
不同林龄橡胶林土壤细菌群落组成及多样性研究
2022-04-15 06:08:12孙树晴张希财兰国玉吴志祥
中国农学通报 2022年9期
孙树晴,丁 炜,孙 瑞,张希财,兰国玉,陈 伟,杨 川,吴志祥
(1中国热带农业科学院橡胶研究所,海南 海口 571101;2海南大学热带作物学院,海南 海口 570228;3陕西种苗有限公司,陕西 杨凌 712100;4农业农村部儋州热带作物科学观测实验站,海南 儋州 571737)
0 引言
亚洲热带地区的橡胶种植园迅速扩大,它们对区域生态系统的影响引起了人们的极大关注。新的橡胶种植园也经常被安置在对生物多样性保护和生态功能很重要的土地上。土壤微生物提供了重要的生态系统服务,以土壤细菌为主的土壤微生物其数量巨大、种类繁多,构成了地球上大部分的遗传多样性,土壤细菌可以转化分解植物凋落物并起着重要作用,进而可以被植物根系吸附。赵帆等[1]在研究草莓土壤微生物群落结构与土壤因子的关系中表明土壤微生物群落结构与土壤环境关系密切,其中全氮对微生物群落的影响最显著其次是pH。Fierer等[2]研究表明土壤pH含水量对不同条件下林地土壤细菌多样性有着重要影响。Gu等[3]在研究水稻土壤细菌时表明土壤环境的变化影响土壤细菌群落结构。荣新山等[4]在研究高寒高原细菌多样性中表明干旱影响土壤微生物群落多样性可以导致降低其多样性。顾松松等[5]在对不同类型茶园土壤细菌群落研究中发现三种茶园土壤细菌群落结构及多样性存在显著性差异。崔佩佩等[6]在对不同施肥处理对高粱根际微生物的研究中表明不同施肥处理对微生物群落功能多样性的影响不同。魏鹏[7]在对准噶尔盆地荒漠土壤进行研究中表明年降水量和有机碳是影响土壤细菌群落组成及其多样性的最重要的环境因子。靳晓拓等[8]在化肥减量配施有机肥对芒果园土壤的影响研究中表明施肥方式的改变会影响细菌的分布,细菌群落结构发生了变化但种类并没有改变。许广等[9]在对不同植茶年限茶树根际土壤细菌研究中表明植茶年限低的细菌多样性显著高于植茶年限高的茶树。梁田雨[10]在对浑善达克沙地榆树土壤微生物研究中表明物种多样性在春夏季显著大于秋季。Lan等[11]在橡胶园土壤细菌群落的驱动因素研究中表明历史的偶然性导致了细菌组成在地理尺度上的变化,而季节性的变化影响着局部尺度上的变化。由于土壤微生物群落与生态系统功能之间有着重要的联系,因此有必要更好地理解林龄以及环境因素是如何调节地下群落的动态和结构的。尽管在细菌群落这方面的研究很多但基于橡胶林的研究较少,更是鲜有对比不同林龄橡胶林土壤细菌群落组成及多样性的研究。需要更好地理解驱动橡胶林下土壤细菌组成和多样性的潜在机制。本研究选取海南儋州3种不同林龄橡胶林采用高通量测序技术研究其土壤细菌多样性和群落结构组成,并分析不同林龄橡胶树土壤理化性质与细菌群落及季节与细菌群落的相关性,微生物的快速生成时间意味着成分可以快速周转,即使是在不同季节也可以快速变化。因此,除了旱季和雨季外,笔者还检测了干旱和降雨交替时期(这里笔者称之为旱雨季节)的细菌组成和多样性,旱季选择3月、旱雨季节选择4月、雨季选择7月。目的是了解土壤细菌的季节性变化,并确定哪些环境因素可能对变化的多样性产生最大的影响。为改善橡胶树的土壤和提高胶树产量提供参考。
1 材料与方法
1.1 研究地与样品采集
采样地设立在位于海南岛西北部的儋州,海南岛属热带海洋性季风气候。11月至4月为旱季,5月至10月为雨季。年平均气温23.5℃,年平均降水量1815 mm;大约84%的降水是在雨季积聚的。
在中国热带农业科学院儋州实验场四队选择幼龄林(5年)和老龄林(25年)2种林龄的橡胶林,三队选择中龄林(10年)橡胶林。在2个实验场共建立9块样地(20× 20 m),3月(旱季)、4月底(旱雨交错期、旱雨季)和7月(雨季)采集0 ~20 cm深度土壤样品。在取样前去除凋落物层。每个小区用直径5 cm的不锈钢圆筒随机采集5个土壤样品。将每个小区采集的样品进行混合均匀化,每小区共有3个复合样品,最终得到27个复合土样。然后将复合土壤样品分为两部分:一部分风干,另一部分储存在-80℃,以备分析(DNA提取)。
1.2 土壤养分测定
土壤样品采用鲁如坤[12]规定的标准土壤试验方法进行分析。在1:1的土-水混合物中测定土壤pH值。土壤水分是用重量法测量的。采用凯氏定氮微消化、水蒸气蒸馏法测定土壤全氮(TN)。用氢氧化钠消化全磷(TP)和全钾(TK)。采用碱水解扩散法分析了碱解氮(Hy_N)[13]。土壤性质见表1。
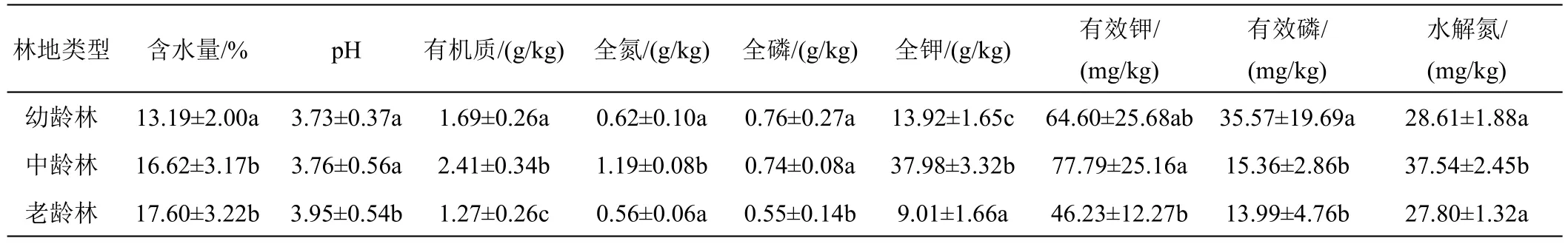
表1 不同林龄橡胶树土壤化学性质
1.3 DNA提取和扩增子测序
使用E.Z.N.A.®-soil DNA试剂盒(Omega Biotek,Norcross,GA,usa)按照制造商的协议从5.0 g土壤中提取微生物DNA。以338F(5′-ACTCCTACGGGA GGCAGCAG-3′)和 806R(5′-ggactachvgggtwtctaat-3′)为引物,采用热循环PCR系统[14](GeneAmp 9700,ABI,USA)扩增细菌16S rRNA基因V3-V4高变区。PCR反应使用以下程序进行:在95℃下变性3 min,在30饱和95℃下进行27个循环,在55℃下退火30 s,在72℃下延伸45 s,最后在72℃下延伸10 min。PCR反应一式三份,每份20 μL混合物,其中包含4 μL 5×FastPfu缓冲液,2μL 2.5 mmol/L dNTPs,0.8 μL 每种引物(5μmol/L),0.4 μL FastPfu聚合酶和10 ng模板DNA。
1.4 Illumina公司MiSeq测序
从2%琼脂糖凝胶中提取扩增子,使用AxyPrep DNA凝胶提取试剂盒(Axygen Biosciences,Union City,美国,CA)按照说明书进行纯化,并使用QuantiFluor-ST(Promega,美国)进行量化。根据标准方案,将纯化后的扩增子在Illumina MiSeq平台上进行等摩尔和配对测序(2250)。原始读取数据存入NCBI序列读取存档(SRA)数据库(序列号:SRP108394)。
1.5 数据统计与分析
利用Usearch软件(Usearch 7.0)将所有样本通过聚类操作,将序列按照彼此的相似性分归为许多小组,一个小组就是一个OTU。可根据不同的相似度水平,对所有序列进行OTU划分,通常对97%相似水平下的OTU进行生物信息统计分析。利用Mothur软件,计算各个样品的Ace指数、Shannon指数、Simpson指数、Coverage指数。采用主坐标分析(PCoA)对群落的门组成数据进行Bray-curtis distance分析,以解释各样本点微生物群落的相对相似性。利用相似性分析(ANOSIM)检验季节间分类学组成是否存在显著差异。为了揭示微生物群落与环境因子之间的相关性,利用R编程环境(http://cran.r-project.org/web/packages/vegan)中的纯素包,基于环境变量和群落的分类组成(门水平)数据进行了冗余分析(RDA)。9个土壤变量分别是土壤有机质(SOM)、全氮(TN)、全磷(TP)、全钾(TK)、含水量(WC)、土壤pH(pH)、有效磷(AP)、有效钾(AK)、水解氮(Hy_N)。采用SPSS 26.0对试验期土壤化学性质进行单因素方差分析。采用One-way ANOVA基于fdr算法进行多重检验校正,检验不同林龄橡胶林在组成上的显著性差异。通过双尾t-test检验确定两组之间的显著性,并且使用单因素方差分析(ANOVA)获得多组的显著性。
2 结果与分析
2.1 不同林龄橡胶树土壤理化性质分析
不同林龄土壤化学性质存在显著差异(表1)。由表1可知,随着林龄的增长,含水量与pH逐渐升高,幼龄林与中龄林、老龄林土壤含水量呈显著差异(P<0.05,下同),幼龄林、中龄林与老龄林pH呈显著性差异。3种林龄土壤有机质、全钾呈显著性差异,中龄林含量最高。幼、中龄林全磷含量与老龄林呈显著差异,随林龄增长全磷含量越低。有效钾和水解氮含量老龄林最低且呈显著差异。有效磷含量幼龄林与中、老龄林呈显著差异,且随着龄林的增长含量逐渐降低。
2.2 不同林龄橡胶树土壤细菌群落组成
由图1可以看出,在门水平上3种林龄橡胶林土壤细菌主要以变形菌门(Proteobacteria)相对丰度变化范围为:29.42% ~37.75%,中龄林>幼龄林>老龄林、酸杆菌门(Acidobacteria)相对丰度变化范围为:21.66% ~31.30%,老龄林>中龄林>幼龄林、绿弯菌门(Chloroflexi)相对丰度变化范围为:10.05% ~13.52%,老龄林>幼龄林>中龄林、放线菌门(Actinobacteria)相对丰度变化范围为:6.83% ~10.42%,幼龄林>中龄林>老龄林为主。在纲水平上3种林龄橡胶林土壤细菌主要以酸杆菌纲(Acidobacteria)相对丰度变化范围为:21.66% ~31.30%,老龄林>中龄林>幼龄林、α-变形杆菌纲(Alphaproteobacteria)相对丰度变化范围为:17.91% ~23.01%,中龄林>幼龄林>老龄林、放线菌纲(Actinobacteria)相对丰度变化范围为:6.83% ~10.42%,幼龄林>中龄林>老龄林、纤线杆菌纲(Ktedonobacteria)相对丰度变化范围为:2.97% ~6.47%,幼龄林>老龄林>中龄林。在属水平上3种林龄橡胶林土壤细菌norank_c__Acidobacteria相对丰度变化范围为:7.06% ~13.96%,老龄林>中龄林>幼龄林、norank_c__OPB35_soil_group相对丰度变化范围为:1.32% ~3.14%,幼龄林>老龄林>中龄林、变异性杆菌属(Variibacter)相对丰度变化范围为:1.77% ~2.81%,幼龄林>中龄林>老龄林,Candidatus_Koribacter相对丰度变化范围为:1.25% ~2.55%,老龄林>中龄林>幼龄林、norank_o__JG30-KF-AS9相对丰度变化范围为:1.29% ~2.64%,幼龄林>中龄林>老龄林、醋酸杆菌杆菌属(Acidibacter)相对丰度变化范围为:1.24% ~2.56%,中龄林>幼龄林>老龄林。结果表明:在门水平上变形菌和酸杆菌、在纲水平上酸杆菌纲和α-变形杆菌纲、在属水平上norank_c__Acidobacteria是3个林龄橡胶人工林土壤中最丰富的细菌群。基于样本中群落丰度数据,利用fdr计算方法,3种林龄土壤样品细菌群落丰度在门水平上酸杆菌门,绿弯菌门,放线菌门,厚壁菌门(Firmicutes),拟杆菌门(Bacteroidetes)呈显著性差异,硝化螺旋菌门(Nitrospirae)呈极显著性差异(P<0.01,下同)。在纲水平上3种林龄土壤样品中酸杆菌纲、放线菌纲、γ-变形菌(Gammaproteobacteria)、鞘脂杆菌纲(Sphingobactaiia)呈显著性差异,纤线杆菌纲、OPB35_soil_group、芽孢杆菌纲(Bacilli)呈极显著性差异(P<0.001,下同)。在属水平上norank_c__Acidobacteria、变异性杆菌属(Variibacter)、Candidatus_Koribacter、norank_o__JG30-KF-AS9呈显著性差异,norank_c__OPB35_soil_group、醋酸杆菌杆菌属(Acidibacter)呈极显著性差异。由结果不难发现,在3种林龄橡胶林土壤中无论是门水平、纲水平还是属水平上酸菌占据着绝对的优势。

图1 不同林龄橡胶树细菌群落组成(上为门水平,中为纲水平,下为属水平)
2.3 不同林龄土壤细菌群落结构特征
采用主坐标分析(PCoA)对细菌群落的门组成数据进行Bray-curtis距离分析,以解释各样本点微生物群落的相对相似性。PCoA第一主成分解释了群落结构差异的20.69%,第二主成分解释了群落结构差异的16.17%。老龄林占据着不同的坐标空间相对来说比较独立,幼龄林与中龄林分布较集中,说明幼龄林到中龄林未使得细菌群落出现显著的变化。PCoA结果还表明,在旱季和旱雨季,橡胶林土壤中的细菌在门水平上没有显著差异(图2)(P>0.05,下同)。
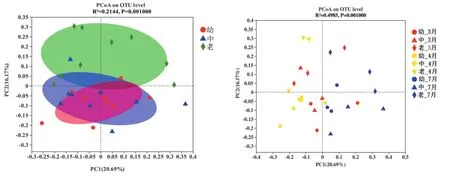
图2 不同林龄橡胶树土壤细菌群落的主坐标轴(PCoA)分析
基于Bray-curtis计算各组距离利用相似性分析(ANOSIM)来检验季节之间的分类组成是否有显著差异。ANOSIM结果证实旱雨季(4月)和雨季(7月)呈显著性差异(图3)。

图3 旱季和旱雨季土壤样品细菌群落相似性(ANOSIM)
2.4 不同林龄橡胶树土壤细菌群落Alpha多样性
通过对3种林龄橡胶树土壤细菌群落多样性指数(表2)分析发现Coverage指数均在95%以上,表明本次测序结果可以反映样本中细菌群落的真实情况。Sobs指数指样本中观察到的物种数目。3种林龄橡胶树样品Sobs指数老龄林>幼龄林>中龄林。Ace指数可反映群落物种丰富度,Simpson指数表示样品的均匀度,3种林龄橡胶树样品Ace指数和Simpson指数均为老龄林>中龄林>幼龄林。Shannon:用来估算样本中微生物多样性的指数之一。Shannon值越大,说明群落多样性越高。3种林龄橡胶树土壤样品Shannon指数是幼龄林>中龄林>老龄林。表3显示,3种林龄土壤细菌群落Ace指数和Shannon指数均表现出旱雨季(4月)和雨季(7月)呈现显著性差异。3种林龄橡胶树土壤样品Simpson指数在季节上无显著性差异。由表3不难发现Ace指数和Shannon指数在幼龄林和中龄林都呈现出旱雨季(4月)指数值最高,而老龄林则与之相反。Sobs指数雨季(7月)与其他两个季节均呈显著性差异,幼龄林和中龄林旱雨季指数最高而老龄林则是旱季最高。3种林龄橡胶树土壤样品雨季多样性指数低于旱季。

表2 不同林龄橡胶树土壤细菌群落多样性指数

表3 不同林龄橡胶树土壤细菌群落多样性指数在季节上的变化
2.5 不同林龄橡胶树土壤环境因子与微生物群落的相关性
图4显示土壤有机质与绿弯菌门、厚壁菌门呈显著负相关,pH与酸杆菌门和芽单胞菌门呈显著正相关,与芽单胞菌门甚至达到极显著性差异,有效磷与拟杆菌门呈极显著正相关,水解氮与变形菌门呈显著正相关。说明土壤细菌群落结构与土壤环境关系密切[1-15]。

图4 不同林龄橡胶树土壤细菌与环境因子相关性热图
RDA的第一轴和第二轴分别解释了44.81%和11.86%,共同解释了门丰度总方差的56.67%。季节解释了24.60%,水解释了1.34%,pH解释了10.06%,有机质解释了7.81%,全氮解释了2.43%,全磷解释了3.27%,全钾解释了5.19%,有效钾解释了5.36%,有效磷解释了3.10%,水解氮解释了7.51%。不同的土壤因子对土壤微生物数量、群落结构及其多样性的影响有所不相同[16],季节对微生物群落影响最大,在这里全氮对其影响最低。水解氮也是影响细菌群落组成的一个因素(图5),这与Lan等[13]结果一致。

图5 不同林龄橡胶树土壤细菌群落组成与环境因子的冗余分析
3 讨论
有研究表明变形菌丰度的高低与土壤中有机质含量的高低有密切的关系[17-19],本研究显示变形菌和酸杆菌为优势菌,变形菌丰度为中龄林>幼龄林>老龄林,与本研究结果一致。也有研究表明[20-21]变形菌门为土壤细菌群落结构中丰度最高的门,本研究中变形菌的丰度次于酸杆菌,与本研究结果不一致,出现该结果的原因可能是由于海南土壤偏酸性造成的。这或许将会成为我们接下来研究的重点之一。3种林龄土壤样品细菌群落丰度无论在门水平上、纲水平还是属水平上酸菌占比较大。酸杆菌门的丰度为老龄林>中龄林>幼龄林,3种林龄橡胶树土壤都呈酸性,幼龄林酸度最高,老龄林酸度最低,而本研究中酸杆菌门含量最高的林龄却是老龄林。酸杆菌门嗜酸,其丰度足以表明土壤的酸性条件,而本研究的结果却与此结论恰恰相反,为什么会出现这种情况需要做进一步的研究。
3种林龄橡胶树样品丰富度指数(Ace指数)和优势度指数(Simpson指数)均为老龄林>中龄林>幼龄林,多样性指数(Shannon指数)是幼龄林>中龄林>老龄林。有研究表明[22]老龄林多样性指数和丰富度指数均高于幼龄林,而本研究中只有丰富度指数与其研究结果一致。在优势度指数(Simpson指数)上邢慧等[22]的研究结果表明老龄林最高,与本研究结果一致。3种林龄橡胶树土壤细菌群落观察指数(Sobs指数)丰富度指数(Ace指数)和多样性指数(Shannon指数)均表现出旱雨季和雨季呈现显著性差异。3种林龄橡胶树土壤样品优势度指数(Simpson指数)在季节上无显著性差异。丰富度指数(Ace指数)和多样性指数(Shannon指数)在幼龄林和中龄林都呈现出旱雨季指数值最高,而老龄林则与之相反。由此可见季节变化也是影响细菌群落多样性的因素之一,这与Lan等[13]的研究结果一致。
秦越等[23]表明土壤细菌群落的组成受许多环境因素共同作用的影响。土壤理化性质是影响土壤细菌群落微生物种群数量[24]和结构组成[25]。本研究中随着林龄的增长pH逐渐增长,有研究[9,26]认为植树年限的增加极易导致土壤酸化的观点不一致。这两篇前人的研究与本研究的地理位置和地上植被均不同,因此导致此结果的原因究竟是由于海南土壤偏酸化或者是地上植被不同还是其他原因,具体的还需要我们做进一步的研究。季节影响细菌群落结构的主要环境因子,这与He等[27]研究结果一致,pH是影响微生物群落结构的次要因子,这与侯建伟等[28]和赵帆等[1]的研究结果有所不同。全磷和有效磷含量随着林龄的增长逐渐降低,王俊华等[29]研究表明增加有效磷可以提高微生物多样性,因此随着林龄的增长要注意增加土壤磷含量。有研究表明[30]土壤湿度是土壤微生物群落的重要驱动因素,水分含量的差异是土壤细菌群落组成变化的原因[31]。本研究冗余分析表明土壤含水量对我们分析的细菌群落的组成的影响不大,解释度仅为1.34%。不同的微生物类群对各环境因子的响应特征各异,土壤有机质与绿弯菌门、厚壁菌门呈显著负相关,pH与酸杆菌门和未分类菌门呈显著正相关,与芽单胞菌门呈极显著正相关,有效磷与拟杆菌门呈极显著正相关,水解氮与变形菌门呈正相关。由此可以看出氮、磷等不仅影响植物的生长发育同样也会对土壤细菌群落结构及其多样性产生较大影响,这与严绍裕[32]研究结果一致。
随着经济的快速发展,橡胶的社会需求量也在逐步增长,促使橡胶树的种植面积也不断扩大。但由于地力的不断消耗和农业经营管理措施的不当,导致橡胶树病虫害的不断增加,迫使橡胶的产量下降。近年来,随着研究的日益深入,对土壤微生物群落结构及其影响因素、土壤微生物结构与生态功能的关系,土壤微生物在维持土壤质量等方面的研究越来越受到土壤学家、生态学家和微生物学家的关注。土壤微生物在地球化学循环及维持土壤肥力和作物产量方面发挥了重要作用。因此作为相关研究人员,这也是我今后不断努力研究的不竭动力与研究方向。
4 结论
(1)在门水平上变形菌和酸杆菌、在纲水平上酸杆菌纲和α-变形杆菌纲、在属水平上norank_c__Acidobacteria是3个林龄橡胶人工林土壤中最丰富的细菌群,酸杆菌在3种林龄橡胶树土壤中呈显著性差异。
(2)3种林龄橡胶树土壤细菌群落多样性指数无显著性差异。但Sobs指数雨季(7月)与其他两个季节均呈显著性差异,幼龄林和中龄林旱雨季指数最高而老龄林则是旱季最高。Ace指数和Shannon指数在幼龄林和中龄林都呈现出旱雨季指数值最高,而老龄林则与之相反。
(3)土壤微生物群落结构多样性与土壤理化性质有关,季节解释了24.60%,水解释了1.34%,pH解释了10.06%,有机质解释了7.81%,全氮解释了2.43%,全磷解释了3.27%,全钾解释了5.19%,有效钾解释了5.36%,有效磷解释了3.10%,水解氮解释了7.51%。季节是影响群落结构及多样性的重要因素。
猜你喜欢
乡村科技(2023年20期)2024-01-08 07:20:00
热带生物学报(2022年1期)2022-03-09 10:53:58
农业科技与信息(2021年9期)2021-12-07 01:53:49
农业与技术(2018年16期)2018-11-28 11:24:08
森林工程(2018年5期)2018-05-14 13:54:30
现代农业科技(2017年10期)2017-07-12 14:14:49
贵州科学(2016年5期)2016-11-29 01:25:31
世界热带农业信息(2016年6期)2016-07-02 06:33:24
广西林业科学(2016年2期)2016-03-20 05:53:41
中国农业信息(2016年17期)2016-02-06 09:50:05
