
草海湖滨带沉积物微生物群落对磷形态的影响
2022-04-24 11:09龙云川张珍明朱成斌苏荣翔周少奇
中国环境科学 2022年4期
龙云川,蒋 娟,胡 菁,张珍明,朱成斌,苏荣翔,周少奇*
草海湖滨带沉积物微生物群落对磷形态的影响
龙云川1,2,蒋 娟1,2,胡 菁2,张珍明1,2,朱成斌2,苏荣翔1,周少奇2*
(1.贵州科学院贵州省生物研究所,贵州 贵阳 550009;2.贵州大学资源与环境工程学院,贵州 贵阳 550025)
为探究高原湖泊湖滨带沉积物中微生物群落对磷赋存形态的影响,以草海湖滨带沉积物为研究对象,分析了磷素赋存形态;通过高通量测序技术分析了细菌和古菌的群落组成.结果表明,草海湖滨带表层沉积物的总磷含量范围在662.89~881.26mg/kg之间,平均值为750.36mg/kg;各磷形态中NaOH-NRP>Res-P>BD-P>HCl-P>NaOH-SRP>NH4Cl-P.细菌群落由变形菌门(Proteobacteria)等59个门和硫杆菌属()、厌氧粘细菌属()等1259个属组成;古菌群落由泉古菌门(Crenarchaeota)等8个门和甲烷丝菌属()等67个属组成.冗余分析、主成分分析和共现性网络分析表明,微生物群落能驱动草海沉积物中磷的形态转化,细菌主要通过影响铁的氧化/还原和碱性磷酸酶的活性;古菌则通过改变有机质和酸性磷酸酶活性来调控沉积物磷形态.
沉积物;高通量测序;磷形态;共现性网络;古菌
沉积物是湖泊系统中污染物的主要蓄积场所,沉积物内源磷是湖泊中水体富营养化的关键因子[1].沉积物中不同磷形态的迁移度和生物利用度存在差异[2].已有研究表明微生物可以通过分泌有机酸、磷酸酶等胞外物质,在土壤和沉积物中磷的循环中起关键作用[3-4].沉积物中丰富的微生物在湖泊生态系统的能量流动、元素迁移转化、污染物降解和水体自净等过程中发挥着重要作用[5].古细菌作为除细菌和真核生物外的主要域,虽介导了甲烷生成、有机物分解、好氧氨氧化、硫酸盐还原和磷酸盐增溶等生物地球化学过程[6],但对古菌的研究较少,需更多的关注.
湖滨带作为水体生态系统与陆地生态系统的水陆交错带[7],其沉积物中微生物群落的组成结构和功能受到周边人类活动的干扰.由于高原湖泊生态系统的特殊性,其沉积物中的微生物群落也与其他生态系统中有着显著不同[8].贵州草海,典型的高原湖泊,临近威宁县城且周边村庄耕地众多,其人类活动带来的湖滨带微生物群落组成变化及其对元素生物地球化学循环的影响也鲜为人知.因此,本研究以贵州草海的湖滨带沉积物为研究对象,分析沉积物细菌和古菌的群落组成、多样性和磷素赋存形态,探究沉积物中微生物群落对磷形态的影响.
1 材料与方法
1.1 研究区域概况
贵州草海位于云贵高原中部,是国家级自然保护区、典型的高原湿地生态系统.草海湖区面积25km2,平均海拔2171.7m,年平均气温10.5℃,年平均日照时长1805.4h,年平均降雨量950mm[9].草海是黑颈鹤等鸟类的越冬栖息地,有“高原明珠”美称[10].威宁县城位于草海东北部,距离草海湖区仅数公里.此外,依托西海码头(位于草海北侧)、江家湾码头(位于草海北侧)、白家嘴码头(位于草海东侧)的旅游活动对湖泊生态系统造成了强烈干扰.
1.2 样品采集与处理
2019年6月,根据草海湿地湖滨带的地形设置如图1所示的12个采样点(编号分别为S21~S32).使用抓斗采样器采集0~10cm表层沉积物样品,装入无菌自封袋内密封低温保存带回实验室.沉积物样品经冷冻干燥后去除石块、动植物残体等杂质,研磨,过100目筛后置于-20℃冰箱内待测.
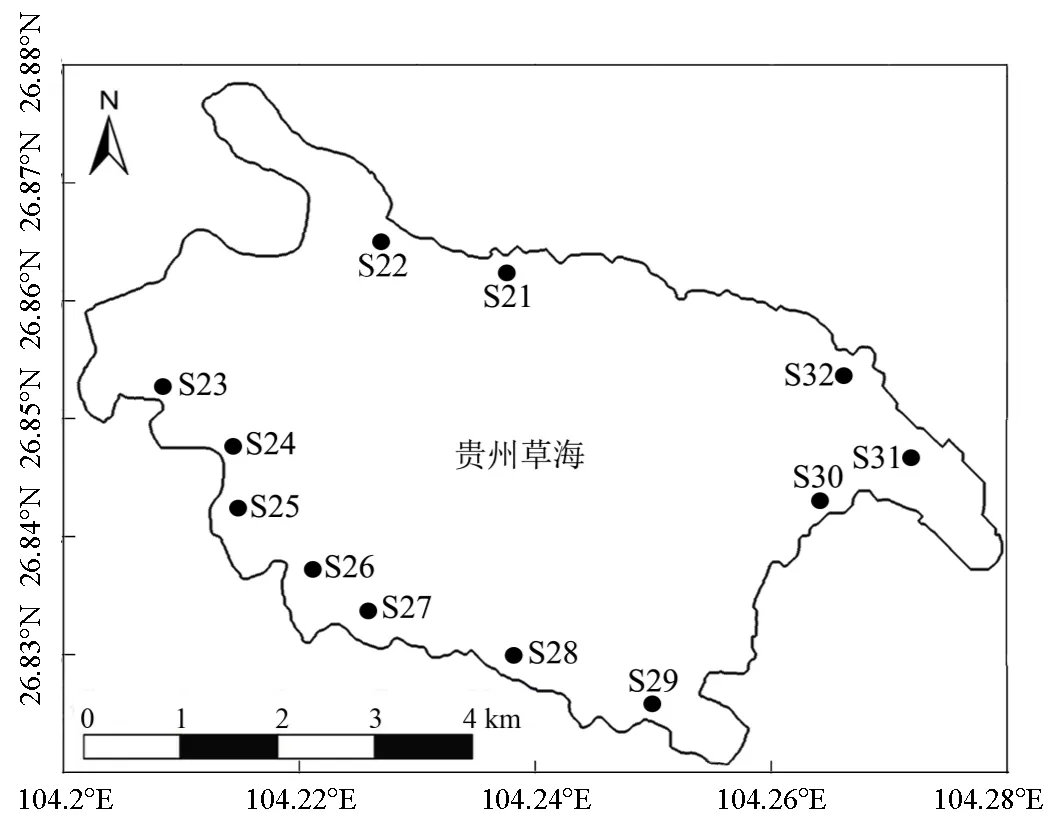
图1 贵州草海湖滨带沉积物采样点分布
1.3 沉积物磷形态及性质测定
沉积物中磷形态的测定采用Psenner化学顺序提取方法[11],该方法将磷分为弱吸附态磷(NH4Cl- P)、可还原态磷(BD-P)、铁铝氧化态磷(NaOH- SRP)、有机质结合态磷(NaOH-NRP)、钙结合态磷(HCl-P)和残渣态磷(Res-P) 6种磷形态.沉积物烧失量(LOI)采用灼烧法测定,沉积物酶活性检测采用索莱宝土壤酶活性检测试剂盒利用分光光度法测定.
1.4 沉积物微生物多样性分析
使用Power Soil®DNA提取试剂盒(Mo Bio Laboratories)提取0.5g沉积物中的细菌和古菌DNA.使用通用引物338F/806R扩增细菌16S rRNA的V3~V4高变区;使用引物524F/958R扩增古菌的16S rRNA.PCR扩增产物纯化、建库后在Illumina MiSeq PE300平台上进行高通量测序(上海美吉生物公司执行).测序原始序列经拼接、筛选、聚类后,获得的有效序列进行生物信息学分析.
1.5 数据分析
利用Excel 2013和SPSS 23.0软件进行数据处理和分析,利用Origin 2018作图.采用Pearson相关系数分析理化指标与磷形态的相关性,采用主成分分析(PCA)和冗余分析(RDA)探讨微生物群落对磷赋存形态的影响,采用Gephi 0.9.2软件分析沉积物微生物群落与磷形态的共现性网络.
2 结果与分析
2.1 草海沉积物中细菌群落组成

表1 微生物群落的alpha多样性指数
对于细菌微生物群落,12个样品的有效序列共452683条,平均长度为417.77bp.根据相似水平取值97%,共聚类8786OTUs.本文采用Chao1指数反映草海沉积物微生物群落的丰富度,Shannon评价微生物群落的多样性.从表1可以看出,草海湖滨带沉积物细菌群落的Chao1指数为(4347±654.89),高于白洋淀湖泊的沉积物(Chao1指数(1354.96±645.82))[12]、华南地区的红树林沉积物(Chao1指数(2243±96))[13],这说明草海湖滨带沉积物中细菌群落的丰富度较高.Shannon值与群落多样性呈正比[14],Shannon指数的平均值为(6.58±0.41),说明草海湖滨带沉积物中的细菌群落较为多样.
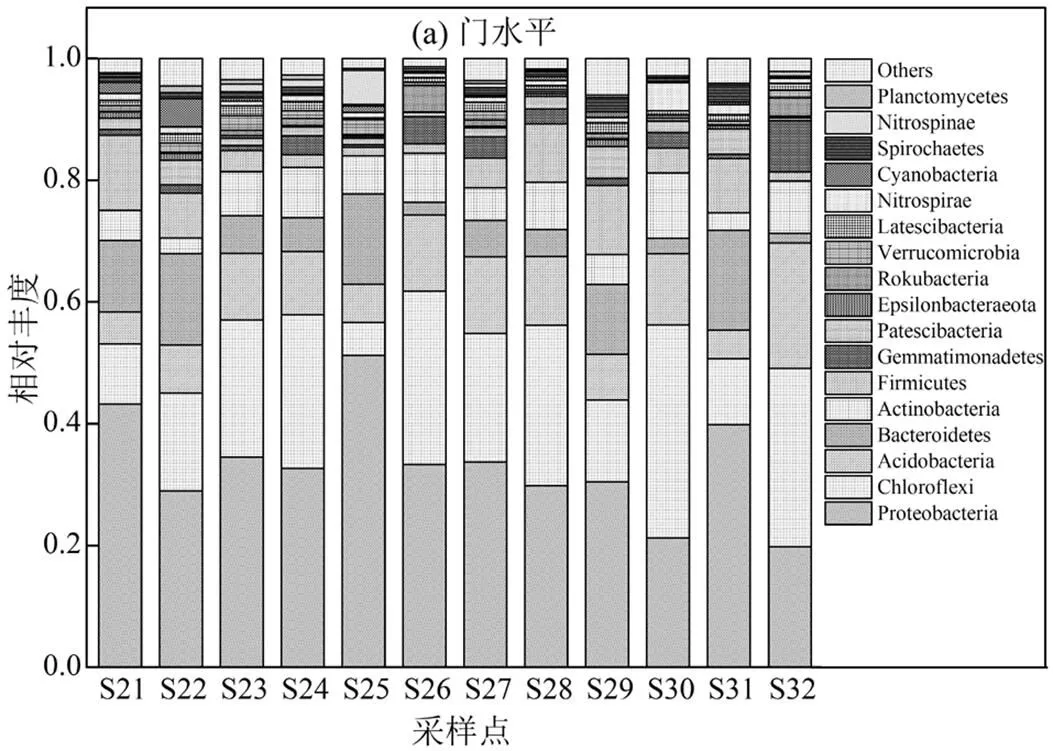
根据分类学分析,12个样本的沉积物细菌群落共包含59门、141纲、375目、654科、1259属、2762种.在门水平上,变形菌门(Proteobacteria)占主导地位(19.80%~51.26%).其次是,绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)、芽单胞菌门(Gemmatimonadetes)、Patescibacteria门、埃普西隆杆菌门(Epsilonbacteraeota)、棒状杆菌门(Rokubacteria)、疣微菌门(Verrucomicrobia)、Latescibacteria门、硝化螺旋菌门(Nitrospirae)、蓝细菌门(Cyanobacteria)、螺旋体门(Spirochaetes)、硝棘菌门(Nitrospinae)、浮霉菌门(Planctomycetes)16门,其相对丰度均大于1%(图2a).在属水平上,硫杆菌属()和厌氧粘细菌属()的丰度最高(图2b),其次是梭菌属()、硫氧化菌属()、铁氧化属()、厌氧绳菌属()、地发菌属()、地杆菌属()、芽单胞菌属()、苔藓杆菌属()等.
2.2 草海沉积物中古菌群落组成
对于古菌群落,12个样品的有效序列共470522条,平均长度为427.94bp.根据97%相似性,聚类至979OTUs.从表1可以看出,古菌群落的Chao1指数为(333.42±59.76),说明草海湖滨带沉积物古菌群落的丰富度低.Shannon指数为(2.90±0.63),说明草海湖滨带沉积物细菌群落多样性不高.比较细菌和古菌群落,两者有效序列数无显著差异,且古菌群落的有效序列数还稍高;聚类得到的OTU数,细菌群落远高于(8.97倍)古菌群落;分类得到的门、属的数量,细菌群落也远高于古菌群落.细菌群落的丰富度远高于(大于11倍)古菌群落,但细菌群落的多样性稍高于(2.27倍)古菌群落.
系统发育分类结果显示,12个样本的古菌群落共包含8个古菌门,包括25纲37目50科67属152种.在门分类水平上,相对丰度大于1%的古菌门有5个,如图3a所示.丰度最高的门为泉古菌门(Crenarchaeota),其相对丰度分布范围为4.46%~71.17%,平均值为44.49%;其次为广古菌门(Euryarchaeota)、奇古菌门(Thaumarchaeota)、Asgardaeota门、Diapherotrites门.未分类(unclassified)古菌门和相对丰度低于0.1%的稀有古菌门归类于“Others”,其占比为1.04%~8.99%,平均值为2.68%.图3b列出了草海沉积物中古菌群落的主要属.甲烷丝菌属()相对丰度最高,其次为、甲烷八叠球菌属()、甲烷杆菌属()、等.值得注意的是,未确定分类地位的属(unclassified、norank)和稀有属(相对丰度在所有样本中均低于0.1%)占每个样本相对丰度的50%以上.此外,在所有样品中无法确定分类地位的属的平均相对丰度约为80%,这表明草海沉积物中存在大量未知作用的古菌.总体来看,尽管各样点沉积物均包含有大部分微生物类群,但各采样点的微生物群落组成存在一定差异.

2.3 草海湖滨带沉积物磷素赋存特征
湖泊沉积物是水生态系统中磷的重要蓄积库,生活污水、农业排放的磷及湖泊生物组成磷都可能进入到沉积物中,使沉积物中的磷素富集增多.草海湖滨带表层沉积物的总磷含量分析结果如图4所示,草海湖滨带沉积物不同点位受不同人类活动影响其总磷有所差异,其中S29、S31、S32采样点的总磷含量最高.S29采样点位于草海东南部,周边村庄耕地分布多;S31、S32采样点位于草海东北部,临近威宁县城.据统计,中国耕地磷肥年平均使用量高达100kg/hm2,分别是美国和欧盟同期磷肥使用量的2.6和2.5倍.在这种长时间、高强度施肥之后,只有10%~15%的磷能被吸收,而其余的则流失到径流和/或地下水中[15].另外,畜禽粪便产量达60亿t(2015年统计数据),但60%以上的畜禽粪便未经处理而随意排放.因此,S29样点总磷含量高可能与周边村庄的含磷生活污水、畜禽粪便及农业面源污染的无序排放有关.此外,2017年之前,威宁县城每天连续向草海直接排放约8000t污水[8].S31、S32采样点临近威宁县城,其总磷含量高与历史上长期排放未经处理的污水有关.总磷含量范围在662.89~881.26mg/kg之间,平均值为750.36mg/kg.贵州草海总磷含量远高于长江中下游的洞庭湖、鄱阳湖、洪泽湖等湖泊(600mg/kg以下)[16],这与贵州草海为半封闭性城市高原湖泊有关.草海海拔较高、换水周期长、易于沉积物磷的累积,并长期受县城污水排放及周边农业面源污染径流输入的影响,因此需加强草海流域生活污水及农业径流的处理与控制.

图4 沉积物各形态磷含量分布
如图4所示,NaOH-NRP是各形态中占比最高的磷组分,其含量为262.37~354.78mg/kg,平均值为314.25mg/kg.通常NaOH-NRP为有机物和微生物生物质结合的磷[11],相对化学惰性,但在一定条件下可转化为小分子正磷酸盐.草海湖滨带沉积物中占比最小的是NH4Cl-P,含量为1.42~5.10mg/kg.NH4Cl-P是一种松散吸附态磷,是最不稳定、生物有效性最高的磷组分,容易被水生植物利用并释放到上覆水体中[17].样点S29,S31, S32的总磷和NH4Cl-P含量都较高,磷素释放潜力较大.BD-P对氧化还原条件非常敏感,主要是与铁氢氧化物和锰化合物结合的磷,具有潜在的迁移性[18].草海湖滨带沉积物BD-P含量为60.07~113.64mg/kg. NaOH-SRP通常被认为是与铁铝结合的磷组分,并与氧化还原不敏感的氢氧化铝强耦合[11].NH4Cl-P、BD-P、NaOH-SRP和NaOH-NRP被认为是活性磷[17].活性磷较为活跃,受沉积物-水界面的物理、化学及生物条件变化影响较大,在生物地球化学作用下可能发生一系列的迁移转化,影响湖泊生态系统中磷的分布.草海湖滨带沉积物中的活性磷总含量为430.99~516.58mg/kg,平均值为472.05mg/kg,占总磷的63.21%.相对较高的活性磷占比预示着大部分磷可释放至上覆水体,草海湖滨带沉积物中能释放至水体的潜在内源磷负荷不容忽视.
Res-P即残渣态磷,主要是大分子有机磷或其他难溶性磷,被认为是永久结合态磷[19], 其平均含量为204.75mg/kg. HCl-P即钙磷,是与沉积碳酸钙或自生的磷灰石结合的磷.它稳定存在于各种岩土矿物中,生物难以直接利用,在弱碱性水环境中比较稳定[20].草海湿地的水质pH值为弱碱性,因而HCl-P释放较难.此外,根据相关理论,当沉积物中Ca/(Fe+ Al)比值大于0.7时,为钙质沉积物[19];草海沉积物的Ca/(Fe+Al)平均值约为0.3,该区域沉积物为非钙质沉积物,因而HCl-P在总磷中占比较低(平均值9.81%).总之,草海湖滨带沉积物中各形态磷的含量分布为NaOH-NRP>Res-P>BD-P>HCl-P>NaOH- SRP>NH4Cl-P.
2.4 沉积物性质
草海湖滨带沉积物性质的结果见表2.烧失量(LOI)可以反映沉积物中有机质的含量.草海湖滨带沉积物的LOI变化范围为8.47%~29.33%.与云南滇池(10.2%~18.5%)、太湖(2.0%~7.6%)[21]相比,草海有机质含量相对较高,且存在明显的空间差异.草海湖滨带沉积物中有机质含量差异较大,总体上草海东部区域(S29~S32采样点)沉积物的LOI明显高于其他区域.草海东部区域更靠近威宁县城,县城大量的生活污水排放(8000t/d)增加了湖滨带沉积物有机质负载[8];另外,该区域水深较浅,有利于水生植物的生长繁殖,分布广泛的水生植物产生的植物残体、腐殖质等物质使得该区域沉积物的LOI含量高.沉积物中的酶多产自环境中的微生物,草海湖滨带沉积物的微生物群落组成差异会引起酶活性大小的变化.从结果来看,碱性磷酸酶(AKP)活性的变化范围为1.49~6.30μmol/(d·g),平均酶活性是(3.49±1.40)μmol/ (d·g);酸性磷酸酶(ACP)活性为5.87~12.03μmol/ (d·g),平均值为(8.07±2.08)μmol/(d·g).这2种酶活性的最大值均出现在S32采样点.该样点紧邻威宁县城,总磷含量也高,且水生植物生长茂密[22],外源营养物质的流入加强了该区域水生植物及植物根际微生物的活动,从而使沉积物里的酶活性增高.

表2 草海沉积物性质
3 讨论
沉积物中磷的迁移转化是一个复杂的过程,涉及物理、化学及生物等诸多因素.其中微生物是沉积物磷转化的关键驱动因子,但对其相关研究较少,微生物对磷的影响机制不清.贵州草海湖滨带沉积物中微生物群落组成较为丰富,共有细菌1259属, Chao1指数高达(4347±654.89).为深入了解沉积物微生物群落对磷形态的影响机制,本文采用了冗余分析(RDA)、主成分分析(PCA)和共现性网络分析3种手段.
微生物群落(丰度最高的10个细菌属和古菌属)对不同形态磷的冗余分析(RDA)分析结果如图5所示.细菌群落的10个主要属共解释了95.1%的磷形态,其中、、、和分别解释了23.6%、27.4%、12.8%和8.4%.是分布广泛、研究最多的一类铁还原菌[23],它的主要作用是铁还原、硫还原及甲烷生成[24].有利用有机物作为电子供体进行铁异化还原的功能.铁是地壳中丰度排第4的元素,在沉积物中分布广泛;由于沉积物的潮湿缺氧的环境,铁多以二价态的形式相对稳定的存在于沉积物中;但同时存在的三价态铁通常以铁氧复合物的形式发生沉淀,并对沉积物中的磷具有强烈的吸附和固定作用.RDA图中对NaOH-SRP和BD-P有显著影响(图5a),而BD-P主要是与铁锰结合的磷,NaOH-SRP是与铁铝结合的磷组分.此外,在NaOH-SRP含量分布最高的区域(S25),也表现出最高的相对丰度(图4).有研究表明,能将单质硫或还原性硫化物氧化为硫酸盐[25].硫化物可以和磷竞争与铁(III)氧化物结合,导致铁(III)结合态中磷的活化[24].综合来看,细菌主要通过铁(III)氧复合物来调节沉积物磷形态.

古菌群落丰度最高的10个属中,、、、分别解释了36.9%、12.0%、11.8%、7.5%的磷形态变异.可以利用硝酸盐或Fe3+或Mn4+作为电子受体,进行甲烷的厌氧氧化[26].多参与胁迫环境下的氮代谢.和都是产甲烷菌,在沉积物厌氧环境下进行有机物降解等碳代谢过程[27].这些古菌基本上都有甲烷代谢等碳代谢功能;此外,细菌Thiobacillus也是土壤中主要的解磷菌,能通过产磷酸酶等方式将难以被利用的磷转化为生物可利用的磷[28].
因此,运用主成分分析(PCA)研究磷酸酶、LOI(反映碳含量)及微生物群落(选取RDA分析中对磷形态解释度最高的4种细菌属和4种古菌属)对沉积物磷形态的影响,结果如图6所示.PC1能解释36.71%的总方差,PC1对总磷、NH4Cl-P、Res-P、HCl-P、NaOH-NRP及ACP、LOI有较高的正载荷,说明有机质(以LOI指示)和磷酸酶(以ACP指示)对沉积物中磷形态有较大影响.一般情况下,有机质是有机磷的重要载体,高浓度的有机质通常伴随着较高的有机磷含量[29],因此有机质与磷组分往往表现出较强的相互作用关系.磷酸酶能水解有机磷的磷酸酯或磷酸酐,将有机磷转化至无机磷,从而让磷被水生生物利用[30].、、、、等微生物与大部分磷形态处于同一区间(-0.294~0.213),这也间接验证了如前所述‘细菌群落和古菌群落对磷形态产生重要影响’的观点.
为了形象的展示沉积物微生物群落(丰度最高的10个细菌属和古菌属)、磷形态及沉积物性质的相互关系,运用Gephi生成相关性网络图(相关系数绝对值大于0.4),如图7所示.细菌群落alpha指数Shannon与NaOH-SRP负相关,相关系数为-0.652.细菌丰度最高的10个属中,、、与多个磷形态相互作用关系紧密.其中,与BD-P、HCl-P、NaOH- NRP及总磷相关性较强;与NaOH-SRP、NaOH-NRP、总磷的相关系数分别为-0.601、-0.704和-0.533;与NaOH-SRP和HCl-P的相关系数分别为0.591和-0.585.是一类典型的铁氧化细菌,能将沉积物中广泛存在、相对稳定的二价态铁通过生物铁氧化反应氧化为三价态铁,并以铁氧复合物的形式吸附沉积物中的磷,导致沉积物中磷的固定[31].结合前面分析的细菌属的功能,发现细菌主要通过铁的氧化/还原来影响沉积物磷形态.相关性网络中的古菌群落中, alpha指数Shannon、Chao1与BD-P、NaOH-SRP、NaOH-NRP等磷形态相关性较强,说明古菌群落的丰富程度和多样性程度强烈影响着沉积物中的磷形态.具体地,丰度最高的10个古菌属中,与多个磷形态相互作用关系较强.
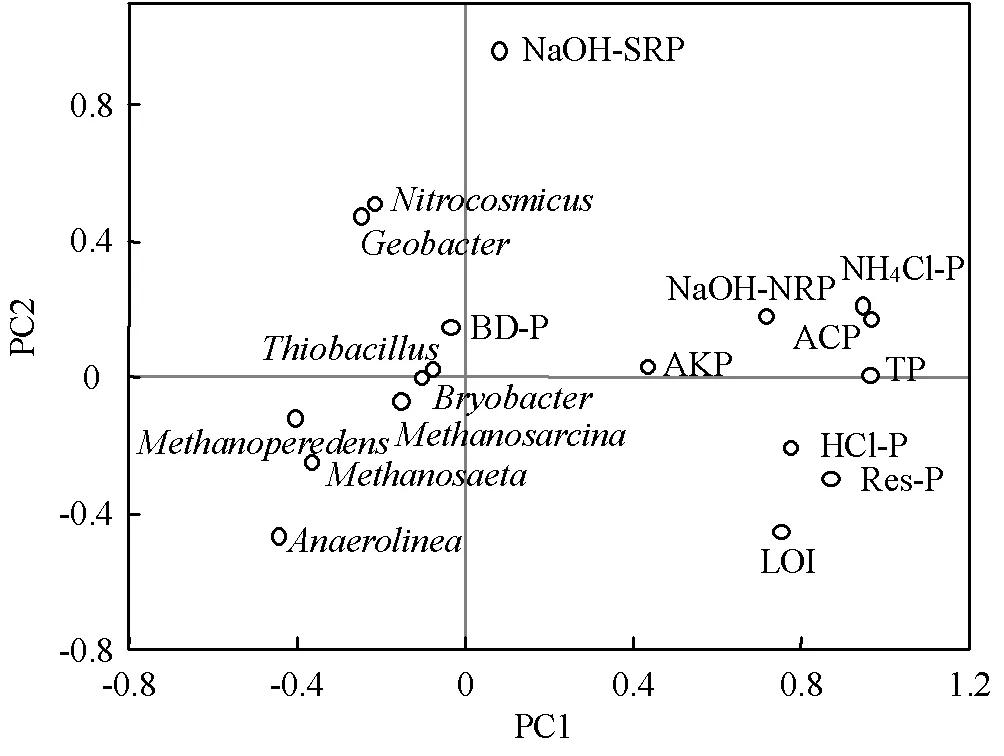
图6 沉积物微生物、性质与磷形态的主成分分析

图7 沉积物微生物、磷形态及沉积物性质的共现性网络
每个节的颜色表示不同的分类.红线和绿线分别表示正相关和负相关;线条粗细表示相关性的大小
此外,分析有机质(LOI指示)和磷酸酶(ACP、AKP指示)在微生物对磷形态影响中的作用.从图7中可以看出,AKP、ACP酶、LOI与多个磷组分相关性强,这也证实了PCA结果中关于有机质和磷酸酶对磷形态影响较大的论述.细菌、、、与AKP酶相关性较强;这说明草海沉积物中细菌群落可以分泌碱性磷酸酶来影响沉积物中磷形态的分布.古菌、、与ACP酶相关性较强;古菌、与LOI相关性较强;这表明草海沉积物中的古菌通过影响酸性磷酸酶和有机质来影响沉积物中的磷形态.
总体而言,草海湖滨带沉积物中微生物群落能驱动磷赋存形态的变化,细菌群落主要通过铁的氧化/还原和调控碱性磷酸酶活性来影响沉积物磷形态,而古菌群落则通过改变酸性磷酸酶活性和有机质来影响沉积物磷形态.下一步,建议加强对包括生物和非生物因子的高分辨率原位分布,磷与碳、氮等关键元素循环耦合的化学计量机制,以及元素循环中关键菌群或物种的作用等关键过程的研究,以进一步探明沉积物磷素生物地球化学循环机制.
4 结论
4.1 草海湖滨带沉积物的总磷含量为662.89~881.26mg/kg,其中NaOH-NRP>Res-P>BD-P>HCl- P>NaOH-SRP>NH4Cl-P.
4.2 草海湖滨带沉积物中微生物群落组成存在一定的空间差异,细菌群落丰富度和多样性高于古菌.细菌群落共含有1259个属,以硫杆菌属、厌氧粘细菌属为优势属;古菌群落以甲烷丝菌属为主.
4.3 草海湖滨带微生物群落能驱动沉积物磷形态的转化,细菌主要通过调控铁的氧化/还原和碱性磷酸酶活性来影响磷形态,而古菌则通过改变有机质和酸性磷酸酶活性来调控磷形态.
[1] Kurek M R, Harir M, Shukle J T, et al. Seasonal transformations of dissolved organic matter and organic phosphorus in a polymictic basin: Implications for redox-driven eutrophication [J]. Chemical Geology, 2021,573:120212.
[2] Crocker R, Blake W H, Hutchinson T H, et al. Spatial distribution of sediment phosphorus in a Ramsar wetland [J]. Science of the Total Environment, 2021,765:142749.
[3] Yao Q, Li Z, Song Y, et al. Community proteogenomics reveals the systemic impact of phosphorus availability on microbial functions in tropical soil [J]. Nature Ecology & Evolution, 2018,2(3):499-509.
[4] Mohapatra M, Yadav R, Rajput V, et al. Metagenomic analysis reveals genetic insights on biogeochemical cycling, xenobiotic degradation, and stress resistance in mudflat microbiome [J]. Journal of Environmental Management, 2021,292:112738.
[5] 周佳敏,黄廷林,刘 茜,等.低C/N亚铁氧化硝酸盐还原菌群脱氮特性 [J]. 中国环境科学, 2021,41(8):3723-3732.
Zhou J M, Huang T L, Liu Q, et al. Denitrifying characteristics of low C/N ferrous oxidation nitrate reducing bacteria flor [J]. China Environmental Science, 2021,41(8):3723-3732.
[6] Wei G, Li M, Shi W, et al. Similar drivers but different effects lead to distinct ecological patterns of soil bacterial and archaeal communities [J]. Soil Biology and Biochemistry, 2020,144:107759.
[7] Wang W, Liu W, Wu D, et al. Differentiation of nitrogen and microbial community in the littoral and limnetic sediments of a large shallow eutrophic lake (Chaohu Lake, China) [J]. Journal of Soils and Sediments, 2018,19(2):1005-1016.
[8] Long Y, Jiang J, Hu X, et al. The response of microbial community structure and sediment properties to anthropogenic activities in Caohai wetland sediments [J]. Ecotoxicology and Environmental Safety, 2021, 211:111936.
[9] Xia P, Ma L, Sun R, et al. Evaluation of potential ecological risk, possible sources and controlling factors of heavy metals in surface sediment of Caohai Wetland, China [J]. Science of the Total Environment, 2020,740:140231.
[10] 朱成斌,胡 菁,龙云川,等.贵州草海沉积物重金属元素分布特征及健康风险评价[J]. 环境科学学报, 2021,41(6):2212-2221.
Zhu C B, Hu J, Long Y C, et al. Distribution fractionation and health risk assessment of heavy metals in surface sediments from Caohai Lake, Guizhou [J]. Acta Scientiae Circumstans, 2021,41(6):2212-2221.
[11] Markovic S, Liang A, Watson S B, et al. Biogeochemical mechanisms controlling phosphorus diagenesis and internal loading in a remediated hard water eutrophic embayment [J]. Chemical Geology, 2019,514: 122-137.
[12] Wang W, Yi Y, Yang Y, et al. Impact of anthropogenic activities on the sediment microbial communities of Baiyangdian shallow lake [J]. International Journal of Sediment Research, 2020,35(2):180-192.
[13] Tong T, Li R, Wu S, et al. The distribution of sediment bacterial community in mangroves across China was governed by geographic location and eutrophication [J]. Marine Pollution Bulletin, 2019, 140:198-203.
[14] Long Y, Jiang J, Hu X, et al. Actinobacterial community in Shuanghe Cave using culture-dependent and -independent approaches [J]. World Journal of Microbiology and Biotechnology, 2019,35(10):1-12.
[15] Ni Z, Wang S, Wu Y, et al. Response of phosphorus fractionation in lake sediments to anthropogenic activities in China [J]. Science of the Total Environment, 2020,699:134242.
[16] 黎 睿,王圣瑞,肖尚斌,等.长江中下游与云南高原湖泊沉积物磷形态及内源磷负荷[J]. 中国环境科学, 2015,35(6):1831-1839.
Li R, Wang S R, Xiao S B, et al. Sediments phosphorus forms and loading in the lakes of the mid-lower reaches of the Yangtze River and Yunnan Plateau, China [J]. China Environmental Science, 2015,35(6): 1831-1839.
[17] Yang P, Yang C, Yin H. Dynamics of phosphorus composition in suspended particulate matter from a turbid eutrophic shallow lake (Lake Chaohu, China): Implications for phosphorus cycling and management [J]. Science of the Total Environment, 2020,741:140203.
[18] Wu Z, Wang S, Ji N, Phosphorus (P) release risk in lake sediment evaluated by DIFS model and sediment properties: A new sediment P release risk index (SPRRI) [J]. Environmental Pollution, 2019,255 (Pt2):113279.
[19] 刘 超,朱淮武,王立英,等.滇池与红枫湖沉积物中磷的地球化学特征比较研究[J]. 环境科学学报, 2013,33(4):1073-1079.
Liu C, Zhu H W, Wang L Y, et al. Comparative study on the geochemical characteristics of phosphorus in sediments from Lake Dianchi and Hongfeng [J]. Acta Scientiae Circumstantiae, 2013,33(4): 1073-1079.
[20] 张紫霞,刘 鹏,王 妍,等.典型岩溶湿地表层沉积物中磷的形态及分布特征研究[J]. 西南林业大学学报(自然科学), 2021,41(3): 78-86.
Zhang Z X, Liu P, Wang Y, et al. Occurrence form and distribution characteristics of phosphorus in surface sediments of typical karst wetlands [J]. Journal of Southwest Forestry University, 2021,41(3): 78-86.
[21] Xie F, Li L, Song K, et al. Characterization of phosphorus forms in a Eutrophic Lake, China [J]. Science of the Total Environment, 2019, 659:1437-1447.
[22] 吴胜利,夏品华,林 陶,等.贵州草海不同水位梯度下沉积物氮赋存形态及分布特征[J]. 湖泊科学, 2020,31(2):407-415.
Wu S, Xia P, Lin T, et al. Contents and distribution characteristics of nitrogen forms in sediments of Guizhou Lake Caohai under different water level levels [J]. Journal of Lake Sciences, 2019,31(2):407-415.
[23] 万 杰,袁旭音,叶宏萌,等.洪泽湖不同入湖河流沉积物磷形态特征及生物有效性 [J]. 中国环境科学, 2020,40(10):4568-4579.
Wan J, Yuan X Y, Ye H M, et al. Characteristics and bioavailability of different forms of phosphorus in sediments of rivers flowing into Hongze Lake [J]. China Environmental Science, 2020,40(10):4568-4579.
[24] Wu S, Zhao Y, Chen Y, et al. Sulfur cycling in freshwater sediments: A cryptic driving force of iron deposition and phosphorus mobilization [J]. Science of the Total Environment, 2019,657:1294-1303.
[25] Xie M, Alsina M A, Yuen J, et al. Effects of resuspension on the mobility and chemical speciation of zinc in contaminated sediments [J]. Journal of Hazardous Materials, 2019,364:300-308.
[26] Juottonen H, Fontaine L, Wurzbacher C, et al. Archaea in boreal Swedish lakes are diverse, dominated by Woesearchaeota and follow deterministic community assembly [J]. Environmental Microbiology, 2020,22(8):3158-3171.
[27] Guo X, Yang Y, Niu Z, et al. Characteristics of microbial community indicate anthropogenic impact on the sediments along the Yangtze Estuary and its coastal area, China [J]. Science of the Total Environment, 2019,648:306-314.
[28] Liu S, Meng J, Jiang L, et al. Rice husk biochar impacts soil phosphorous availability, phosphatase activities and bacterial community characteristics in three different soil types [J]. Applied Soil Ecology, 2017,116:12-22.
[29] Mao C, Li T, Rao W, et al. Chemical speciation of phosphorus in surface sediments from the Jiangsu Coast, East China: Influences, provenances and bioavailabilities [J]. Marine Pollution Bulletin, 2021, 163:111961.
[30] 王敬富,陈敬安,陈 权,等.深水水库磷的生物地球化学循环[J]. 第四纪研究, 2021,41(4):1194-1207.
Wang J F, Chen J A, Chen Q, et al. Biogeochemical cycles of phosphorus in deep-water reservoirs [J]. Quaternary Sciences, 2021,41(4):1194-1207.
[31] Field H R, Whitaker A H, Henson J A, et al. Sorption of copper and phosphate to diverse biogenic iron (oxyhydr)oxide deposits [J]. Science of the Total Environment, 2019,697:134111.
致谢:本实验的现场采样工作由贵州草海国家级自然保护区管理委员会协助完成,在此表示感谢.
Effects of microbial communities on phosphorus speciation in lakeside sediments of Caohai Lake.
LONG Yun-chuan1,2, JIANG Juan1,2, HU Jing2, ZHANG Zhen-ming1,2, ZHU Cheng-bin2, SU Rong-xiang1, ZHOU Shao-qi2*
(1.Guizhou Institute of Biology, Guizhou Academy of Sciences, Guiyang 550009, China;2.College of Resource and Environmental Engineering, Guizhou University, Guiyang 550025, China)., 2022,42(4):1869~1876
To explore the effect of the microbial community on phosphorus speciation in the sediment from plateau lake, the lakeside sediments from Caohai were collected, then the phosphorus speciation was analyzed, and the community compositions of bacteria and archaea were investigated using high-throughput sequencing. The results showed that the total phosphorus content in the sediments ranged from 662.89 to 881.26 mg/kg, with an average of 750.36mg/kg. Among thephosphorus fractions, NaOH- NRP>Res-P>BD-P>HCl-P>NaOH-SRP>NH4Cl-P. The bacterial community consisted of 59 phyla (dominated by Proteobacteria) and 1259 genera (dominated byand). The archaeal community consisted of 8 phyla (dominated by Crenarchaeota) and 67 genera (dominated by). The redundancy analysis, principal component analysis, and co-occurrence network analysis revealed that the microbial community could drive the transformation of the phosphorus speciation in lakeside sediments from Caohai. The bacteria affected the phosphorus species mainly through regulating the activity of alkaline phosphatase and the oxidation/reduction of iron, while the archaea impacted it by changing organic matter and acid phosphatase.
sediment;high-throughput sequencing;phosphorus fraction;co-occurrence network;archaea
X524
A
1000-6923(2022)04-1869-08
龙云川(1989-),男,重庆铜梁人,助理研究员,博士研究生,主要从事环境微生物研究.发表论文40余篇.
2021-09-06
国家自然科学基金资助项目(U1612441);贵州省科技计划项目([2020]4Y020,[2020]1Y168);贵州省生物研究所科技计划项目([2021]08)
*责任作者, 教授, zhousq@gzu.edu.cn
猜你喜欢
区域治理(2022年1期)2022-02-19
作物学报(2022年3期)2022-01-22
杭州(2021年15期)2021-09-08
杭州(2020年17期)2020-11-09
作文成功之路·小学版(2020年5期)2020-06-11
浙江林业(2020年12期)2020-01-11
学苑创造·C版(2019年9期)2019-11-07
分析化学(2018年7期)2018-09-17
天津农业科学(2014年12期)2014-12-11
当代贵州(2014年3期)2014-09-18
