较为敏感的某些类群以及分解过程中的先锋类群生物量得到小幅提高,导致总量有所增加但并无差异性[30-31],而凋落物的去除处理则使得雪盖下具有活性的土壤微生物类群缺少可利用的资源,可能导致微生物死亡,总体生物量降低.
3.2 凋落物输入对土壤真菌Beta多样性和群落组成的影响
多样性指数是生态学和生物多样性科学中重要的群落多样性度量指标. Alpha多样性是最直接、最直观的群落比较方式,也是比较群落区别的综合指数[20];而Beta多样性是强调群落结构沿梯度的变化情况或特定梯度范围内变异的方向性[32-34]. 该研究中,添加不同质量凋落物的雪盖下土壤真菌的Alpha多样性指数虽各不相同,但并不存在显著差异,营养成分的变化并未对土壤真菌的多度、丰富度和均匀度造成显著影响. 但不同处理下土壤真菌的组成发生了变化,且在属水平上具有极显著和显著差异. 这说明虽然凋落物的改变没有影响真菌类群总体的Alpha多样性,但真菌类群的组成可能发生了转换,出现了部分耐低温的真菌类群,并能够利用凋落物维持生存和代谢. 这些嗜冷类群取代了嗜温类群,可能成为冬季低温环境中的主要真菌类群,并对土壤生态过程起到主导作用.
从表征Beta多样性的PCoA分析以及在此基础上加入环境因子的db-RDA结果可以看出,由于凋落物的添加,各组之间计算得到的Bray-Curtis距离有不同程度的差异,LR和LD与其他两组差异较大,说明凋落物的改变造成土壤养分的不同,导致土壤真菌对土壤养分的利用也存在差别. 这从土壤真菌的群落组成和相对丰度的变化上也能够得到印证,特别是优势菌属中的红菇属以及相对丰度较大的丝盖菌属在各组之间都具有极显著性差异,而且这两个属的菌群与碳源、氮源均呈显著相关. 土壤真菌生态功能组随凋落物输入的变化也出现了相似的结果,相对丰度较大的外生菌根真菌和腐生真菌群变化并不相同,这可能是由于各菌群与碳、氮元素的偏好和碳氮元素的限制性有关. “碳亏缺”假说认为,在冬季,土壤微生物对碳的消耗会造成碳供应亏缺,导致土壤真菌逐渐死亡[9]. 综上,土壤真菌对于外源营养物质特别是碳元素的敏感性更高,证明碳元素是土壤真菌生长的限制因子. 该研究还发现,氮元素与一些生态功能菌群的相关性很高,说明氮元素可能是冬季土壤真菌活性的又一个限制因子,而能否成为“碳亏缺”理论的补充仍然需要进一步研究.
全球气候变化背景下,大兴安岭寒温带兴安落叶松林冬季雪盖下凋落物质量的变化改变了土壤养分,但并未显著改变土壤真菌类群总体的Alpha多样性,而去除凋落物显著降低了土壤真菌生物量;Beta多样性和db-RDA分析显示,土壤真菌在属水平上存在显著性差异,不同菌属的真菌类群与各项土壤化学特征具有显著相关性.
4 结论
a) 在大兴安岭寒温带兴安落叶松林设置有效雪被覆盖,不同质量凋落物的添加改变了土壤的理化性质,其中土壤养分含量随凋落物质量的增加而上升.其中,LD的土壤pH显著低于其他3组,而LR的MBC含量显著高于其他3组(P<0.05);此外,CK、LA、LD的TN、AN、AP和AK含量均显著高于LR(P<0.05),表明土壤养分含量会随凋落物质量的增加而呈上升的趋势.
b) 就土壤真菌而言,其多度、丰富度、均匀度等多样性指数和微生物生物量并未发生显著改变.
c) “碳亏缺”假说认为碳是冬季土壤微生物活性的限制因子,该研究发现丝膜菌属和子囊菌门中未定义的菌属均与TN、AN含量呈极显著正相关(P<0.01),而丝盖菌属相对丰度与TN、AN含量呈负显著相关(P<0.05). 由此可见,氮元素可能是寒温带落叶针叶林冬季土壤真菌活性的又一个限制因子.
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
人民黄河(2020年12期)2020-12-30 12:55:30
山西农业科学(2020年9期)2020-09-14 08:20:08
山西农业科学(2019年12期)2019-12-19 04:03:30
江苏农业科学(2019年5期)2019-09-02 14:01:46
广东农业科学(2017年5期)2017-08-29 10:37:31
电子制作(2017年10期)2017-04-18 07:23:14
水生生物学报(2015年1期)2015-02-28 16:01:05
应用海洋学学报(2014年1期)2014-11-22 07:17:44
