
microRNA参与植物次生代谢与非生物胁迫的调控
2022-04-21 04:02:04李可欣田同同
中国生物化学与分子生物学报 2022年2期
李可欣, 田同同, 周 波,2)*
(1)东北林业大学生命科学学院, 哈尔滨 150040;2)东北盐碱植被恢复与重建教育部重点实验室(东北林业大学), 哈尔滨 150040)
植物中的 miRNA(microRNA)由MIR基因编码,其生物化学合成途径包括3个主要步骤:转录、加工和功能复合物装配。高通量测序等生物技术的进步和发展使鉴定大量编码基因和非编码RNA成为可能。成熟的miRNA长度大多为21 nt,它们与Argonaute (AGO)家族蛋白质结合,形成RNA诱导的沉默复合体(RNA-induced silencing complex,RISC)后进一步通过降解靶mRNA或抑制翻译来负调控基因表达水平[1-2]。
植物miRNA结合和调控靶基因表达的方式主要有3种类型:翻译抑制、剪切降解和介导植物DNA的甲基化。通常认为翻译抑制和剪切降解取决于miRNA与靶RNA之间互补配对的复杂性。序列几乎或完全互补时会引起靶mRNA的快速降解;当序列互补程度较低时,靶mRNA的翻译被直接抑制,有效达到蛋白质表达量降低的效果。此外,植物中的MIR基因还可以通过转录水平介导目标DNA甲基化的水平[3]。
植物在生长发育的过程中受到外部环境的影响,本文主要概述了参与非生物胁迫和植物次生代谢功能相关的miRNA及其靶基因的研究进展,以期更好的理解参与次生代谢和环境响应的miRNA的调控通路和功能。
1 植物微RNA与次生代谢
1.1 黄酮类生物合成相关miRNA
黄酮类化合物(flavonoids)是广泛分布于高等植物中的一类次生代谢产物,具有增强免疫力、预防及治疗心血管疾病、抑癌[4-5]、抗衰老和抗病毒等生理活性。研究证明,miRNAs主要通过调控参与黄酮类化合物合成过程的关键基因或转录因子,影响植物的多种生理进程[6]。近年来,花青素(anthocyanins)的合成调控得到了广泛研究,明确了MYB(v-myb avian myeloblastosis viral oncogene homolog)、SPL(squamosa promoter-binding protein like)等转录因子家族作为miRNA的靶标广泛参与花青素的合成。Li等[7]通过小RNA测序、KEGG通路分析等技术将miR858确定为奇异果花青素合成的负调控因子,miR858的过度表达导致其靶基因MYBC1编码的MYBC1转录因子表达量下降,不能启动与花青素合成关键酶无色花青素双加氧酶(leucoanthocyanidin dioxygenase,LODX)基因的表达,进而使得花青素的积累量下调。对茶树miR828a的研究表明,其能够靶向MYB4、MYB23和MYB82,进而负调节花青素的合成水平[8]。在过表达miR156的苹果[9]和白杨[10]中,也发现了多种转录因子MYB、SPL表达量下调,花青素水平变化的现象。因此,miRNA作为非编码的sRNA可以通过靶向转录调控基因,调控植物花青素合成途径,这为理解植物花青素合成的复杂调控网络提供了更多的线索。
1.2 萜类生物合成相关miRNA
青蒿素(artemisinin,ART)作为一种重要的次生代谢产物,其生物合成过程得到了广泛的研究。Khan等人[11]经研究发现,青蒿的miR159、miR172和miR166均靶向细胞色素P450还原酶(cytochrome P450 reductase,CPR)合成基因。CPR参与催化青蒿素合成的三个步骤。miR858的靶基因是编码紫穗槐二烯合酶(amorpha-4,11-diene synthase, ADS)的ADS。ADS也是参与青蒿素合成过程中的关键酶,在青蒿素合成通路中发挥着重要作用。极少种miRNA能够增强青蒿素的表达,如miR5083和miR403可正调控NAC1基因,上调ADS、细胞色素P450单加氧酶基因CYP71AV1的表达量。Saifi等人[12]发现,miRStv_11和miR319 g分别上调贝壳杉烯酸羟化酶(Kaurenoicacid-13-hydroxylase,KAH)和下调贝壳杉烯氧化酶 (Kaureneoxidase,KO)、贝壳杉烯合酶 (Kaurenesynthase,KS)的表达。KAH、KO、KS都是参与合成甜菊苷的重要酶类。
1.3 生物碱类生物合成相关miRNA
生物碱(alkaloid)是一类含氮小分子化学物质,种类繁多,合成途径复杂。pso-miR13和pso-miR 2161能分别抑制BIA生物合成过程中的关键酶7′-O-甲基转移酶 (7′-O- transmethylase,7-OMT)和4′-O-甲基转移酶(4′-O- transmethylase,4-OMT)基因,从而下调苄基异喹啉生物碱(benzylisoquinoline alkaloid,BIA)表达[13]。在马铃薯中鉴定发现,miR408靶向VS1环化酶(vetispiradiene cyclase)的合成基因,VS1的作用底物是糖苷生物碱合成过程的前体物质法尼基焦磷酸(farnesyl pyrophosphate,FPP)。miR408抑制VS1的合成导致底物FPP含量增加,从而使糖苷生物碱获得一定的积累。Chen等[15]通过研究发现,在火龙果中多种miRNA通过下调甜菜碱合成通路中目标基因的转录水平参与甜菜碱的积累过程。例如miR157b靶向SPL6、miR858靶向MYBC1和MYB12等。

Fig.1 The miRNA-mediated synthesis pathway of secondary metabolites in plants A variety of miRNAs affect the synthesis of secondary metabolites by promoting or inhibiting the expression of target genes in different plants. “—| ”means negative regulation. “→”means positive regulation
2 植物微RNA与非生物胁迫
2.1 植物miRNA与干旱胁迫
水是制约植物生存、发育和繁衍等各种生命活动不可或缺的重要因素。植物缺水时会导致体内水分平衡被破坏,出现光合速率减弱、呼吸作用异常、叶片萎蔫等常见的生理生化变化,严重时造成植物死亡。人们在研究后发现,miRNA在维持植物水分平衡、抵抗干旱带来的氧化应激影响中发挥着不可忽视的作用。研究证明,大部分植物在干旱条件下都有与正常生理环境差异表达的miRNA,它们极有可能在消除氧化应激影响中扮演着重要角色。有研究发现,大麦miR397的靶基因是漆酶基因LAC。漆酶能够催化合成木质素,被认为参与了植物的发育和逆境反应[16]。除此之外,Lpez-Galiano等人[17]的研究指出,miR159及其靶基因MYB33的表达模式相反。干旱胁迫模式下导致miR159下调和MYB33表达增强。miR166f是桑树抵抗干旱的正调控因子。过表达miR166f会使靶标同源异型域—亮氨酸拉链(HD-Zip)转录因子和具有组蛋白精氨酸去甲基化酶活性的JMJD6(Jmjc domain-containing 6,JMJD6)表达量下调,这种变化增强了桑树对干旱环境的适应性[18]。研究发现,在水分充足和干旱情况下西藏沙棘(Hippophaethibetana)2种新miRNA(novel_miR_24和novel_miR_87)的表达量发生了显著变化[19]。这在一定程度上为完善miRNA参与的耐旱植物复杂调控网络奠定了基础。研究表明,低到中等程度表达miR156可以沉默花青素合成关键酶二氢黄酮醇-4-还原酶((dihydroflavonol 4-reductase,DFR)编码基因的转录阻遏因子SPL13,增强紫花苜蓿的抗旱能力。
2.2 植物miRNA与温度胁迫
2.2.1 植物miRNA与高温胁迫 高温胁迫能够造成植物生物膜破坏,叶片出现死斑和萎蔫,光合速率和蒸腾速率下降,呼吸速率降低等多种不利影响。研究发现,在拟南芥中miR160可以对ARF17,ARF16和ARF10等靶基因的表达产生抑制作用,对增强热胁迫的适应力产生消极影响[21]。在玉米中也发现了几个关键的热响应miRNA及其对应的靶标。高温下miR172、miR164、miR396被抑制表达,导致靶向的AP2/ERF(APETALA2/ethylene-responsive factor)、NAC和生长调控因子(growth-regulating factor,GRF)下调,器官发育和信号转导等过程均被影响[22]。为阐明miRNA介导的玉米对高温胁迫的转录后调控提供了研究基础。Ahmed等人[23]对38℃热处理后的大白菜叶片组织RNA文库进行测序,得出miR172在高温胁迫下调控开花白菜中包括花期同源蛋白质AP2、乙烯响应转录因子在内的AP2-like转录因子表达的结论。棉花在产孢子的细胞增殖阶段、减数分裂阶段、小孢子释放阶段和花粉成熟阶段分别由多种miRNA及其靶基因响应干旱胁迫[24]。例如miR172与其靶基因AP2在减数分裂时期发挥防御作用,花粉成熟的最后阶段则由miR393-TIR1/AFB(transport inhibitor response 1/Auxin signaling F-box)积极响应。这些miRNA是棉花适应高温环境的主要和重要调控因子,具有正向或负向调控模式。
2.2.2 植物miRNA与低温胁迫 温度是影响植物生长发育和果实采后生活质量的主要环境因子。为了研究miR528提高水稻抗寒性的潜在分子作用机制,Tang等人[25]检测了胁迫相关MYB转录因子在miR528转基因水稻细胞中的表达量。研究证明,miR528通过抑制胁迫应答相关转录因子MYB30基因的表达,从而增强BMY2、BMY6和BMY10基因表达量增加,进而增强细胞活性、活化抗坏血酸过氧化物酶(ascorbate peroxidase,APX)等提高水稻对低温的耐受性。在香蕉中也发现,许多参与响应低温胁迫的miRNA及靶标例如SPL、蛋白激酶2(casein kinase 2,CK2)等[26,27]。它们参与信号转导,协调调控冷胁迫下机体的许多生化反应。也有研究发现,部分药用植物黄芪响应冷胁迫的部分基因调控网络,其中miR168通过靶向AGO基因在反馈调控中发挥重要作用。miR858、miR171、miR169等多个miRNA通过靶向多种转录因子例如SPL、MYB调控植物生长发育过程[28]。
2.3 植物miRNA与盐胁迫
盐分对植物的生长有着至关重要的作用,但超过一定限度,过量的可溶性盐对植物将产生毒害作用。过高的盐分容易引发次生胁迫,例如氧化应激反应和营养代谢失调,导致细胞损伤、生长抑制和作物产量下降等诸多问题的发生。铁超氧化物歧化酶(iron superoxide dismutase 1 ,FSD1)是棉花抵抗盐胁迫的重要酶。过表达miR414c的棉花FSD1表达量下降,使棉花抵御盐毒害的能力显著下降[29]。miR408在植物中稳定存在,核酸序列保守,过表达大麦miR408的烟草株系在磷饥饿和盐胁迫条件下有明显的表型变化,且miR408的靶基因磷酸转运体(phosphate transporter,PT)基因NtPT2、编码脱落酸受体和SnRK2蛋白质的基因NtPYL2和NtSAPK3转录丰度较野生型相比均有所上调,增强了大麦吸收磷的能力[30]。Ma等人[31]则通过研究验证了miR156a靶向SPL13转录因子,SPL13进一步靶向参与多种植物盐胁迫耐受生物通路的转录因子WRKY100的启动子区域,阐明了苹果miRNA响应盐胁迫的网络机制。另有研究分析了盐生植物红砂miRNA参与盐胁迫环境下种子萌发的具体机制。研究数据显示,miR4995和miR396分别靶向编码脱落酸8’-羟化酶的关键基因CYP707A2和生长素通路中的重要基因HB33,参与红砂种子的萌发[32]。
2.4 植物miRNA与重金属胁迫
铁、铜、锌等重金属在植物体内多种生理过程中发挥重要作用。然而重金属在植物体内的过度积累会毒害植物,影响植物的生长和生产力。此外,重金属的积累还会影响植物的生长和生产力,通过食物链富集对人和动物产生影响,引发疾病,诱导细胞损伤。铜元素是多种酶的辅因子,也是质体蓝素的重要组成部分。铜积累会制约植物的光合作用和新陈代谢等过程,导致细胞膜破坏、活性氧基团增加。研究发现,miR393及靶标运输抑制剂响应蛋白1 (transport inhibitor response 1,TIR1)、AFB1、AFB2和AFB3的相互作用,可能在葡萄维持铜离子稳态中扮演着重要角色[33]。在山核桃中也发现,能够靶向铜锌超氧化物歧化酶和线粒体细胞色素C氧化酶5b亚基的miR398[34]。抑制miR398的表达可以使超氧化物歧化酶浓度增加,清除氧自由基,减轻铜的毒害影响。镉元素也会对植物产生氧化毒害作用,镉的积累会导致植物发育不良,植株枯黄甚至死亡。Jian等人[35]研究认为,玉米中miR166靶向bZIP转录因子(basic region-leucine zipper,bZIP)、miR156b靶向Squamosa启动子结合蛋白(squamosa promoter binding protein,SBP),并且可能是通过负调控来维系玉米根系中Cd2+的稳态。在镉处理后的水稻中也发现,miR268的靶基因自然抵抗相关巨噬细胞蛋白3表达显著下降的现象[36]。
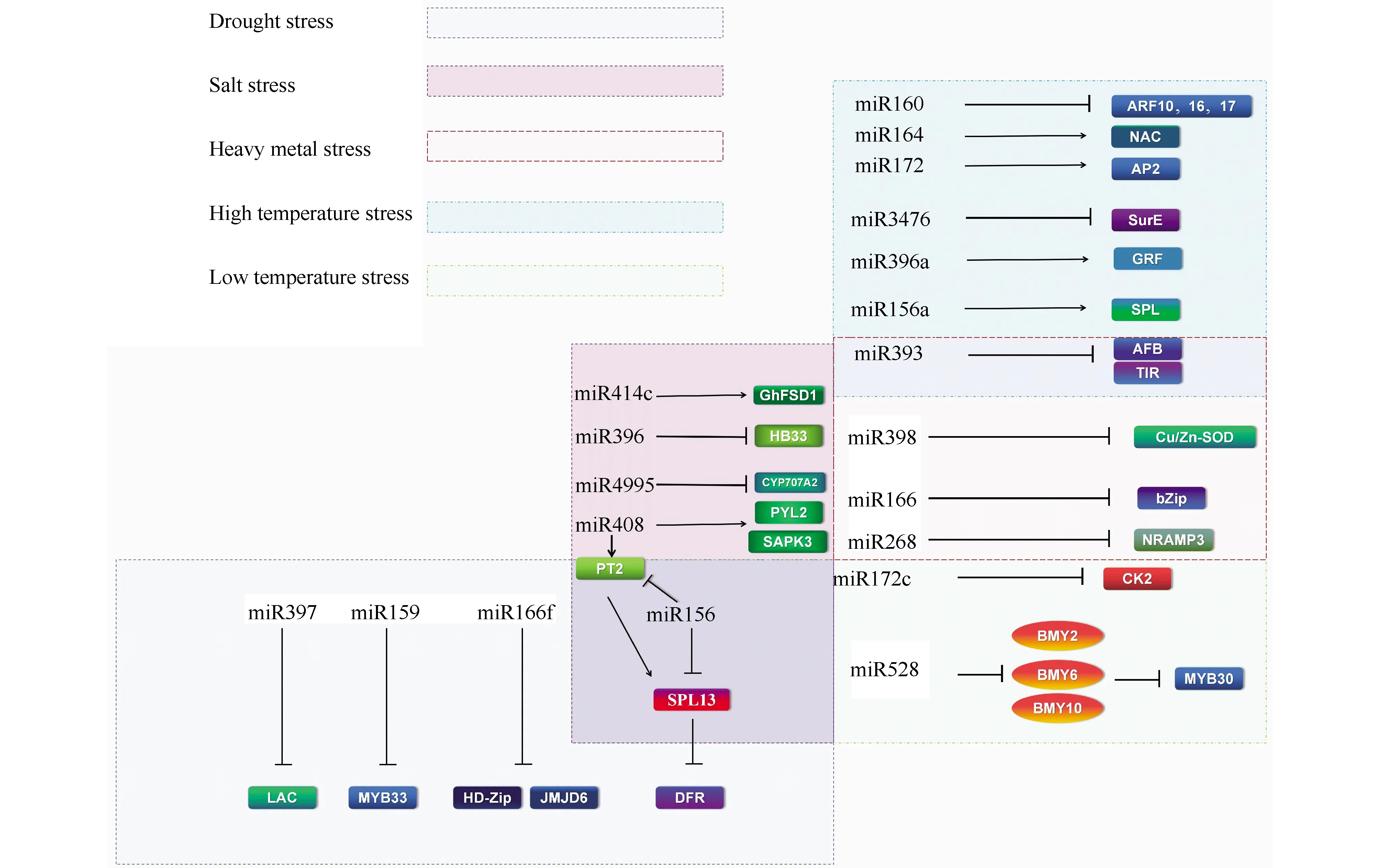
Fig.2 The miRNA-mediated pathway under abiotic stress Various miRAs are involved in response to abiotic stress by targeting different transcription factors in plants. These transcription factors play a positive or negative role in enhancing plant stress tolerance. “—| ”means negative regulation. “→”means positive regulation
3 次生代谢与非生物胁迫的联系
当植物受到外环境不利因素影响时,miRNA及次生代谢产物会发生相应变化以响应压力刺激,减轻活性氧基团对机体的毒害作用。
木质素、花青素等多种次生代谢产物被认为在植物应对生长和发育过程中对生物和非生物胁迫具有重要作用[37]。亚麻中次生代谢产物木质素的形成与miR397b、miR397及各自的靶基因表达相关。miR319靶向转录因子MYB,预测其调控植物木质素生物合成过程[38]。也有研究发现,甘薯参与花青素合成代谢的miRNA例如miR156、miR159大多数靶向参与响应非生物胁迫的转录因子例如MYB、SPL等,推测这些miRNA通过调控靶基因的表达量参与甘薯花青素的合成代谢,从而响应逆境胁迫[39]。在应激条件下,miR156-SPL9可能通过花青素着色产物因子PAP1(productionofanthocyaninpigment1 )和DFR途径直接影响花青素的生物合成,进而阻断花青素的生物合成,提高了植物对盐胁迫和干旱胁迫的敏感性[40]。HY5(elongated hypocotyl 5)转录因子能调节花青素生物合成途径中参与低温响应的关键成分查尔酮合成酶、查尔酮异构酶的积累。低温诱导下甜瓜miR319c表达量下降,从而促进HY5的增加[41]。
此外,某些非生物胁迫也会调控植物次生代谢产物的合成与降解。盐和干旱胁迫均可增强丹参miR396b的表达。过表达mR396b抑制丹酚酸的合成,促进了丹参酮的积累[42]。植物激素例如脱落酸、乙烯等也能通过调控信号传导过程参与植物响应逆境的生理过程。Sun等人[43]在盐胁迫处理的萝卜转录组中发现miR160b表达上调,其靶基因ARF16和ARF17在调节生长素表达过程中发挥作用。在甘薯、黄芪[44]和浮萍[45]中,也发现类似的miRNA靶向ARF,响应非生物胁迫的现象。这在一定程度上说明,miR160及其靶基因生长素响应因子ARF在响应非生物胁迫、调节生长发育过程中发挥重要的作用。综上所述,植物次生代谢与非生物胁迫具有密切联系。环境压力例如盐、温度会影响植物的代谢过程,调控次生代谢产物的表达。次生代谢产物合成后通过信号转导、氧化还原等机制缓解或清除活性氧基团等的毒害作用,提高机体的生存率。
4 问题与展望
miRNA作为基因调控网络中重要的调控因子,在协调次生代谢产物的生物合成和抵御环境压力的过程中扮演了重要角色。保守miRNA例如miR160在多种植物,例如甘薯、黄芪和萝卜中都靶向相似的基因家族,在非生物胁迫下发挥类似的作用[39,43,44]。非保守miRNA在不同植物中选择性表达,调控不同生理过程。随着生物技术的不断发展以及多领域交叉新兴学科的诞生,多种参与次生代谢产物合成的植物miRNA及其靶基因被鉴定和预测出来,为完善miRNA调控次生代谢网络,研究miRNA具体作用的分子机制提供了新方法和新思路。近年来,在不同种类的植物中也发现了大量参与响应非生物胁迫的miRNA。这些miRNA与目标基因的相互作用构成监管机体稳态网络的一个重要组成部分,控制各种生化反应以响应压力,增强自身适应环境的能力。目前,关于miRNA在参与合成次生代谢产物,以及次生代谢产物与非生物胁迫的联系方面的研究性文献较少,可供参考的数据不多,仍有较多植物尚未得到更深层次的探索。虽然已经发现了许多与响应环境压力和调控次生代谢过程有关的miRNA,但对于miRNA在不同植物中调控非生物胁迫和次生代谢产物生物合成的具体机制尚未得到全面的研究,且大多集中在药用植物和农作物中。次生代谢产物在植物清除氧自由基,缓解恶劣环境毒害作用的分子机制也未见统一的结论,需要更多可靠的证据来证明它们的真正作用。研究者们较热衷于挖掘与植物抵御恶劣环境的优良特性相关miRNA及其作用基因,以期改良植物特性,培育优势作物,增加作物产量。由于miRNA发现较晚,在植物代谢、胁迫响应通路中的研究较少,要想深入阐释其分子机制,还有待于我们进一步探索和发现。相信在生物技术更加先进的未来,植物miRNA的作用机制及调控网络能得到进一步的解读和完善,为培育优良品种做出贡献。
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:56
保健医苑(2022年5期)2022-06-10 07:46:38
中国临床医学影像杂志(2021年6期)2021-08-14 02:21:56
肝博士(2020年5期)2021-01-18 02:50:18
天然产物研究与开发(2019年10期)2019-11-05 10:12:44
天然产物研究与开发(2018年11期)2018-11-30 02:06:28
中成药(2017年5期)2017-06-13 13:01:12
中成药(2016年8期)2016-05-17 06:08:41
中国康复理论与实践(2015年10期)2015-12-24 05:42:43
医学研究杂志(2015年7期)2015-06-22 11:01:01
