
广州闽楠-樟人工混交林物种组成与多样性研究
2022-04-14 06:03:42程欣欣洪维邓双文魏雪莹张干荣冯蔚郭勇叶华谷王发国
热带亚热带植物学报 2022年2期
程欣欣, 洪维, 邓双文, 魏雪莹, 张干荣, 冯蔚, 郭勇, 叶华谷,王发国*
广州闽楠-樟人工混交林物种组成与多样性研究
程欣欣1,2, 洪维3, 邓双文2, 魏雪莹2, 张干荣3, 冯蔚3, 郭勇3, 叶华谷2,王发国2*
(1. 仲恺农业工程学院,广州 510225; 2. 中国科学院华南植物园, 广东省应用植物学重点实验室, 广州 510650;3. 广东省龙眼洞林场,广州 510520)
为了解人工混交林群落结构特征和物种多样性关系,对广州龙眼洞林场内闽楠()-樟()人工混交林进行群落结构分析。结果表明, 0.37 hm2样地内共有128种植物9 563株,隶属于57科99属;其中乔木35种,灌木61种,草本98种。在乔木层中,重要值排名前5的植物分别是山乌桕()(33.86%)、闽楠(17.24%)、樟(12.96%)、三桠苦()(7.92%)和山苍子()(3.70%);灌木层以茜草科(Rubiaceae)植物为主;草本层以禾本科(Poaceae)植物为主。乔木层、灌木层和草本层的Shannon-Wiener指数分别为1.230、1.708和1.754,Simpson指数分别为0.635、0.680和0.707,表明该人工混交林下草本物种丰富。非度量多维尺度分析(NMDS)表明,不同样方的物种组成相似性并不一致,部分样方含有偶见种。群落的径级结构呈倒“J”型,其中11.64%为幼苗,苗木占比6.08%, 小树占比55.34%,壮树占比25.28%,大树占比仅为0.70%,因此推测闽楠-樟人工混交林仍处于演替初期。
闽楠-樟人工林;物种多样性;群落结构;群落演替
城市化快速发展给自然环境带来了严重的破坏,城市生态平衡面临着严峻的考验。而城市森林作为城市生态系统的重要组成成分,被喻为城市之肺,在空气污染物清除、碳储存和碳螯合、降温与节能、生物挥发物排放以及水文效益等方面具有独特生态功能[1],在城市复合生态系统中具有不可替代的作用。城市森林群落的物种组成、结构和物种多样性,是认识、管理森林进而保证城市生态健康的基础[2]。物种多样性是群落组成结构和稳定性的重要指标[3],是维持生态系统功能和提供生态服务的重要组成部分[4]。研究城市森林的群落结构和生物多样性,能够为人工林的建设提供树种选择和配置,为城市森林的科学管理提供理论依据[5]。
闽楠()是樟科(Lauraceae)的常绿大型乔木, 耐阴, 忌强光[6],是高级建筑、家具、雕刻和精密木模的珍贵用材树种。樟()是樟科的常绿大型乔木;木材为造船、橱箱和建筑等用材,根、枝、叶可提取樟脑和樟油, 供医药及香料工业用;果核含脂肪,为重要工业用油。闽楠和樟树不仅为国家二级重点保护野生植物,还具有较高的观赏价值,是著名的庭园观赏和城市绿化树种[7]。
广州市现有森林多为人工林,而人工林培育中森林长期生产力、土壤肥力、林分抗性等已成为人们关注的热点问题[8]。目前对于闽楠、樟人工林混交比例[9]、生长状况[10]的研究已有相关报道,而对于闽楠-樟人工林群落结构的研究少有报道。本文对广州市龙眼洞林场闽楠-樟人工林优势树种的生长情况进行了调查,分析了林下的物种组成和生物多样性, 旨在为城市森林物种多样性保护、人工林的经营管理、森林资源可持续利用提供依据,以推进人工林建设。
1 试验地概况
广东省龙眼洞林场位于广州市东北部,经营面积达1 620.3 hm2,森林面积达1 560.2 hm2,生态公益林占林业用地的60%[11]。研究地点位于林场筲窝区域,位于13°20′~113°27′ E,23°11′~23°18′ N,属于亚热带海洋性季风气候区, 年均温为21.8 ℃, >10 ℃的年积温达8 000℃以上,最热月(7月)均温为29.6℃,最冷月(1月)均温为13 ℃。年降雨量为1 760 mm,雨季集中在4—9月,年均日照时数1 820~1 960 h。土壤主要为赤红壤,主要由花岗岩和砂页岩发育而成,pH值4.75~5.20,具有良好的透水性。
2 方法
2.1 样地设置
2020年5月,采用典型抽样的方法,于广州市龙眼洞林场筲箕窝区域闽楠-樟人工混交林内布设10 m×10 m的样方37个,共0.37 hm2。对样方内胸径(DBH)≥2 cm的乔木进行每木调查,记录种名、树高、胸径和冠幅等;在每个样方内设置1个5 m× 5 m的灌木层样地和4个2 m×2 m的草本层样地,记录灌木和草本的种名、高度和数量等。
2.2 数据的统计和分析
2.2.1 群落数量特征[12]
重要值(IV)=(相对显著度+相对频度+相对多度)/3;相对多度=(某种植物的个体数/所有种个体数总和)×100%;相对频度=(某种植物的频度/所有种的频度总和)×100%;相对显著度=(某种植物胸高断面积/所有种胸高断面积总和)×100%。
2.2.2 物种多样性计算
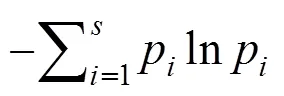
2.2.3 群落排序分析
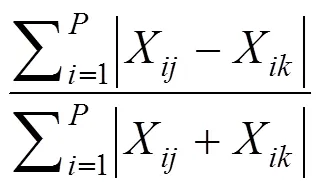
2.2.4 群落优势种群径级划分标准[17]
Ⅰ级(幼苗): 高度<33 cm;Ⅱ级(苗木): 高度≥33 cm, DBH<2.5 cm; Ⅲ级(小树): 2.5 cm≤DBH< 7.5 cm; Ⅳ级(壮树): 7.5 cm≤DBH<22.5 cm; Ⅴ级(大树): DBN≥22.5 cm。
3 结果和分析
3.1 群落的物种组成
闽楠-樟人工混交林中,37个样方共记录到存活植物128种9 563株,隶属于57科99属。其中,蕨类植物8科10属15种,裸子植物1科1属1种,被子植物48科88属112种。含属数最多的科为大戟科(Euphorbiaceae, 10属, 下同),其次是禾本科(Poaceae, 8)、茜草科(Rubiaceae, 6)、百合科(Liliaceae, 5)、蔷薇科(Rosaceae, 5)、山茶科(Theaceae, 5)、樟科(Lauraceae, 5)和紫金牛科(Myr- sinaceae, 5)。
乔木层有35种植物,隶属于20科31属,共842株。重要值(IV)≥1%的有12种(表1),其中大于10%的有3种:山乌桕(, 33.86%)、闽楠(17.24%)和樟(12.96%),另外三桠苦()、山苍子()、红锥()、鹅掌柴()锥栗()华南毛柃()壳菜果()等的IV也较大,在群落中发挥重要作用。
灌木层有61种植物,隶属于37科56属,其中茜草科植物最多(4属4种),有九节()、玉叶金花()、龙船花()和牛白藤()。
草本层有98种植物,隶属于50科79属,以禾本科植物最多(6属8种),有蔓生莠竹()、芒()、五节芒()、小花露籽草()、短叶黍()、弓果黍()、散穗弓果黍()和淡竹叶()。
另外,群落中还有4种珍稀濒危植物,分别为巴戟天()、宽叶线柱兰()、钳唇兰()和金毛狗()。
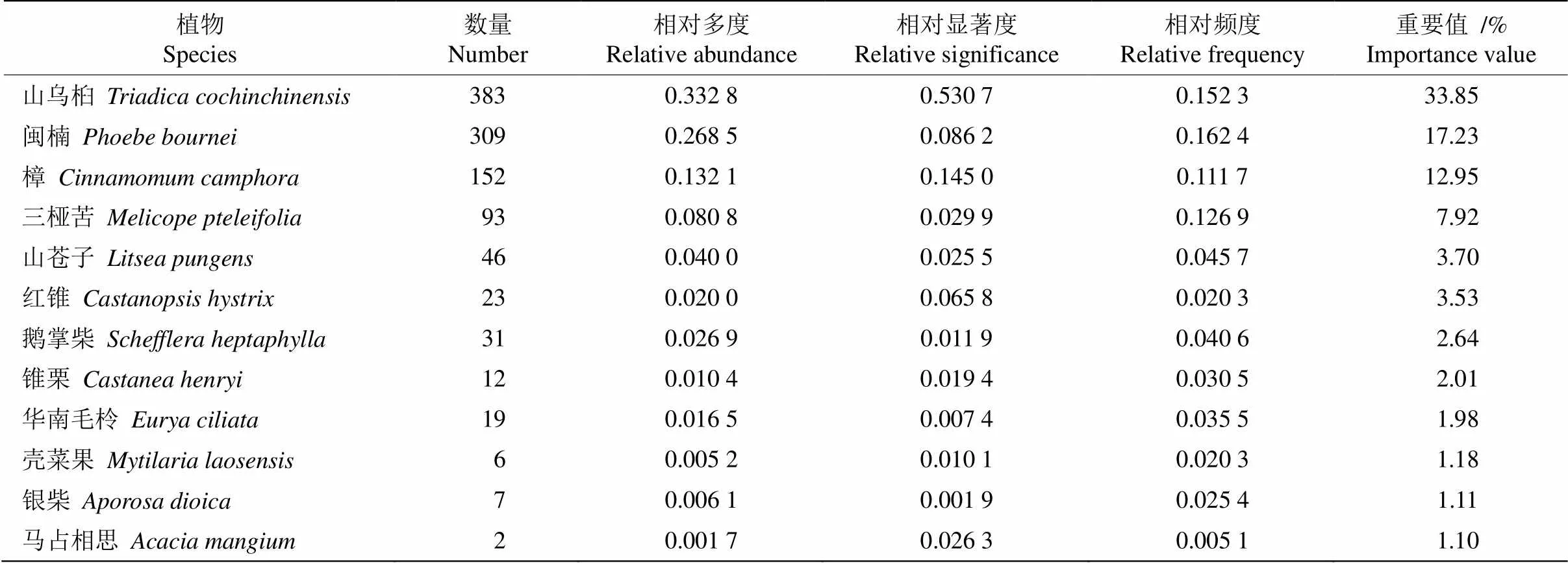
表1 乔木层中IV≥1%的物种
3.2 群落的物种多样性
对样方内的物种数量进行统计(图1),样方1、2、3、14、29的物种均大于25种,占总物种数高于20%;少数样方仅有闽楠和樟2物种,物种丰富度低。采用非度量多维标度分析法对样方间物种组成的相似性进行排序分析和评估(图2, 3),结果非线性拟合度2=0.968,线性回归2=0.852,表明拟合效果较好,排序结果可靠。从图2可见,大多数样方的物种分布较为集中,其中样方1、2、3、14和29间的距离较近,这几个样方内的物种组成相似性更高,与统计结果一致;样方33、34、36、37与其他样方相距较远,仅集中在图2右侧区域,表明林下物种组成和数量特征与其他样方的差异性较大,可能是这几个样方由于分布在道路旁,受人为活动干扰较大,从而导致物种组成单一。而苍白秤钩风()、锈毛莓()、粪箕笃()、蓝树()、楤木()在图2中处于边缘,与其他物种距离较远,表明其为群落中的偶见种,仅在少数样方中出现。
分别对群落各层级的物种多样性指数进行计算(表2),不同层级的物种丰富度、Shannon-Wiener指数、Simpson指数表现出相同的变化趋势,均为草本层>灌木层>乔木层,表明草本层中物种数量最多, 最丰富,优势度与多样性较高。而Pielou指数则呈现不同的趋势,表现为乔木层>草本层>灌木层,表明乔木层中物种分布更均匀。将闽楠-樟群落与广东省同属亚热带季风气候的其他植物群落[2,18–19]进行物种多样性比较(表3),结果表明,闽楠-樟群落的物种丰富度较高,而乔木层多样性指数(ʹ)较低。
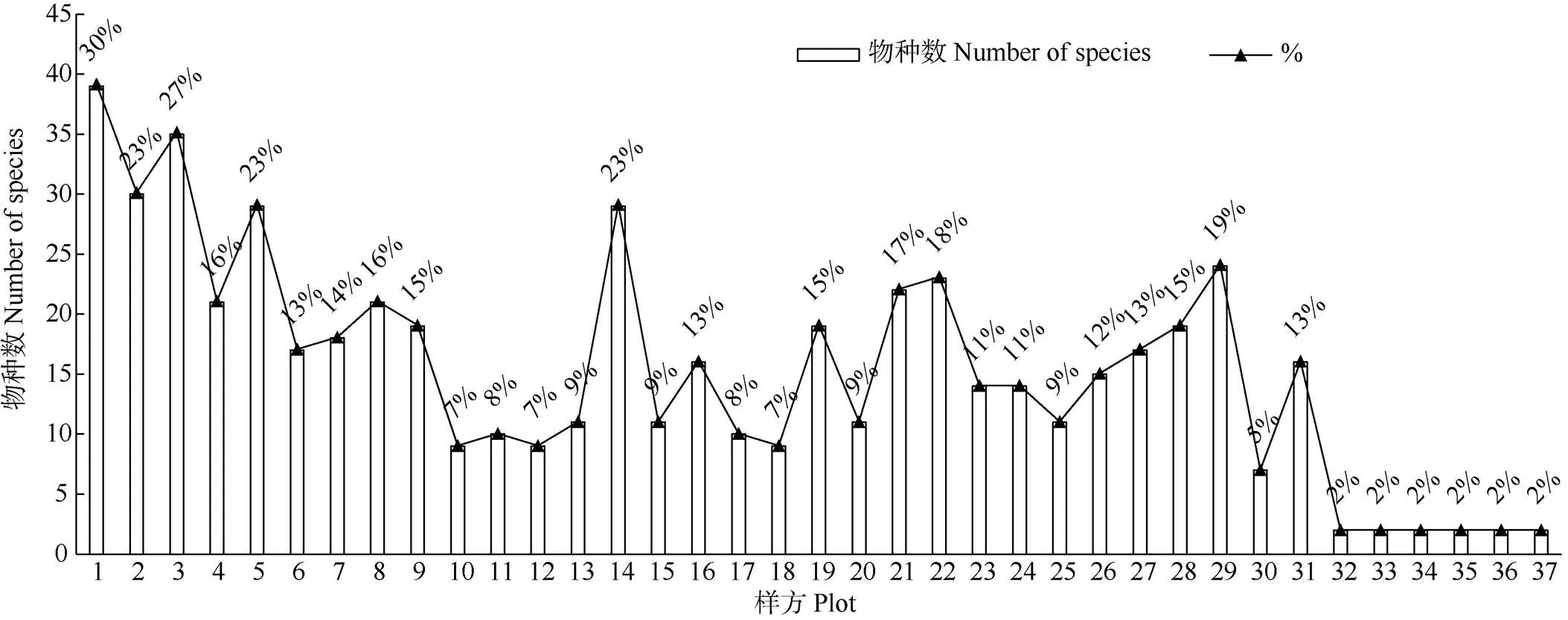
图1 样方中的物种数量

图2 基于物种多度Bray-Curtis相异矩阵数据的NMDS双序图
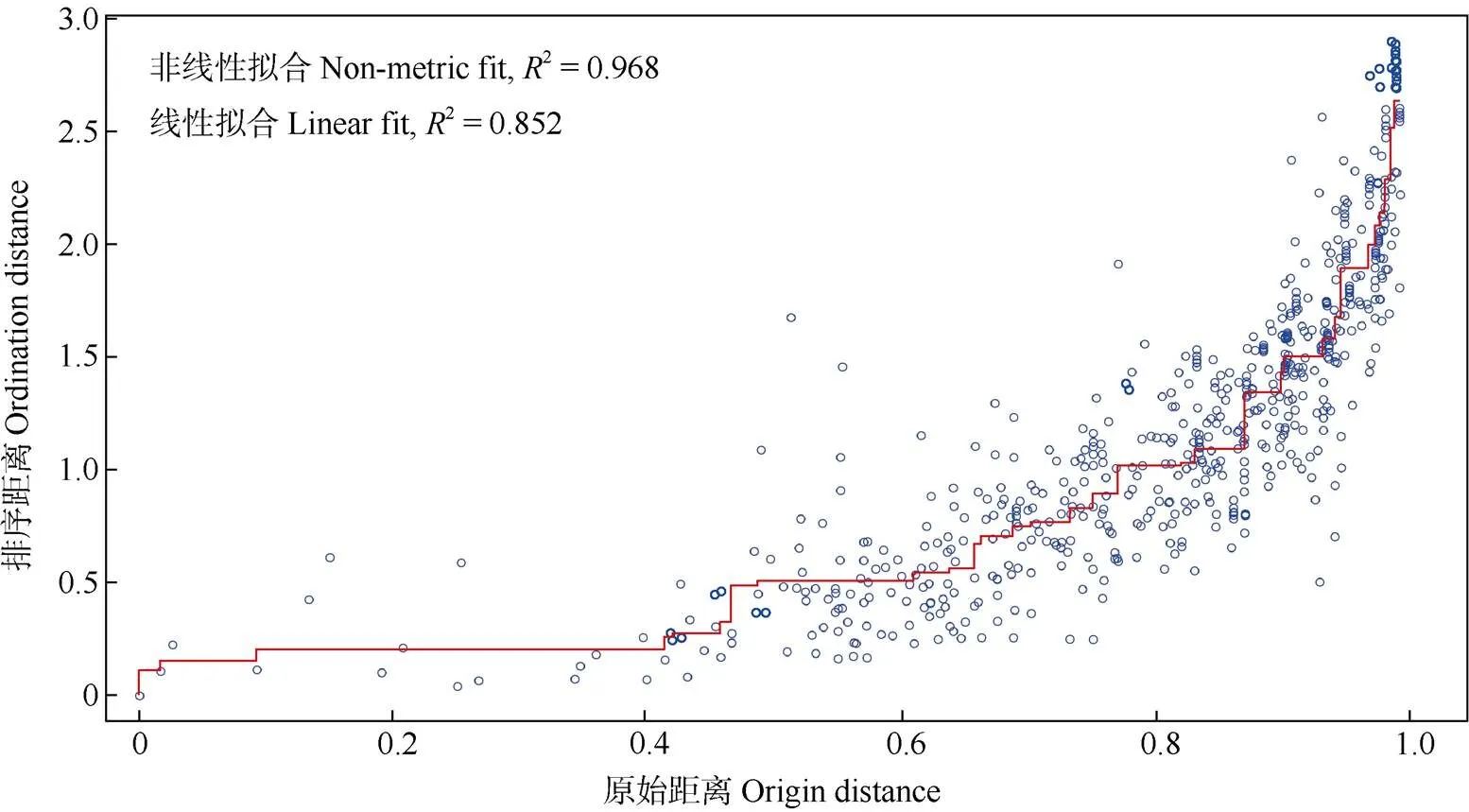
图3 检验NMDS结果的Shepard图
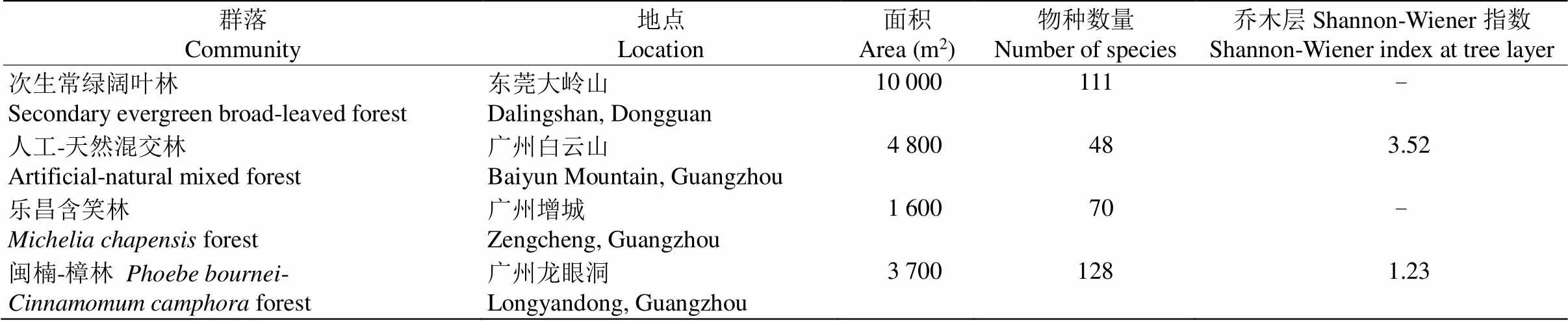
表3 不同植物群落的物种多样性比较
3.3 群落的径级结构和演替
对闽楠-樟群落中乔木层物种的径级结构进行划分,其中高度<33 cm的幼苗有145株,占11.64%;高度≥33 cm,DBH<2.5 cm的苗木有20株,占6.08%;2.5 cm≤DBH<7.5 cm的小树有637株,占55.34%;7.5 cm≤DBH<22.5 cm的壮树有291株,占25.28%;DBH≥22.5 cm的大树有8株,占0.70%。乔木层平均胸径为6.15 cm,随着径级的增加,乔木数量逐渐减少,于DBH为8~12 cm时,出现1个小双峰,符合倒“J”型分布(图4)。
进一步对群落主要建群种的生长情况进行评估(表4),山乌桕的胸径、树高、冠幅均大于闽楠和樟,且密度更高,重要值更大(33.86%),闽楠、樟重要值略低,表明山乌桕在群落中占最大优势,闽楠、樟为次优树种。另外,从图5可见,山乌桕的径级结构以7.5 cm≤DBH<22.5 cm的壮树最多(196株),占群落物种总数的17.03%,而闽楠和樟的径级结构以2.5 cm≤DBH<7.5 cm的小树较多,分别有193和102株,占群落总株树16.77%和8.86%,表明闽楠和樟处于增长阶段,而山乌桕处于稳定期。
4 结论和讨论
对龙眼洞筲窝的闽楠-樟人工林群落进行植物样方调查,在0.37 hm2样地内共记录57科99属128种植物。与其他森林群落相比,比白云山人工-天然混交林群落(48种)[2]、大岭山次生常绿阔叶林群落(111种)[18]、增城乐昌含笑林群落(70种)[19]的多,物种丰富度更高。乔木层、灌木层和草本层分别有35、61和98种植物,灌草层对群落的物种丰富度贡献更大,可能是由于广州龙眼洞林场位于亚热带沿海地区,高温多雨的气候环境促进了林下物种的自然更新。并且闽楠-樟人工林位于森林面积达1 560.2 hm2的龙眼洞林场内,大面积的常绿阔叶林为林下物种丰富度、生物多样性的提高创造了有利条件。珍稀植物的出现更是促进了林中植物多样性的发展[20]。
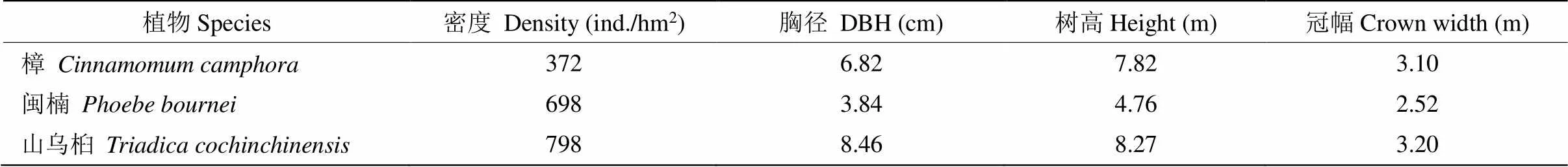
表4 群落中主要建群种的生长情况
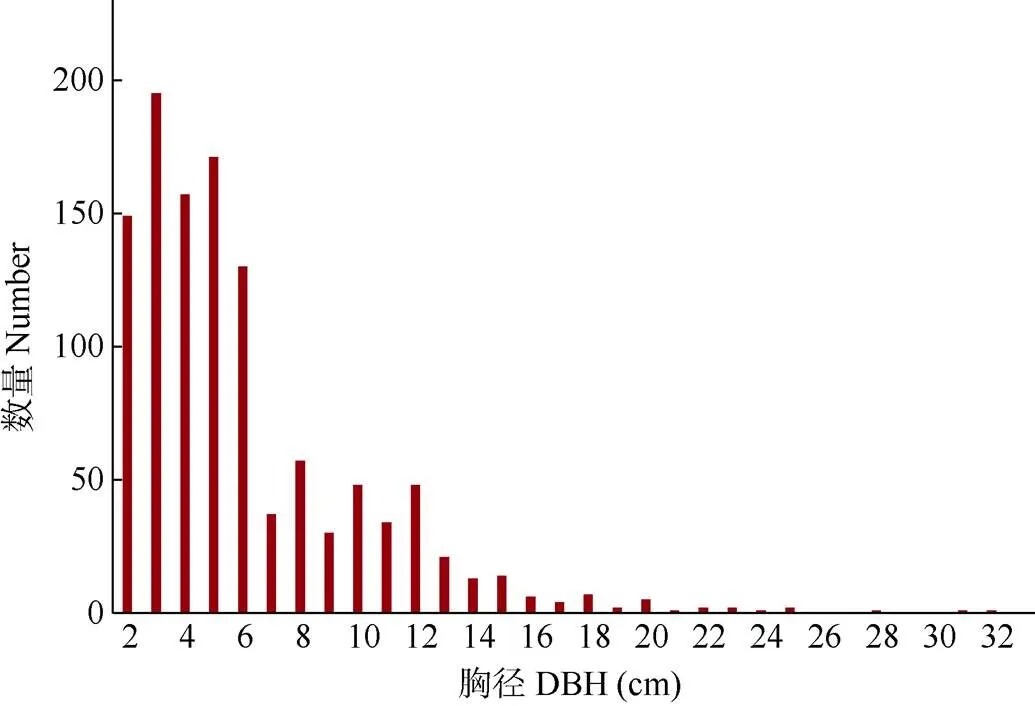
图4 群落的径级结构分布
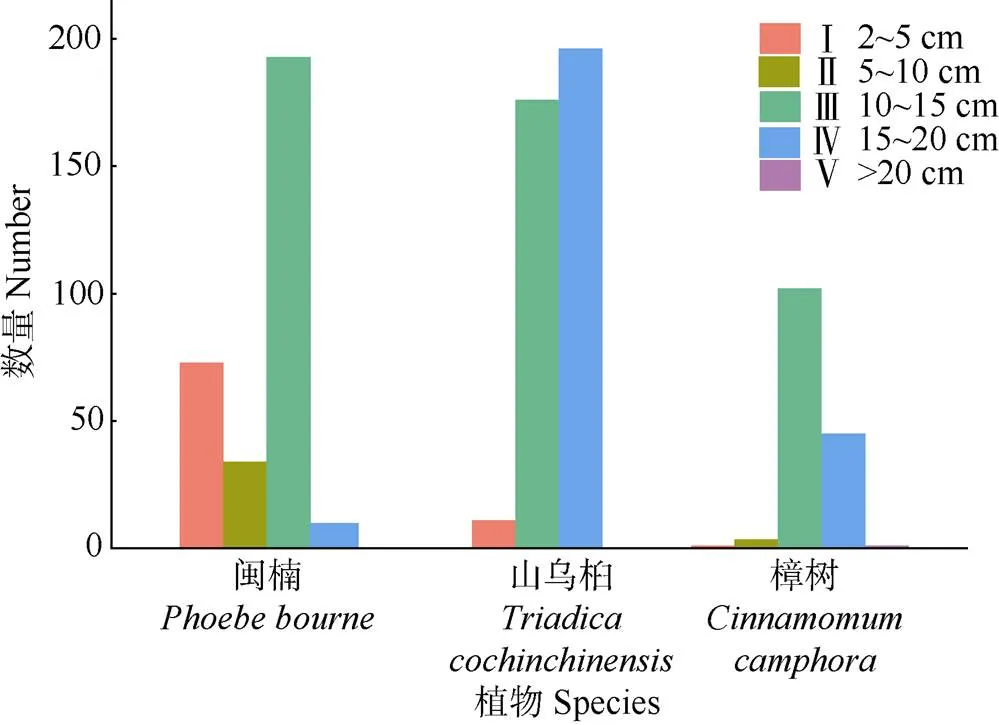
图5 群落中主要树种的径级结构
山乌桕、樟及闽楠为该群落的主要建群种,其中山乌桕为优势种,其次是樟及闽楠。在年龄结构方面,樟和闽楠同一时间种植,樟的平均胸径和树高分别为6.82 cm和7.82 m,闽楠为3.84 cm和4.76 m,表明樟的生长状况要优于闽楠,这可能是因为樟树适应能力较强,而闽楠对环境要求较为苛刻,初期生长速度较慢。闽楠属于耐荫植物,强光照的环境不利于生长,适当的遮荫会促进生长,提高植株的株高和冠粗[21–22]。樟、山乌桕早期快速生长为闽楠后期生长提供了合适的生活环境。而速生物种山乌桕经过12 a的生长,迅速发展成为第1优势种,占有更多的环境资源,对樟和闽楠的生长不利。在中后期群落演替中,植被结构趋于复杂,生态位重叠将会更加明显,种内竞争更加激烈[23]。因而建议在闽楠和樟的生长后期,林分应适当间伐,促进闽楠和樟的生长。
在群落演替方面,闽楠-樟人工混交林在育林后未受到严重的破坏,林分苗木自然更新状况良好。群落中乔木的径级结构大致呈倒“J”型,随着径级的增加,乔木数量逐渐减少,大多数乔木植株仍处在幼龄阶段(2.5 cm≤DBH<7.5 cm),平均胸径为6.15 cm。胸径为8~12 cm时,出现1个小高峰,有明显的断层,可能受过一定的干扰和破坏。同时研究表明,群落中所有植株的平均高度≤6 m时表示群落演替处于初级阶段,平均高度13~17 m的为顶级群落[17]。本调查中,个体植株树高≤6 m的共有9 030株,占总数的94.43%;树高≥13 m的植株仅16株,占总数的0.16%,因此推测闽楠-樟树人工混交林群落演替仍处于初级阶段。
群落的物种组成特征及其物种多样性,对研究群落的起源、演替和属性等具有重要的参考意义[20]。本研究基于广州市龙眼洞林场闽楠-樟树人工混交林的调查数据,分析了该区域的物种组成、物种多样性和目标苗木闽楠、樟树的生长情况,结合长期的监测工作,为闽楠、樟树人工林的物种组成和生物多样性提供理论依据,也为能够深入探讨人工林长期生产力、土壤肥力、抗逆性、森林蓄积量、经营周期等生态问题打下良好的基础。在往后的群落演替中,林下物种组成与多样性会发生如何变化还有待研究。
[1] HAN M C, LI Z Y. Ecological benefits evaluation of urban forest and its models [J]. World For Res, 2011, 24(2): 42–46. doi: 10.13348/j. cnki.sjlyyj.2011.02.011.
韩明臣, 李智勇. 城市森林生态效益评价及模型研究现状 [J]. 世界林业研究, 2011, 24(2): 42–46. doi: 10.13348/j.cnki.sjlyyj.2011. 02.011.
[2] ZHAI S L, CHEN B F, LIN N, et al. Structure and species diversity of typical forests in Guangzhou City [J]. Ecol Environ Sci, 2015, 24(10): 1625–1633. doi: 10.16258/j.cnki.1674-5906.2015.10.006.
翟石磊, 陈步峰, 林娜, 等. 广州市典型森林植被的结构及多样性研究 [J]. 生态环境学报, 2015, 24(10): 1625–1633. doi: 10.16258/j. cnki.1674-5906.2015.10.006.
[3] GOU L J, ZHANG W H, LIU G B. Species diversity and interspecific association in development sequence ofplan- tations in Loess Hilly Region [J]. Chin J Appl Ecol, 2007, 18(1): 9–15. doi: 10.16258/j.cnki.1674-5906.2015.10.006.
郭连金, 张文辉, 刘国彬. 黄土丘陵区沙棘人工林发育过程中物种多样性及种间关联变化 [J]. 应用生态学报, 2007, 18(1): 9–15. doi: 10.16258/j.cnki.1674-5906.2015.10.006.
[4] CARDINALE B J, DUFFY J E, GONZALEZ A, et al. Biodiversity loss and its impact on humanity [J]. Nature, 2012, 489(7401): 59–67. doi: 10.1038/nature11148.
[5] CHEN Y, SUN B, LIAO S B, et al. Classification of main phyto- community and biodiversity in Shenzhen [J]. For Res, 2013, 26(5): 636–642. doi: 10.13275/j.cnki.lykxyj.2013.05.016.
陈勇, 孙冰, 廖绍波, 等. 深圳市主要植被群落类型划分及物种多样性研究 [J]. 林业科学研究, 2013, 26(5): 636–642. doi: 10.13275/j. cnki.lykxyj.2013.05.016.
[6] WANG Z X, ZHU J M, WANG J, et al. The response of photosynthetic characters and biomass allocation ofyoung trees to different light regimes [J]. Acta Ecol Sin, 2012, 32(12): 3841–3848. doi: 10. 5846/stxb201112021842.
王振兴, 朱锦懋, 王健, 等. 闽楠幼树光合特性及生物量分配对光环境的响应 [J]. 生态学报, 2012, 32(12): 3841–3848. doi: 10.5846/ stxb201112021842.
[7] WU J Y, HUANG M J, CHEN M G, et al. Study on seedling stage growth and early selection of different provenances of[J]. J CS Univ For Technol, 2015, 35(11): 1–4. doi: 10.14067/j.cnki. 1673-923x.2015.11.001.
吴际友, 黄明军, 陈明皋, 等. 闽楠种源苗期生长差异与早期选择研究 [J]. 中南林业科技大学学报, 2015, 35(11): 1–4. doi: 10.14067/j. cnki.1673-923x.2015.11.001.
[8] SHENG W T. On the maintenance of long-term productivity of plantation in China [J]. For Res, 2018, 31(1): 1–14. doi: 10.13275/j. cnki.lykxyj.2018.01.001.
盛炜彤. 关于我国人工林长期生产力的保持 [J]. 林业科学研究, 2018, 31(1): 1–14. doi: 10.13275/j.cnki.lykxyj.2018.01.001.
[9] CHEN L D. Evaluation of the mixing effect ofand[J]. Anhui Agric Sci Bull, 2013, 19(15): 102– 103. doi: 10.16377/j.cnki.issn1007-7731.2013.15.070.
陈来德. 闽楠与香樟混交效果评价 [J]. 安徽农学通报, 2013, 19 (15): 102–103. doi: 10.16377/j.cnki.issn1007-7731.2013.15.070.
[10] XIAO S D. Comparative experiment on afforestation with,and[J]. Anhui Agric Sci Bull, 2019, 25(1): 118–119. doi: 10.16377/j.cnki.issn1007-7731. 2019.01.045.
肖世栋. 闽楠、木荷与香樟造林效果分析 [J]. 安徽农学通报, 2019, 25(1): 118–119.doi: 10.16377/j.cnki.issn1007-7731.2019.01.045.
[11] LI Z H. Discussion on the construction of wisdom forest farm: Take Longyan cave forest farm in Guangdong Province as an example [J]. For Sci Technol Informat, 2018, 50(4): 57–59. doi: 10.3969/j.issn. 1009-3303.2018.04.020.
李志宏. 智慧林场建设的探讨——以广东省龙眼洞林场为例 [J]. 林业科技情报, 2018, 50(4): 57–59. doi: 10.3969/j.issn.1009-3303. 2018.04.020.
[12] ZHOU B H, YU S J. Characteristics and environmental functions evaluation of thecommunity in Tianzhu Mountain [J]. Geogr Res, 2008, 27(2): 257–265. doi: 10.3321/j.issn:1000-0585.2008. 02.003.
周葆华, 余世金. 天柱山黄山松群落特征及其环境功能评价 [J]. 地理研究, 2008, 27(2): 257–265. doi: 10.3321/j.issn:1000-0585.2008. 02.003.
[13] MA K P. Measuring method of diversity of biological community: I. Measuring method ofdiversity [J]. Biodiv Sci, 1994, 2 (3):162–168.
马克平. 生物群落多样性的测度方法: I.多样性的测度方法 [J].生物多样性, 1994, 2(3): 162–168.
[14] HAN D Y, LI H Y, YANG Y F.-Diversity patterns of plant community in fragmented habitat in a degenerated meadow in Songnen Plain, China [J]. Chin Geogr Sci, 2009, 19(4): 375–381. doi: 10.1007/s11769- 009-0375-7.
[15] CLARKE K R, SOMERFIEL P J, CHAPMAN M G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages [J]. J Exp Mar Biol Ecol, 2006, 330(1): 55–80. doi: 10.1016/j.jembe. 2005.12.017.
[16] DIXON P. VEGAN, a package of R functions for community ecology [J]. J Veg Sci, 2003, 14(6): 927–930. doi: 10.1111/j.1654-1103.2003. tb02228.x.
[17] DU J X, LIU C, YIN C M, et al. Characteristics of the+community in Mt. Mihouling Nature Reserve, Hainan, China [J]. Plant Sci J, 2020, 38(5): 609–617. doi: 10.11913/PSJ.2095-0837.2020.50609.
杜家贤, 刘闯, 殷崇敏, 等. 海南猕猴岭自然保护区海南锥+黄牛木群落特征研究 [J]. 植物科学学报, 2020, 38(5): 609–617. doi: 10. 11913/PSJ.2095-0837.2020.50609.
[18] LIU S S, YE Y C, ZHANG Z S, et al. The species composition of secondary forest at Dalinshan in Dongguan City and its contribution to the species pool [J]. Guangdong For Sci Technol, 2005, 21(4): 18–22. doi: 10.3969/j.issn.1006-4427.2005.04.005.
刘颂颂, 叶永昌, 张柱森, 等. 东莞大岭山村边自然次生林群落物种组成特征及其对区域物种库的贡献 [J]. 广东林业科技, 2005, 21(4): 18–22. doi: 10.3969/j.issn.1006-4427.2005.04.005.
[19] MA L, WU L F, YE H G, et al. Studies on community characteristics ofin Yejingbei, Zengcheng, Guangzhou [J]. J Trop Subtrop Bot, 2012, 20(2): 169–176. doi: 10.3969/j.issn.1005-3395. 2012.02.011.
马磊, 吴林芳, 叶华谷, 等. 广州增城叶迳背乐昌含笑林群落特征研究 [J]. 热带亚热带植物学报, 2012, 20(2): 169–176. doi: 10.3969/ j.issn.1005-3395.2012.02.011.
[20] TETETLA-RANGEL E,DUPUY J M,HERNÁNDEZ-STEFANONI J L, et al. Patterns and correlates of plant diversity differ between common and rare species in a neotropical dry forest [J]. Biodiv Conserv, 2017, 26(7): 1705–1721. doi: 10.1007/s10531-017-1327-7.
[21] RODRÍGUEZ-GARCÍA E, BRAVO F. Plasticity inpopulations of diverse origins: Comparative seedling responses to light and nitrogen availability [J]. For Ecol Manage, 2013, 307: 196–205. doi: 10.1016/j.foreco.2013.06.046.
[22] TANG X L, LIU G Z, JIANG J, et al. Effects of growth irradiance on photosynthesis and photorespiration ofleaves [J]. Funct Plant Biol, 2020, 47(12): 1053–1061. doi: 10.1071/FP20062.
[23] YU F Y, YE X, ZHOU R H, et al. Dynamic changes in plant diversity and soil physical and chemical properties in different habitats in Wenjiang section of Jinma River [J]. J Trop Subtrop Bot, 2021, 29(1): 1–8. doi: 10.11926/jtsb.4237.
余飞燕, 叶鑫, 周润惠, 等. 金马河温江段河岸带不同生境植物物种多样性与土壤理化性质的动态变化 [J]. 热带亚热带植物学报, 2021, 29(1): 1–8. doi: 10.11926/jtsb.4237.
Studies on Species Composition and Diversity ofPlantation in Guangzhou
CHENG Xinxin1,2, HONG Wei3, DENG Shuangwen2, WEI Xueying2, ZHANG Ganrong3, FENG Wei3, GUO Yong3, YE Huagu2, WANG Faguo2
(1. Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China; 2. South China Botanical Garden, Chinese Academy of Sciences, Guangdong Provincial Key Laboratory of Applied Botany,Guangzhou 510650, China; 3. Longyandong Forest Farm of Guangdong Province, Guangzhou 510520, China)
In order to understand the relationship between community structure with species diversity of artificial mixed forest, the community structure of-mixed plantation in Longyandong Forest Farm in Guangzhou was analyzed. The results showed that there were 128 species 9 563 plants in 0.37 hm2quadrat, belonging to 57 families and 99 genera, in which there were 35 tree, 61 shrub, and 98 herb species. In tree layer, the species with importance value at top 5 were(33.86%),(17.24%),(12.96%),(7.92%), and(3.70%), the shrub layer was dominated by Rubiaceae, and did the herb layer by Poaceae. The Shannon-Wiener indexes of tree, shrub and herb layers were 1.230, 1.708, and 1.754, respectively, and the Simpson indexes were 0.635, 0.680, and 0.707, indicating that artificial mixed forest was rich in herbaceous species. The non-metric multidimensional scale (NMDS) analysis showed that the similarity of species composition was not consistent in different plots, and some plots contained occasional species. The diameter class structure of the community showed an inverted “J” type, of which 11.64% were seedlings, and saplings, small trees, strong trees and large trees account for 6.08%, 55.34%, 25.28%, and 0.70%, respectively. Therefore, it was suggested that the-mixed plantation was still in the early successional stage.
-plantation; Species diversity; Community structure; Community succession
10.11926/jtsb.4434
2021-04-25
2021-07-13
广州市野生动植物保护管理办公室项目(SYZFCG-[2017]032); 广东省龙眼洞林场项目(Y934111001)资助
This work was supported by the Project of Guangzhou Municipal Wildlife Conservation and Management Office (Grant No. SYZFCG-[2017]032), and the Project of Longyandong Forest Farm in Guangdong (Grant No. Y934111001).
程欣欣(1997~ ),女,硕士研究生,主要从事植物分类研究。E-mail: 18316234337@163.com
. E-mail: wangfg@scbg.ac.cn
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
建筑与预算(2022年6期)2022-07-08 13:59:18
林业调查规划(2022年3期)2022-06-13 09:31:04
林业勘查设计(2022年1期)2022-02-15 05:35:12
广西林业科学(2021年6期)2022-01-21 07:39:10
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
林业调查规划(2020年3期)2020-06-03 07:02:36
西北植物学报(2015年9期)2015-07-05 07:03:42
建筑与预算(2014年5期)2014-04-11 07:29:28
