
437个小麦品种钾收获指数的变异特征
2022-04-14 01:40刘硕张慧高志源许吉利田汇
中国农业科学 2022年7期
刘硕,张慧,高志源,许吉利,田汇
437个小麦品种钾收获指数的变异特征
刘硕,张慧,高志源,许吉利,田汇
西北农林科技大学资源环境学院/农业农村部西北植物营养与农业环境重点实验室,陕西杨凌712100
【】调查437个小麦品种钾收获指数(potassium harvest index,KHI)的变化范围,分析KHI与产量、籽粒钾含量、籽粒、秸秆、颖壳钾吸收量和钾利用效率之间的关系,同时研究不同株高、育成年代、麦芒对KHI的影响,为钾高效小麦品种的选育提供科学依据。以不同特性的437个小麦品种为试验材料,在河南洛阳和陕西杨凌进行2年(2018—2020)的田间试验,采用增广随机区组试验设计,设置14个区组,每个区组设置31个试验品种,5个对照品种,每个小麦品种种植6行、行长3 m。成熟期取样,每个小区中部选取一行不缺苗的小麦进行收割,脱粒烘干后计算小麦籽粒产量。每个小区中随机选择5个采样点,每个采样点盲抽6个分蘖,贴地剪断,茎穗分别装入网袋,用于测定不同器官钾含量。采用H2SO4-H2O2消煮,火焰光度计法测定籽粒、秸秆和颖壳钾含量,并计算KHI、籽粒、秸秆、颖壳、地上部吸钾量、籽粒钾利用效率(grain K utilization efficiency,GKUE)和地上部钾利用效率(shoot K utilization efficiency,SKUE)等指标。4个环境中(19洛阳、20洛阳、19杨凌和20杨凌),不同小麦品种间KHI均存在显著差异(<0.01),437个小麦品种的KHI变化范围为0.04—0.40,杨凌平均KHI高于洛阳。扬麦18、烟农5158、川麦104、华麦5号、郑麦1860、Dromedaris和太空6号等7个小麦品种具有较高的KHI和产量。KHI与小麦产量在3个环境中呈显著正相关关系(<0.05)。KHI与小麦籽粒钾含量和小麦籽粒吸钾量在3个环境呈显著正相关关系(<0.05)。KHI与秸秆、颖壳及地上部吸钾量存在显著负相关关系(<0.05)。小麦产量、籽粒钾含量、籽粒钾吸收量随着KHI的提高而升高,秸秆、颖壳和地上部吸钾量随着KHI的提高而降低。KHI与GKUE和SKUE呈显著正相关关系(<0.001)。20世纪70年代以前和70—90年代育成的小麦品种的KHI显著低于90年代以后培育的品种(<0.05),1990—2010年与2010年以后育成的小麦品种的KHI无显著差异(>0.05)。随着株高下降,KHI升高。有无麦芒对KHI没有显著影响。不同小麦品种的KHI存在明显的基因型差异;KHI可以作为小麦钾利用效率的评价指标。创新育种方式,提高钾从营养器官到籽粒的转移效率才能进一步提高小麦KHI。育种过程中将小麦株高保持在合适的范围内有利于提高小麦的KHI。
小麦;育种;株高;芒型;钾收获指数
0 引言
【研究意义】小麦是重要的主粮作物,实现小麦绿色生产对于保障中国粮食和生态环境安全具有重要意义[1]。钾是小麦生长发育所必需的营养元素,每生产1 t小麦籽粒需钾20—40 kg[2-3]。钾肥肥效的发挥受多种因素的影响,中国小麦钾肥当季利用率一般低于50%,意味着半数以上的钾肥并没有被作物吸收利用[4-5]。中国已探明钾盐资源储量约为11亿t,总量很少,钾矿资源贫瘠,一半以上的钾盐需要依赖于进口,随着钾肥价格的升高,钾肥施用成本很高[6]。所以提高小麦的钾效率,更有效地利用钾肥和土壤中的钾素是实现农业生产中绿色可持续发展的重要方式之一。作物钾效率的评价可在植物营养学层面、农学层面以及生理层面采用多样化的指标,但这些评价指标主要反映钾的吸收效率和利用效率2个方面。钾吸收效率是指植物吸收的钾占总供钾量的百分比;而钾利用效率是指作物籽粒产量或生物量与地上部总钾吸收量的比值[7-8]。钾收获指数(potassium (K) harvest index,KHI)是指籽粒钾吸收量与地上部总钾吸收量的比值,反映了植物将钾从营养器官向籽粒的转移效率。研究发现,高产小麦具有更高的KHI,可以更多地将钾素转移分配到籽粒[9]。Pettigrew[10]研究发现籽粒蛋白质浓度高的作物具有更高KHI,较高的钾离子水平促进了氨基酸的运输,特别是向发育籽粒的运输。因此,筛选和培育高KHI小麦品种,对于提高小麦对钾肥的利用、降低农业生产成本具有重要意义。【前人研究进展】现有研究表明,作物的KHI存在明显的基因型变异,并且钾肥施用水平对KHI有明显影响。Fageria等[11]研究发现10种基因型菜豆(Linn.)的KHI存在显著差异(<0.01),基因型和钾肥水平互作对菜豆的KHI影响显著,低钾水平下,KHI平均为0.56,在高钾水平下为0.47。吴金涛[12]把钾利用效率和KHI都高的大麦基因型评价为钾高效基因型,发现钾高效基因型大麦的钾素收获指数是低效基因型的2.56倍,KHI高的基因型对钾素有更高的吸收和利用能力。Zhan等[13]研究发现,小麦由低到高的4个产量区间(<4.5、4.5—6、6—7.5和>7.5 t·hm-2)平均KHI分别为23.6%、23.0%、19.8%和20.2%,不同产量的小麦KHI不同。刘璐等[9]对123个小麦品种进行了2年的大田试验,结果发现,当籽粒产量从5 474 kg·hm-2增加到7 891 kg·hm-2时,KHI从20%提升到37%。刁超朋等[14]研究发现小麦在低磷水平下的KHI高于高磷水平,而氮磷收获指数无显著差异。刁超朋等[15]认为在优良小麦品种的选育过程中,应该注重小麦氮磷钾收获指数的提高,使营养物质更多地向籽粒中转移从而提高产量。【本研究切入点】尽管目前已有研究发现不同小麦品种间KHI存在显著差异,但多数研究调查的品种数量较少(一般在100个品种以下),且仅在单一环境中种植,难以了解小麦KHI品种间变异的普遍规律。此外,不同育成年代、不同株高、不同麦芒特性小麦品种KHI的变异特征也尚不明确,而这些信息对于选育钾高效小麦品种具有重要意义。【拟解决的关键问题】本研究以437份不同特性小麦品种为材料,在2个地点进行了2年的田间试验,分析不同小麦品种之间KHI差异以及KHI与小麦产量、不同器官的钾吸收量、钾利用效率评价指标之间的关系,同时还分析不同株高、育成年代、芒型对小麦KHI的影响,以期为选育钾高效和高产小麦品种提供理论依据。
1 材料与方法
1.1 试验地点概况
2018—2019年和2019—2020年在河南省洛阳(112.45°E,34.62°N)、陕西杨凌(108.07°E,34.28°N)进行2年2个地点共4个环境(19洛阳、20洛阳、19杨凌和20杨凌)的田间试验。这两个试验点位于中国小麦主产区,洛阳土壤类型为褐土,杨凌为土。河南洛阳和陕西杨凌均位于温带大陆性季风气候带,2个地点的年均降雨量、气温和无霜期天数均相近[16-17]。2个试验点土壤基本理化性质见表1。
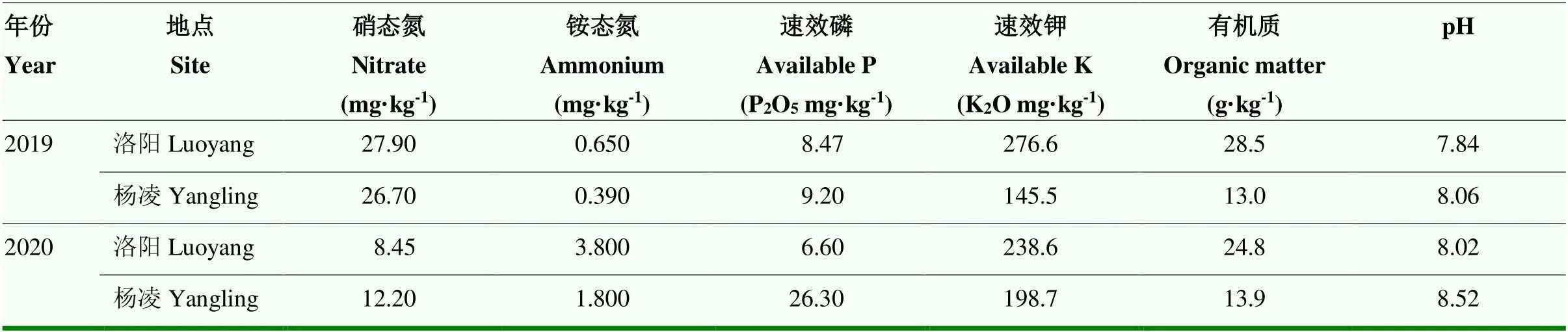
表1 试验点土壤基本理化性质
1.2 试验材料和试验设计
小麦品种取自西北农林科技大学种质资源库,437个小麦品种中,来自亚洲的品种有378个,欧洲15个,美洲22个,大洋洲13个,非洲7个,另2个来源不详。这些品种覆盖了中国主要小麦产区(黄淮麦区208个,长江中下游46个,西南麦区35个,西北麦区80个)。437个品种中,1950年前育成的59个,1951—1970年育成的50个,1971—1990年育成的70个,1991—2010年育成的183个,2010年后育成的32个,43个育成年代不详。有麦芒品种384个,无麦芒品种53个(电子附表1)。
田间试验为增广区组设计,共设14个区组,前13个区组每个区组设置31个试验品种、5个对照小麦品种(周麦18、偃展4110、西农511、济麦22和百农207),第14号区组设置29个试验品种和5个对照品种。利用ACBD-R软件V4.0设计区块组和品种的田间小区排列。每个小麦品种均匀播种6行,每行长3.5 m,行距20 cm,沟深3—5 cm,株距2.5 cm,2个品种之间间隔50 cm。播种前,在每个试验点随机选取10个点,取0—20 cm土层,风干后测定土壤基础指标(pH、有机质、铵态氮、硝态氮、速效磷和速效钾)。施肥、灌溉、杂草和病虫害防治管理均依照当地农民习惯实行(表2)。
1.3 样品的采集及测定
成熟时,每个小区选取中间一行不缺苗的小麦进行收割,装入网袋并编号,风干晾晒以后用脱粒机进行脱粒,烘干后计算小麦籽粒产量。采用盲抽法进行采样,用于测定小麦钾含量。具体实施步骤为:在每个小区中随机选择5个采样点,在每个采样点从小麦基部数出6个分蘖,贴地剪断,茎和穗分别装入不同的网袋中。将小麦植株烘干后,进行人工脱粒,再将籽粒和秸秆在烘箱中烘至恒重,并计算秸秆、颖壳和籽粒的干重。烘干的样品用球磨仪(RETSCHMM400,Germany,氧化锆研磨罐)粉碎,并密封在塑料袋中用于钾含量的测定。用天平称取0.2500 g秸秆和颖壳,籽粒样品称取0.2000 g,放入消煮管中,加入5 ml浓硫酸(95%),先消煮1 h,然后分批加入少量过氧化氢(优级纯),直到液体清亮,后定容并转移到离心管中,用火焰光度计(Model 410,Sherwood Company,England)测定消解液中的钾含量[15]。

表2 2个地点的施肥情况
1.4 数据分析
用R语言进行增广随机区组设计的方差分析,并计算校准值,相关参数计算如下:
籽粒钾吸收量(kg·hm-2)=籽粒产量(kg·hm-2)×籽粒钾含量(g·kg-1)/1000;
秸秆钾吸收量(kg·hm-2)=秸秆干重(kg·hm-2)×秸秆钾含量(g·kg-1)/1000;
颖壳钾吸收量(kg·hm-2)=颖壳干重(kg·hm-2)×颖壳钾含量(g·kg-1)/1000;
地上部钾吸收量(kg·hm-2)=籽粒钾吸收量(kg·hm-2)+秸秆钾吸收量(kg·hm-2)+颖壳钾吸收量(kg·hm-2);
钾收获指数(KHI)=籽粒钾吸收量(kg·hm-2)/地上部钾吸收量(kg·hm-2);
籽粒钾利用效率(grain K utilization efficiency,GKUE)=籽粒产量(kg·hm-2)/地上部钾吸收量(kg·hm-2);
地上部钾利用效率(shoot K utilization efficiency,SKUE)=地上部生物量(kg·hm-2)/地上部钾吸收量(kg·hm-2)。
采用Microsoft Excel 2016进行各项指标的计算、R包augmented RCBD进行增广随机区组设计方差分析;采用IBM SPSS Statistics 26.0进行相关性分析和回归分析,用Excel 2016及Origin 2021进行作图。
2 结果
2.1 小麦KHI、产量和钾效率性状在4个环境的总体分布
4个环境中增广随机区组设计方差分析表明,不同小麦品种间KHI均存在显著差异(<0.01),在19杨凌对照品种之间无显著差异(表3)。
19洛阳、20洛阳、19杨凌和20杨凌4个环境的KHI范围分别为0.02—0.40、0.04—0.25、0.02—0.38和0.05—0.38,最大最小值分别相差0.38、0.21、0.36和0.34。19杨凌和20杨凌KHI之间无显著差异,且平均值高于19洛阳和20洛阳。19洛阳和20洛阳KHI存在显著差异,且19洛阳(0.14)平均值高于20洛阳(0.10)(图1-a)。

表3 不同地点小麦钾收获指数(KHI)增广随机区组设计的方差分析
19洛阳:2019洛阳;20洛阳:2020洛阳;19杨凌:2019杨凌;20杨凌:2020杨凌。:自由度;NS:>0.05;*:<0.05;**:<0.01;***:<0.001。下同
19 Luoyang: 2019 Luoyang; 20 Luoyang: 2020 Luoyang; 19 Yangling: 2019 Yangling; 20 Yangling: 2020 Yangling.: Degree of freedom; NS:>0.05; *:<0.05; **:<0.01; ***:<0.001. The same as below
小麦籽粒产量、籽粒钾含量、籽粒钾吸收量、籽粒钾利用效率(GKUE)、地上部钾利用效率(SKUE)在4个环境中的范围分别为408.01—12 994.07 kg·hm-2、0.92—7.12 g·kg-1、1.42—60.94 kg·hm-2、5.04—97.41 kg·kg-1和44.92—296.30 kg·kg-1(图1)。20洛阳的籽粒产量显著高于其他3个环境,20杨凌的产量最低(图1-b);19杨凌的籽粒钾含量和籽粒吸钾量均显著高于其他3个环境,20杨凌最低(图1-c和图1-d)。20杨凌籽粒钾利用效率和地上部钾利用效率均显著高于其他3个环境,而20洛阳最低(图1-e和图1-f)。将437个品种分别按照KHI和籽粒产量排序,发现扬麦18、烟农5158、川麦104、华麦5号、郑麦1860、Dromedaris和太空6号等7个小麦品种在4个环境中均表现出较高的产量和KHI,平均产量范围为6 631.31 —7 751.69 kg·hm-2,KHI的平均范围为0.17—0.20。
2.2 KHI与产量的关系
4个环境中有3个环境(19洛阳、20洛阳和20杨凌)的小麦KHI与产量的关系符合线性回归关系,且线性方程为最优拟合方程,而19杨凌环境中二者则无显著相关关系(图2)。多数环境中,小麦产量随KHI的提高而增加,回归分析表明小麦KHI每提高0.1,小麦的产量在19洛阳、20洛阳和20杨凌分别提高693.1、648.2和541.3 kg·hm-2,平均提高627.5 kg·hm-2。

图中箱体上下边代表25%和75%位点,上下限代表1.5倍四分位数,箱体中横线代表中位数,方形代表平均值,圆点代表离群值,不同字母表示差异显著(LSD检验,P<0.05)。19洛阳:2019洛阳;20洛阳:2020洛阳;19杨凌:2019杨凌;20杨凌:2020杨凌。下同

*:P<0.05;**:P<0.01;***:P<0.001。下同The same as blow
2.3 KHI与籽粒钾含量及籽粒吸钾量的关系
除19杨凌无显著相关性(>0.05)以外,在其他3个环境(19洛阳、20洛阳和20杨凌)中,KHI与籽粒钾含量之间均存在显著正相关关系(<0.001),且线性方程为最优拟合方程。在3个环境中,小麦KHI每提高0.1,小麦籽粒含钾量分别提高0.41、0.41和0.43 g·kg-1,平均提高0.42 g·kg-1。小麦KHI与籽粒钾吸收量之间,除19杨凌(>0.05)以外,其他3个环境(19洛阳、20洛阳和20杨凌)均存在显著正相关关系(<0.001)。20洛阳和20杨凌最优拟合方程为线性方程,小麦KHI每升高0.1,小麦籽粒钾吸收量升高4.85和3.67 kg·hm-2,而19洛阳KHI与籽粒钾含量的最优拟合方程为对数方程(<0.001),KHI升高时,籽粒吸钾量先快速增加后趋于缓慢增加(图3)。
2.4 KHI与秸秆、颖壳及地上部吸钾量之间的关系
KHI与秸秆钾吸收量在4个环境(19洛阳、20洛阳、19杨凌、20杨凌)中均呈显著负相关关系(<0.001)。19洛阳和20杨凌2个环境最优拟合方程为线性方程,KHI每提高0.1,秸秆钾吸收量分别下降59.82和45.43 kg·hm-2,而20洛阳和19杨凌最优拟合方程分别为二项式和幂函数方程,即在这两个环境中随着KHI的增加而秸秆钾吸收量先快速降低,而后趋于平缓。除19洛阳KHI与颖壳钾吸收量无显著相关性(>0.05)外,在其他3个环境(20洛阳、19杨凌和20杨凌)中二者均呈显著负相关关系(<0.001)。20洛阳环境中KHI与颖壳钾吸收量之间最优拟合方程为线性方程,而19杨凌和20杨凌2个环境中最优拟合方程分别为幂函数方程和对数方程。KHI与地上部钾吸收量在4个环境均呈显著负相关关系(<0.001),地上部钾吸收量随着KHI的增加而降低。19洛阳和20杨凌KHI与地上部钾吸收量呈显著线性关系,KHI每升高0.1,地上部钾吸收量分别降低25.73和44.37 kg·hm-2,平均降低35.05 kg·hm-2。其他2个环境中二者的最优拟合方程分别为对数方程和幂函数方程(图4)。

图3 小麦钾收获指数(KHI)与籽粒钾含量及籽粒吸钾量的关系
2.5 KHI与籽粒钾利用效率(GKUE)和地上部钾利用效率(SKUE)之间的关系
KHI与GKUE在4个环境中均呈显著的正相关关系(<0.001)。回归分析表明,KHI每升高0.1,GKUE分别增加24.3、20.9、22.0和22.5 kg·kg-1,平均增加22.4 kg·kg-1(图5-A)。KHI与SKUE在4个环境中也均呈显著的正相关线性关系(<0.001)。回归分析表明,KHI每提高0.1,SKUE分别增加30.9、26.3、26.4和34.3 kg·kg-1,平均增加29.4 kg·kg-1(图5)。
2.6 不同育成年代小麦品种的KHI差异
在19洛阳中,不同育成年代之间的KHI存在显著差异,KHI随着育成年代的增加有增加的趋势,2011年以后育成的品种KHI显著高于2011年以前(<0.05)。在19杨凌中,1990年是一个分界点,1990年以后育成的品种KHI显著高于1990年以前的品种(<0.05)。20洛阳1991—2010年和2011年以后育成的品种KHI显著高于1990年前的品种,且1971—1990年的品种KHI显著高于1970以前的品种(<0.05)。20杨凌1970年以后的育成品种KHI之间无显著差异,1991—2010年育成的小麦品种KHI显著高于1970年前的品种(<0.05)。4个环境中1991—2010年和2011年以后的小麦品种KHI无显著差异(>0.05)(图6)。

图4 麦钾收获指数(KHI)与秸秆、颖壳及地上部总钾吸收量之间的关系
2.7 不同株高及芒型小麦品种KHI差异
4个环境中小麦的钾收获指数与株高呈显著负相关关系(<0.001),随着小麦株高的增加,小麦的KHI呈下降趋势,且最优拟合方程4个环境均为线性方程。小麦株高每增加20 cm,小麦的KHI分别降低0.016、0.014、0.022和0.008,平均降低0.015。由此可以看出小麦株高越低,钾向籽粒转移的效率越高(图7)。
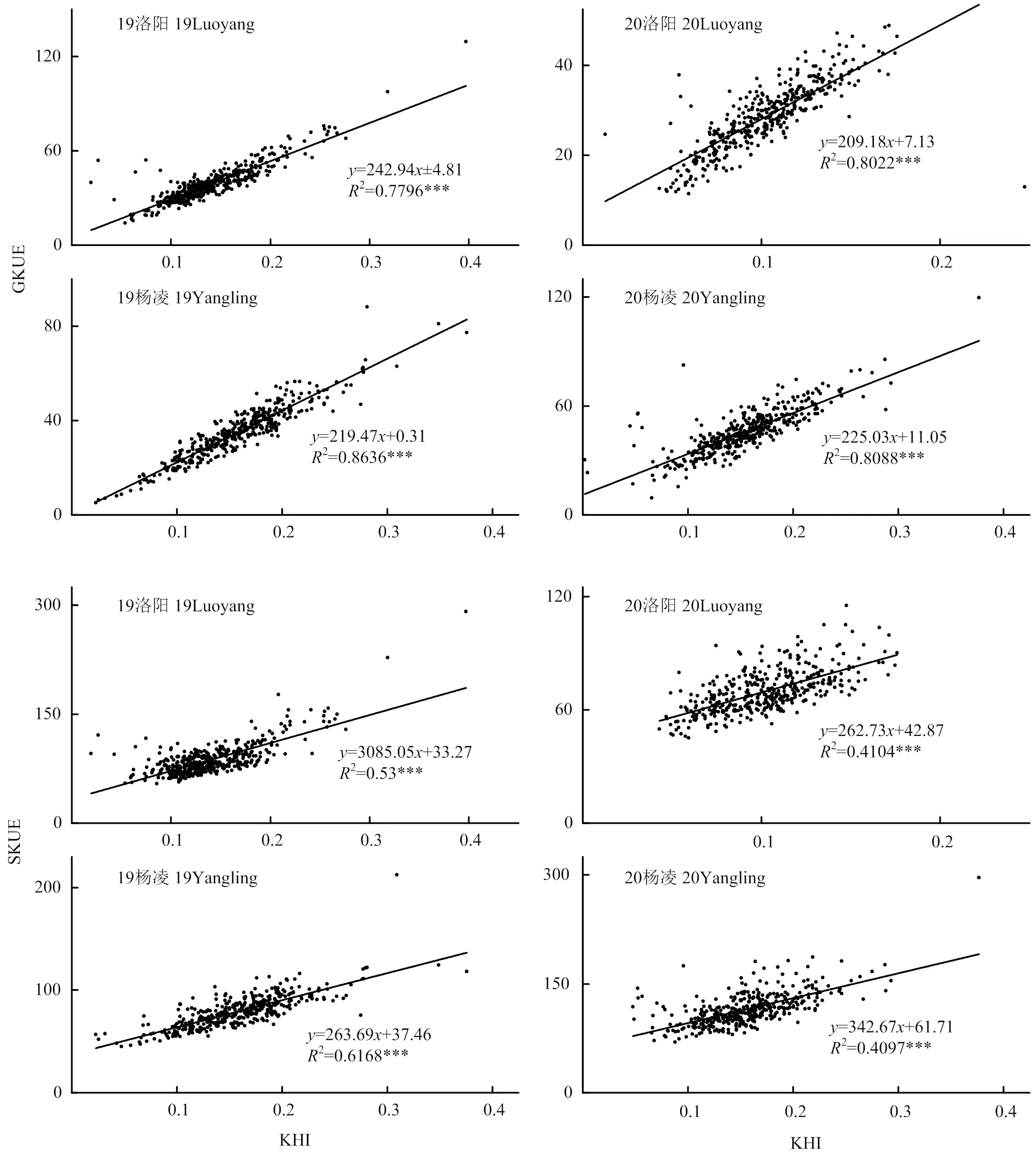
图5 小麦钾收获指数(KHI)与籽粒钾利用效率(GKUE)和地上部钾利用效率(SKUE)之间的关系
综合分析4个环境,仅20洛阳环境中小麦芒型显著影响了KHI(<0.01),有麦芒的高于无麦芒的小麦KHI,而其他3个环境(19洛阳、19杨凌和20杨凌)麦芒有无对KHI无显著影响(>0.05)(图8)。
3 讨论
3.1 小麦KHI在4个环境中的变异特征
本研究发现437个小麦品种的KHI范围为0.04—0.40。汪强[18]利用4个小麦品种设置施钾与不施钾2个不同的处理,发现不施钾处理的KHI为0.19—0.30,施钾处理的KHI为0.16—0.20。化党领[19]利用9个小麦品种,在水培条件下设置4个钾水平,发现小麦KHI的范围为0.28—0.66。赵学强[20]利用9个小麦品种,设置2个钾水平,发现在沙培条件下小麦KHI范围为0.14—0.23,土培条件KHI为0.23—0.43,大田试验为0.30—0.39。张国平[21]利用58个不同基因型的小麦品种在盆栽条件下发现KHI的范围为0.30—0.52。刘璐等[9]利用123个小麦品种进行2年田间试验发现,2014年KHI介于0.15—0.32,2015年介于0.19—0.38。可见,与前人研究相比,本研究主要是扩大了KHI范围的下限,盆栽和水培环境下KHI的上限高于本研究,可能的原因是盆栽试验和田间试验环境差异较大[22]。在本研究中,杨凌的平均KHI高于洛阳(图1)。在总钾供应量(土壤速效钾含量+施钾量)上洛阳2年平均为692.15 kg·hm-2,而杨凌平均为387.27 kg·hm-2,洛阳的钾供应量高于杨凌。土壤钾含量可直接影响作物钾吸收量以及籽粒钾含量[23]。刘荣乐等[24]认为秸秆还田、增施钾肥等均能明显提高作物的钾吸收量,特别是作物秸秆的钾含量,但同时会降低作物籽粒中钾累积量占作物钾库的比例,即提高土壤钾供应量虽然可以提高作物整体的钾吸收量,但会造成KHI的降低。其他研究也发现,随施钾量增加小麦吸钾量随之增加,然而多吸收的钾素大量积累在小麦茎叶器官中[13, 25-27]。尹笑笑[3]研究表明不同土壤肥力虽然对冬小麦植株总钾素积累量影响较大,但对植株钾素的利用能力影响相对较小,很大部分体现在茎秆和籽粒中钾素的差异,土壤肥力较高的地块,能够增加钾素向茎秆中的分配比例,减小向籽粒中的分配比例。所以洛阳试验点较高的供钾水平造成了小麦秸秆钾累积量的增加,最终导致KHI降低。

图6 不同育成年代小麦品种的钾收获指数(KHI)差异

图7 不同株高小麦品种钾收获指数(KHI)差异

图8 不同芒型小麦品种钾收获指数(KHI)差异
3.2 小麦KHI与产量、各器官吸钾量和钾利用效率之间的关系
在本研究中小麦KHI与产量在多数环境中呈显著正相关性关系(<0.05),提升KHI对于小麦的产量有积极的影响(图2)。刘璐等[9]发现小麦KHI随籽粒产量的增加而升高,马小龙等[28]也发现高产小麦KHI高于中低产,这与本研究结果一致。吴金涛[12]也认为选育高KHI的大麦品种是提高大麦产量的一种途径。有研究认为,小麦吸收的钾素可以通过促进输导组织的发育而保障光合产物及营养物质从营养器官向籽粒的顺畅运输,从而有助于籽粒产量的提高[29-30]。然而,本研究发现KHI同样增加0.1,洛阳的小麦产量增加的幅度高于杨凌(图2),这表明KHI与小麦的产量的关系受到土壤供钾能力的影响,即在供钾能力高的土壤上,提高小麦KHI能够更加明显的提高小麦产量。
小麦KHI与籽粒钾含量和籽粒吸钾量在3个环境(19洛阳、19杨凌和20洛阳)均呈显著的正相关关系(图3),说明KHI的提高对于小麦籽粒钾含量和籽粒吸钾量有积极意义。Pettigrew[10]发现籽粒中的K+通过刺激淀粉合成酶的活性,可以促进小麦籽粒灌浆,同时高K+浓度也能促进籽粒中细胞分裂素合成,延缓脱落酸(ABA)产生,有利于提高小麦产量。籽粒中K+对于维持细胞稳态具有重要作用,籽粒中钾素供应充足可以提高蛋白质等有机分子的合成过程中酶的活性[18]。MacLeod[31]发现为了有效利用氮素,大麦籽粒等各组织中的氮钾比例应至少为1﹕1。这都证明了籽粒钾的重要性。Munson[32]研究发现籽粒蛋白质浓度高的作物具有更高KHI,这可能是钾参与氮向籽粒中蛋白质合成部位的运输过程。此外,钾也参与蛋白质合成所需的其他物质(糖、三磷酸腺苷或其他能源)向籽粒的运输[33]。
提高KHI可以促使小麦中钾从秸秆、颖壳等部位转移到籽粒中去,提高了籽粒钾的有效利用率[34]。钾素在秸秆、颖壳、穗和叶中分配比例较高,而钾素在籽粒中分配比例相对来说比较低[3],小麦秸秆表现出钾素“奢侈”吸收现象[35]。所以要提高地上部钾向籽粒中转移,把奢侈吸收的钾素充分利用起来。本研究也发现KHI与颖壳、秸秆、地上部钾的吸收量呈显著负相关关系,并且有的呈二次、对数或者指数关系。KHI在较低的范围时,随着KHI的升高,秸秆和颖壳中钾吸收量降低的较快,向籽粒中转移的较多,而KHI在较高范围时,KHI的提高,秸秆颖壳钾吸收量下降的较慢,这可能是因为在籽粒K+浓度较低时,秸秆颖壳与籽粒之间的K+浓度梯度差低,秸秆颖壳等部位向籽粒中转移顺浓度梯度。随着籽粒钾含量的提高,钾向籽粒中的转移需要的能量更多。籽粒中钾的浓度不宜过高,过高的K+浓度对植物有毒害作用[32],籽粒中的钾需要维持一个稳态,这可能是后边曲线变缓的原因。KHI与地上部总吸钾量呈负相关关系可能是因高KHI的籽粒钾吸收量高,使得地上部吸钾量减少,比值变大。并且我们的数据也发现KHI高的小麦品种秸秆的高度比较低(图7),小麦中的钾素多存在于秸秆中,秸秆干重是地上部的主要组成部分,这可能是KHI高的小麦品种地上部吸钾量反而低的主要原因。本研究发现KHI与GKUE和SKUE呈显著的正相关关系(图5)。GKUE表示吸收单位钾素所能产生的籽粒的量,SKUE表示吸收单位的钾素产生地上部干物质的量[36],本研究结果表明提高KHI有助于小麦高效利用体内的钾素以产出较高的干物质。有研究发现[28, 37],随着产量的提高小麦的GKUE和KHI也随着提高,呈显著的正相关关系。也有研究发现[34, 38-39],在钾胁迫的条件下,KHI与籽粒、地上部和整株钾利用效率均显著增加。因此,在育种过程中选育KHI高的小麦品种意味着其钾利用效率也高,以KHI指标来选育钾高效小麦品种是一种可行方法。
3.3 不同育成年代、不同芒型小麦KHI的变异特征
在本研究中1990年以后育成的小麦品种其KHI显著高于1990年以前的小麦品种,1990年后趋于稳定(图6)。自20世纪40年代以来,品种每一次的更新换代不仅产量提升很大,而且小麦品种的抗病、品质等方面都会得到改善[40]。刘洪升等[41]发现现代品种干物质分配更为合理,利用水分和养分的效率也有很大提升。冯帆[42]研究发现不同年代冬小麦品种光合产物积累和转移特性存在较大的差异,现代品种相对于历史品种有着更高的光合产物转运率。冬小麦品种演变过程中,随着光合性能、产量构成相关性状的改良,源库类型也相对应的发生了改变。Bingham等[43]利用1935—2005年的75个小麦品种,发现小麦氮收获指数(nitrogen harvest index,NHI)也随着育成年代的增加而增加。同样,磷收获指数(phosphorus harvest index,PHI)以每年0.15%的速度增长[44]。Safdar等[45]利用150个1933—2015年的巴基斯坦小麦品种研究发现,绿色革命后的小麦品种钾吸收效率高于绿色革命以前的品种。1990年以后小麦养分利用效率的提高很大程度上可能是由于株高降低所致,株高降低后,叶片光合作用产物向秸秆的分配转移比例也会随之降低,同时小麦的抗倒伏能力得到增强,使其能够更加充分地利用水分和养分等条件提高产量[46]。本研究数据表明,株高每增加20 cm,KHI仅降低0.015,这说明在育种过程中,虽然降低株高对提高KHI有益,但是提高幅度有限(图7)。然而,植株过矮,会造成田间小气候恶劣,光合作用效果减弱,植株体内有机物质积累量减少,进而导致产量降低[47]。曹亚伟等[48]认为株高过高,会降低收获指数,引起植株倒伏,想要获得较高产量,株高最好控制在70—80 cm。所以在选育过程中株高应该保持在一个合适的范围内,不能太高或者太矮。另外本研究还发现,1991—2010年的品种和2011年以后的品种KHI相差不大。因为株高不能无限降低[47],只能通过增加化肥和农药投入或优化农业管理来提高小麦籽粒产量,过量施肥又造成肥力利用率下降[4, 6, 49],并且不能显著提高养分利用效率参数。近十年来,常规育种技术已经遇到了提高小麦养分利用效率的瓶颈,想要提高小麦的养分利用效率需要通过新型的技术手段来提高。目前,已有越来越多关于不同基因型作物钾利用效率差异及其机理的研究,并且已经有多个关于钾效率的QTL被鉴定[1, 36, 45, 50-51],随着基因组学研究的加速,一些植物钾离子通道(shaker、TPK和TPC)和钾离子转运蛋白(KT、HAK、KUP和HKT)也被发现,这些通道和转运蛋白在植物养分吸收和转运中起着重要作用[52-54],分子生物学在营养高效的小麦品种培育过程中具有巨大潜力。
本研究发现有芒小麦和无芒小麦的KHI之间无显著差异(图8)。麦芒在对于氮的转运、同化和积累等方面发挥着重要的作用[55-56],而小麦麦芒在钾吸收和利用效率方面的作用还未见报道。研究表明小麦芒是影响小麦产量的重要组织器官,特别是在干旱胁迫下对于稳定产量有重要意义[57-59];李玲等[60]也发现麦芒对提高小麦光合效率和产量、防虫防鸟、抗逆等方面都有积极意义。芒是最年轻的光合器官,并且在整个籽粒充实过程均具有光合活性,说明芒在植物的养分吸收与利用、转化过程中有重要作用[55]。麦芒不影响小麦KHI的可能原因一是麦芒的生物量较小,麦芒对籽粒钾的贡献率不足以引起小麦KHI的明显提高;二是麦芒在干旱胁迫的环境下才会有更加明显的作用,本研究4个环境下均未遇到明显干旱胁迫的情况。
此外,本研究发现7个小麦品种(扬麦18、烟农5158、川麦104、华麦5号、郑麦1860、Dromedaris、太空6号)同时在多个环境中具有较高的产量和KHI,其中,郑麦1860是2019年育成的国审品种(电子附表1),在黄淮海麦区具有重要推广意义。川麦104、华麦5号和太空6号等品种也是2010年以后育成的较新品种,除了具有高产高KHI的特点,还具有分蘖力强、成穗率较高、品质优良、综合抗病性强等特点[61-63],也适宜作为钾高效小麦品种在中国北方麦区推广种植,或者作为培育钾高效新品种的育种材料。本研究选择了土壤、生产水平和钾素管理差异较大的2个地点4个环境进行大田试验,更加充分地考虑了环境因素对试验结果的影响,有些结论在多个差异较大的环境中表现一致,说明这些结论较少受环境变化的影响,具有较高的重演性。
4 结论
4个生长环境中,437个小麦品种的KHI范围为0.04—0.40,小麦的KHI的变异程度受试验环境的显著影响。小麦KHI与产量之间显著正相关性。随着KHI的升高,小麦籽粒钾含量升高,地上部其他营养器官钾含量降低,KHI提高可以增加籽粒钾的利用效率使其他器官的钾更多地供应给籽粒。KHI与GKUE和SKUE高度正相关,KHI不仅可以反映钾的再利用效率,还可以很好地反映钾的利用效率,是评价钾高效小麦品种的一个重要指标。以产量、品质、抗逆等为育种目标的常规育种技术在提高小麦KHI方面的潜力有限,有针对性地提高小麦钾向籽粒中的转移是未来钾高效小麦品种培育的重要方面。育种过程中将小麦株高保持在合适的范围内有利于提高小麦的KHI;育种过程中是否保留麦芒对提高小麦KHI无显著影响。
[1] ABOU-ELWAFA S F, SHEHZAD T. Genetic diversity, GWAS and prediction for drought and terminal heat stress tolerance in bread wheat (L.). Genetic Resources and Crop Evolution, 2020, 68(2): 711-728.
[2] 赵贤友. 小麦作物氮磷钾肥利用率试验. 现代农业科技, 2018(1): 11-12.
ZHAO X Y. Experiment on utilization efficiency of nitrogen, phosphorus and potassium fertilizer in wheat crops. Modern Agricultural Science and Technology, 2018(1): 11-12. (in Chinese)
[3] 尹笑笑. 不同肥力麦田钾肥运筹对冬小麦产量和钾素利用效率的影响[D]. 泰安: 山东农业大学, 2019.
YIN X X. Effects of potassium fertilizer management on yield and potassium use efficiency of winter wheat under different fertility conditions[D]. Taian: Shandong Agricultural University, 2019. (in Chinese)
[4] 赵玉芬, 尹应武. 我国肥料使用中存在的问题及对策. 科学通报, 2015, 60(36): 3527-3534.
ZHAO Y F, YIN Y W. Problems and countermeasures in the use of fertilizer in China. Chinese Science Bulletin, 2015, 60(36): 3527-3534. (in Chinese)
[5] 曹卫东. 小麦(L.)苗期氮、磷、钾吸收和利用的数量性状位点研究[D]. 北京: 中国农业科学院, 2000.
CAO W D. Study on QTL for absorption and utilization of nitrogen, phosphorous, and potassium in wheat (L.) seedlings[D]. Beijing: Chinese Academy of Agricultural Sciences, 2000. (in Chinese)
[6] 屈小荣, 亓昭英. 我国钾肥行业现状及未来发展趋势分析. 肥料与健康, 2020, 47(5): 7-11.
QU X R, QI Z Y. Analysis of current situation and future development trend of China’s potash fertilizer industry. Fertilizer & Health, 2020, 47(5): 7-11. (in Chinese)
[7] KAMH M, HORST W J, AMER F, MOSTAFA H, MAIER P. Mobilization of soil and fertilizer phosphate by cover crops. Plant and Soil, 1999, 211(1): 19-27.
[8] 雷晶, 郝艳淑, 王晓丽, 王典, 姜存仓. 植物钾效率差异的营养生理及代谢机制研究进展. 中国土壤与肥料, 2014(1): 1-5.
LEI J, HE Y S, WANG X L, WANG D, JIANG C C. Review of nutrient physiology and metabolism in potassium efficiency of plants. Soils and Fertilizers Sciences in China, 2014(1): 1-5. (in Chinese)
[9] 刘璐, 王朝辉, 刁超朋, 王森, 李莎莎. 旱地不同小麦品种产量与干物质及氮磷钾养分需求的关系. 植物营养与肥料学报, 2018, 24(3): 35-44.
LIU L, WANG Z H, DIAO C P, WANG S, LI S S. Grain yields of different wheat cultivars and their relations to dry matter and NPK requirements in dryland. Journal of Plant Nutrition and Fertilizers, 2018, 24(3): 35-44. (in Chinese)
[10] PETTIGREW W T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiologia Plantarum, 2008, 133(4): 670-681.
[11] FAGERIA N K, Barbosa Filho M P, Costa J G C. Potassium-use efficiency in common bean genotypes. Journal of Plant Nutrition, 2001, 24(12): 1937-1945.
[12] 吴金涛. 大麦钾高效基因型筛选及其生理机制研究[D]. 雅安: 四川农业大学, 2010.
WU J T. Study on screening for K-efficient genotype and relevant physiological mechanism[D]. Yaan: Sichuan Agricultural University, 2010. (in Chinese)
[13] ZHAN A, ZOU C, YE Y, LIU Z, CUI Z, CHEN X. Estimating on-farm wheat yield response to potassium and potassium uptake requirement in China. Field Crops Research, 2016, 191: 13-19.
[14] 刁超朋, 黄宁, 李小涵, 王朝辉, 李莎莎, 王森, 刘璐, 惠晓丽, 罗来超. 旱地高产小麦品种籽粒含磷量差异与氮磷钾吸收利用的关系. 植物营养与肥料学报, 2019, 25(3): 351-361.
DIAO C P, HANG N, LI X H, WANG Z H, LI S S, WANG S, LIU L, HUI X L, LUO L C. Difference in grain phosphorus content of high-yielding wheat cultivars and its relation to NPK uptake and utilization in dryland. Journal of Plant Nutrition and Fertilizers, 2019, 25(3): 351-361. (in Chinese)
[15] 刁超朋, 王朝辉, 李莎莎, 刘璐, 王森, 黄宁. 旱地高产小麦品种籽粒氮含量差异与氮磷钾吸收利用的关系. 植物营养与肥料学报, 2018, 24(2): 285-295.
DIAO C P, WANG Z H, LI S S, LIU L, WANG S, HUANG N. Differences in grain nitrogen contents of high-yielding wheat cultivars and relation to NPK uptake and utilization in drylands. Journal of Plant Nutrition and Fertilizers, 2018, 24(2): 285-295. (in Chinese)
[16] 李俊红, 吕军杰, 丁志强, 张洁, 姚宇卿, 蔡典雄, 吴会军, 于新峰. 保护性耕作冬小麦产量及土壤水分变化研究. 土壤通报, 2014, 45(6): 1343-1348.
LI J H, LÜ J J, DING Z Q, ZHANG J, YAO Y Q, CAI D X, WU H J, YU X F. Study of conservation tillage on winter wheat yield and soil moisture. Chinese Journal of Soil Science, 2014, 45(6): 1343-1348. (in Chinese)
[17] 马清霞, 党海燕, 王朝辉, 惠晓丽, 张翔, 张悦悦, 侯赛宾, 黄宁, 罗来超, 张世君. 基于产量和养分含量的旱地小麦施磷量和土壤有效磷优化. 中国农业科学, 2019, 52(1): 73-85.
MA Q X, DANG H Y, WANG Z H, HUI X L, ZHANG X, ZHANG Y Y, HOU S B, HUANG N, LUO L C, ZHANG S J. Optimization of phosphorus rate and soil available phosphorus based on grain yield and nutrient contents in dryland wheat production. Scientia Agricultura Sinica, 2019, 52(1): 73-85. (in Chinese)
[18] 汪强. 不同基因型小麦钾素营养差异性研究[D]. 郑州: 河南农业大学, 2001.
WANG Q. Studies on differences in potassium nutrition of wheat genotypes[D]. Zhengzhou: Henan Agricultural University, 2001. (in Chinese)
[19] 化党领. 不同基因型小麦钾效率差异及生理机制[D]. 郑州: 河南农业大学, 2002.
HUA D L. The differences and physiological mechanisms on potassium efficiency among different wheat genotypes[D]. Zhengzhou: Henan Agricultural University, 2002. (in Chinese)
[20] 赵学强. 不同基因型小麦品种钾效率差异性研究[D] 郑州: 河南农业大学, 2003.
ZHAO X Q. Studies on differences of potassium efficiency of different genotype wheat varieties[D]. Zhengzhou: Henan Agricultural University, 2003. (in Chinese)
[21] 张国平. 小麦钾素利用效率的基因型变异和相关分析. 浙江农业大学学报, 1996, 22(3): 278-283.
ZHANG G P. Variation among wheat genotypes in K utilization and its relationship with some agronomic and K nutrition characters. Journal of Zhejiang Agricultural University, 1996, 22(3): 279- 283. (in Chinese)
[22] 邢宏燕, 王二明, 李滨, 李继云, 李振声. 有效利用土壤磷的小麦种质筛选方法研究. 作物学报, 2000, 26(6): 839-44.
XING H Y, WANG E M, LI B, LI J Y, LI Z S. Studies on wheat germplasm screening method for efficient utilization of soil phosphorus. Acta Agronomica Sinica, 2000, 26(6): 839-844. (in Chinese)
[23] YANG N, WANG Z, GAO Y, ZHAO H, LI K, LI F, MALHI S S. Effects of planting soybean in summer fallow on wheat grain yield, total N and Zn in grain and available N and Zn in soil on the Loess Plateau of China. European Journal of Agronomy, 2014, 58: 63-72.
[24] 刘荣乐, 金继运, 吴荣贵, 梁鸣早. 我国北方土壤作物系统内钾素循环特征及秸秆还田与施钾肥的影响. 植物营养与肥料学报, 2000, 6(2): 123-132.
LIU R L, JIN J Y, WU R G, LIANG M Z. Study on the characteristics of potassium cycling in different soil-crop systems in northern China. Journal of Plant Nutrition and Fertilizers, 2000, 6(2): 123-132. (in Chinese)
[25] 串丽敏. 基于产量反应和农学效率的小麦推荐施肥方法研究[D]. 北京: 中国农业科学院, 2013.
CHUAN L M. Methodology of fertilizer recommendation based on yield response and agronomic efficiency for wheat[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. (in Chinese)
[26] 何刚, 王朝辉, 李富翠, 戴健, 李强, 薛澄, 曹寒冰, 王森, 刘慧. 地表覆盖对旱地小麦氮磷钾需求及生理效率的影响. 中国农业科学, 2016, 49(9): 1657-1671.
HE G, WANG Z H, LI F C, DAI J, LI Q, XUE C, CAO H B, WANG S, LIU H. Nitrogen, phosphorus and potassium requirement and their physiological efficiency for winter wheat affected by soil surface managements in dryland. Scientia Agricultura Sinica, 2016, 49(9): 1657-1671. (in Chinese)
[27] PEITER E, SENBAYRAM M, ZORB C. Potassium in agriculture: status and perspectives. Journal of Plant Physiology, 2014, 171(9): 656-669.
[28] 马小龙, 王朝辉, 曹寒冰, 佘旭, 何红霞, 包明, 宋庆赟, 刘金山. 黄土高原旱地小麦产量差异与产量构成及氮磷钾吸收利用的关系. 植物营养与肥料学报, 2017, 23(5): 1135-1145.
MA X L, WANG Z H, CAO H B, SHE X, HE H X, BAO M, SONG Q Y, LIU J S. Yield variation of winter wheat and its relation to yield components, NPK uptake and utilization in drylands of the loess plateau. Journal of Plant Nutrition and Fertilizer, 2017, 23(5): 1135-1145. (in Chinese)
[29] 熊明彪, 雷孝章, 宋光煜, 曹叔尤. 长期施肥条件下小麦对钾素吸收利用的研究. 麦类作物学报, 2004, 24(1): 51-54.
XIONG M B, LEI X Z, SONG G Y, CAO S Y. Studies on K absorption and utilization in wheat under long-term fertilization. Journal of Triticeae Crops, 2004, 24(1): 51-54. (in Chinese)
[30] 周华, 王月福, 房德强, 赵长星. 氮钾配施对强筋小麦济麦20光合物质积累与分配及产量的影响. 青岛农业大学学报(自然科学版), 2008, 25(3): 193-197.
ZHOU H, WANG Y F, FANG D Q, ZHAO C X. Effects of nitrogen application combined with potassium on photosynthate accumulation, distribution and yield of strong gluten wheat “jimai20”. Journal of Qingdao Agriculture University (Nature science), 2008, 25(3): 193-197. (in Chinese)
[31] MACLEOD L B. Effects of N, P, and K and their interactions on the yield and kernel weight of barley in hydroponic culture. Agronomy Journal, 1969, 61(1): 26-29.
[32] MUNSON R D. Role of potassium in protein metabolism in plants// BLEVINS D G. Potassium in Agriculture. Madison: ASA, CSSA and SSSA, 1985: 413-424.
[33] 毛培培, 赵云云. 植物对钾营养的吸收、运转和胁迫反应的研究进展. 生物学通报, 2008, 43(8): 11-13.
MAO P P, ZHAO Y Y. Research progress on absorption, transport and stress response of plants to potassium nutrition. Bulletin of Biology, 2008, 43(8): 11-13. (in Chinese)
[34] 梁雪. 小麦钾利用相关性状的基因型差异及其QTL分析[D]. 泰安: 山东农业大学, 2012.
LIANG X. Genotypic variation and QTL mapping of potassium use related traits in wheat[D]. Taian: Shandong Agricultural University, 2012. (in Chinese)
[35] TAN D, JIN J, JIANG L, HUANG S, LIU Z. Potassium assessment of grain producing soils in North China. Agriculture, Ecosystems & Environment, 2012, 148: 65-71.
[36] KONG F M, GUO Y, LIANG X, WU C H, WANG Y Y, ZHAO Y, LI S S. Potassium (K) effects and QTL mapping for K efficiency traits at seedling and adult stages in wheat. Plant and Soil, 2013, 373: 877-892.
[37] 黄倩楠, 王朝辉, 黄婷苗, 侯赛宾, 张翔, 马清霞, 张欣欣. 中国主要麦区农户小麦氮磷钾养分需求与产量的关系. 中国农业科学, 2018, 51(14): 2722-2734.
HUANG Q N, WANG Z H, HUANG T M, HOU S B, ZHANG X, MA Q X, ZHANG X X. Relationships of N, P and K requirement to wheat grain yield of farmers in major wheat production regions of China. Scientia Agricultura Sinica, 2018, 51(14): 2722-2734. (in Chinese)
[38] 宫晓平. 小麦钾养分效率及产量相关性状的QTL分析[D]. 泰安: 山东农业大学, 2014.
GONG X P. QTL mapping for K efficiency traits at seedling and yield related traits at harvest stages in wheat[D]. Taian: Shandong Agricultural University, 2014. (in Chinese)
[39] 赵艳艳. 基于SNP标记的小麦钾效率性状的关联分析[D]. 泰安: 山东农业大学, 2016.
ZHAO Y Y. Association analysis for wheat potassium efficiency traits based on molecular marker of SNP[D]. Taian: Shandong Agricultural University, 2016. (in Chinese)
[40] 连慧达. 陕西省不同年代主栽旱地冬小麦品种对氮肥响应的差异研究[D]. 杨凌: 西北农林科技大学, 2020.
LIAN H D. Response of yield and water and nitrogen utilization efficiency to different nitrogen levels in dryland winter wheat (L.) released in different decades of Shaanxi Province of China[D]. Yangling: Northwest A&F University, 2020. (in Chinese)
[41] 刘洪升, 李凤民. 水分胁迫下春小麦根系吸水功能效率的研究. 西北植物学报, 2003, 23(6): 942-948.
LIU H S, LI F M. The study of spring wheat root function efficiency under water stress conditions. Acta Botanica Boreali-Occidentalia Sinica, 2003, 23(6): 942-948. (in Chinese)
[42] 冯帆. 黄淮海麦区不同年代冬小麦品种产量及相关性状演替规律[D]. 杨凌: 西北农林科技大学, 2018.
FENG F. Volution in yield and related traits of winter wheat released in different decades in Huang-Huai-Hai region[D]. Yangling: Northwest A&F University, 2018. (in Chinese)
[43] BINGHAM I J, KARLEY A J, WHITE P J, THOMAS W T B, RUSSELL J R. Analysis of improvements in nitrogen use efficiency associated with 75 years of spring barley breeding. European Journal of Agronomy, 2012, 42: 49-58.
[44] WANG Z, SADRAS V O, HOOGMOED M, YANG X, HUANG F, HAN X, ZHANG S. Shifts in nitrogen and phosphorus uptake and allocation in response to selection for yield in Chinese winter wheat. Crop and Pasture Science, 2017, 68(9): 807-816.
[45] SAFDAR L B, ANDLEEB T, LATIF S, UMER M J, TANG M, LI X, LIU S, QURAISHI U M. Genome-wide association study and QTL meta-analysis identified novel genomic loci controlling potassium use efficiency and agronomic traits in bread wheat. Frontiers in Plant Science, 2020, 11: 70.
[46] 李润芳, 张晓冬, 王栋, 王存娥, 刘世华, 路凌云, 丁汉凤, 李娜娜. 山东省近60年来主推小麦品种主要农艺性状演变规律. 中国农学通报, 2019, 35(7): 20-27.
LI R F, ZHANG X D, WANG D, WANG C E, LIU S H, LU L Y, DING H F, LI N N. Evolution characteristics of major agronomic characters of main planting wheat varieties in Shandong province in 60 years. Chinese Agricultural Science Bulletin, 2019, 35(7): 20-27. (in Chinese)
[47] 刘兆晔, 于经川, 辛庆国. 小麦株高问题的探讨. 山东农业科学, 2014, 46(3): 130-134.
LIU Z Y, YU J C, XIN Q G. Study on plant height of wheat. Shandong Agricultural Sciences, 2014, 46(3): 130-134. (in Chinese)
[48] 曹亚伟, 王健, 刘坤, 唐跃辉, 张怡, 张菊, 徐克东, 李晓丽, 于德水, 齐静, 胡小玉, 原鑫, 张思馨, 高倩倩. 30个冬小麦株高与产量的关系. 浙江农业科学, 2018, 59(7): 1108-1110.
CAO Y W, WANG J, LIU K, TANG Y H, ZHANG Y, ZHANG J, XU K D, LI X L, YU D S, QI J, HU X Y, YUAN X, ZHANG S X, GAO Q Q. Relationship between plant height and yield of 30 winter wheat. Journal of Zhejiang Agricultural Sciences, 2018, 59(7): 1108-1110. (in Chinese)
[49] 刘鑫, 王蕾, 胡飞龙, 马月, 于赐刚, 卢晓强, 刘立, 郑苏平. 《生物多样性公约》下有关农药化肥减量化要求及我国的对策建议. 生态与农村环境学报, 2021(9): 1129-1136.
LIU X, WANG L, HU F L, MA Y, YU C G, LU X Q, LIU L, ZHENG S P. Requirements for reduction of pesticides and fertilizers under the Convention on Biological Diversity and suggestions for China. Journal of Ecology and Rural Environment, 2021(9): 1129-1136. (in Chinese)
[50] SHEN X, YUAN Y, ZHANG H, GUO Y, ZHAO Y, LI S, KONG F. The hot QTL locations for potassium, calcium, and magnesium nutrition and agronomic traits at seedling and maturity stages of wheat under different potassium treatments. Genes (Basel), 2019, 10(8): 607.
[51] 徐易如, 赵艳艳, 孙福来, 郭营, 赵岩, 李斯深, 孔凡美. 小麦成熟期产量及钾效率相关性状的全基因组关联分析. 植物营养与肥料学报, 2020, 26(6): 1081-1090.
XU Y R, ZHAO Y Y, SUN F L, GUO Y, ZHAO Y, LI S S, KONG F M. Genome-wide association analysis for yield and potassium efficiency related traits of wheat at maturity stage. Journal of Plant Nutrition and Fertilizers, 2020, 26(6): 1081-1090. (in Chinese)
[52] LI W, XU G, ALLI A, YU L. Plant HAK/KUP/KT K+transporters: function and regulation. Seminar in Cell & Developmental Biology, 2018, 74: 133-141.
[53] SZE H, CHANROJ S. Plant endomembrane dynamics: Studies of K+/H+antiporters provide insights on the effects of pH and ion homeostasis. Plant Physiology, 2018, 177(3): 875-895.
[54] WANG Y, CHEN Y F, WU W H. Potassium and phosphorus transport and signaling in plants. Journal of Integrative Plant Biology, 2021, 63(1): 34-52.
[55] 谢志新. 氮肥用量对大麦氮代谢和籽粒品质的效应. 浙江农业大学学报, 1989, 15(1): 1-7.
XIE Z X. Effects of fertilizer rates on nitrogen metabolism and grain quality in winter barley. Journal of Zhejiang Agricultural University, 1989, 15(1): 1-7. (in Chinese)
[56] 徐寿军, 包海柱, 张凤英, 刘志萍, 杨恒山, 许如根, 庄恒扬. 施肥水平对冬大麦干物质和氮素积累与转运的影响. 核农学报, 2012, 26(8): 1183-1189, 1203.
XU S J, BAO H Z, ZHANG F Y, LIU Z P, YANG H S, XU R G, ZHUANG H Y. Effects of nitrogen application rates on dry matter, nitrogen accumulation and transformation in barley. Journal of Nuclear Agricultural Science, 2012, 26(8): 1183-1189, 1203. (in Chinese)
[57] 陈培元, 李英. 小麦芒的功能及去芒对籽粒重的影响. 作物学报, 1981(4): 279-282.
CHEN P Y, LI Y. The effect of wheat awns on grain weight and their physiological function. Acta Agronomica Sinica, 1981(4): 279-282. (in Chinese)
[58] 王忠, 高煜珠. 小麦穗的光合特性. 植物学报, 1991, 33(4): 286-291.
WANG Z, GAO Y Z. The photosynthetic characteristics of wheat ear. Acta Botanica Sinica, 1991, 33(4): 286-291. (in Chinese)
[59] 巴青松, 傅兆麟. 小麦穗内芒长与粒重关系的粒位效应. 中国农学通报, 2011, 27(3): 31-34.
BA Q S, FU Z L. Effect of grain position of the relationship between the wheat awn length and grain weight in the spikelet. Chinese Agricultural Science Bulletin, 2011, 27(3): 31-34. (in Chinese)
[60] 李玲, 刘盼, 张蕾, 张浩, 贾继增, 高丽锋. 小麦芒基因定位及其与农艺性状的相关性分析. 植物遗传资源学报, 2021, 22(1): 102-114.
LI L, LIU P, ZHANG L, ZHANG H, JIA J Z, GAO L F. Awn genes mapping and correlation analysis for agronomic traits in wheat. Journal of Plant Genetic Resources, 2021, 22(1): 102-114. (in Chinese)
[61] 代君丽, 崔磊, 刘珂, 宗莹莹, 袁虹霞, 邢小萍, 李洪杰, 李洪连. 小麦品种太空6号对郑州群体的抗性遗传分析. 作物学报, 2013, 39(4): 642-648.
DAI J L, CUI L, LIU K, ZONG Y Y, YUAN H X, XING X P, LI H J, LI H L. Genetic analysis of common wheat cultivar Tai Kong 6 for resistance tozhengzhou population. Acta Agronomica Sinica, 2013, 39(4): 642-648. (in Chinese)
[62] 陆惠芳, 王斌. 小麦新品种华麦5号在平湖地区种植表现及栽培技术. 现代农业科技, 2015(3):47-52.
LU H F, WANG B. Planting performance and cultivation techniques of a new wheat variety Huamai 5 in Pinghu area. Agricultural Science and Technology, 2015(3): 47-52. (in Chinese)
[63] 李式昭, 王琴, 郑建敏, 朱华忠, 李俊, 万洪深, 罗江陶, 刘泽厚, 伍玲. 利用基因芯片分析西南麦区主栽小麦品种川麦104的遗传构成. 麦类作物学报, 2021, 41(6): 665-672.
LI S Z, WANG Q, ZHENG J M, ZHU H Z, LI J, WAN H S, LUO J T, LIU Z H, WU L. Analysis of genetic components in the major wheat cultivar Chuanmai104 in southwest wheat region based on three wheat SNP arrays. Journal of Triticeae Crops, 2021, 41(6): 665-672. (in Chinese)
Genetic Variations of Potassium Harvest Index in 437 Wheat Varieties
LIU Shuo, ZHANG Hui, GAO ZhiYuan, XU JiLi, TIAN Hui
College of Natural Resources and Environment, Northwest A & F University/Key Laboratory of Plant Nutrition and Agro-environment in Northwest China, Ministry of Agriculture and Rural Affairs, Yangling 712100, Shaanxi
【】The present study aimed to investigate the variation of potassium (K) harvest index (KHI) among 437 wheat varieties, analyze the relationships between KHI and yield, grain K content, K absorption of different organs and K utilization efficiency; and clarify the effects of the release year of wheat varieties, plant height and awn types on KHI. The present study provided useful information for breeding wheat cultivars with high yield and K use efficiency. 【】Field experiments were conducted in Luoyang of Henan Province and Yangling of Shaanxi Province during the 2018-2019 and 2019-2020 growth seasons. Four hundred and thirty-seven wheat varieties with different release years, plant heights and awn types were used as materials. An augmented randomized complete block design was applied and 14 blocks were set up, with 31 experimental varieties and 5 control varieties in each block. Each wheat variety was planted with 6 rows and 3 m long. At maturity, a complete row in the middle of each plot was selected for grain harvest, and the grains were oven-dried and weighed. For tissue K concentration measurement, six tillers were blindly selected in five sampling sites in each plot. Grain, straw and glume were separated, oven-dried, and digested using H2SO4-H2O2. K concentrations of different organs were measured with a flame photometer, and parameters including KHI, grain, straw and glume K uptake, grain K utilization efficiency (GKUE) and shoot K utilization efficiency (SKUE) were calculated. 【】There were significant differences in KHI among different wheat varieties (<0.01) under all the four environments (19Luoyang, 20Luoyang, 19Yangling and 20Yangling) and KHI of 437 wheat varieties varied from 0.04 to 0.40. The average KHI of Yangling was higher than that of Luoyang. Seven wheat varieties including Yangmai 18, Yannong 5158, Chuanmai 104, Huamai 5, Zhengmai 1860, Dromedaris and Space 6 had higher KHI and yield. There were significant positive correlations between KHI and wheat yield, grain K concentration and grain K uptake in three environments (<0.05). There were significant negative correlations between KHI and straw K uptake, glume K uptake and shoot total K uptake (<0.05). Grain yield, grain K concentration and uptake increased along with the increase of KHI, however, straw, glume and the shoot total K uptake decreased along with the increase of KHI. There was a significant positive correlation between KHI and GKUE or SKUE (<0.001). The KHI of the wheat varieties released before 1970 and from 1970 to 1990 was significantly lower than that of the varieties released after 1990 (<0.05). The wheat varieties released between 1990 to 2010 had similar KHI with the varieties released after 2010 (>0.05). There was a significant negative correlation between plant height and KHI. There was no significant difference in KHI between the wheat varieties with and without awn. 【】There was a distinct inter-variety variation in wheat KHI. Increasing wheat KHI may positively influence the grain yield of wheat. KHI may be also a good indicator of K utilization efficiency of wheat. To further improve KHI of wheat, novel breeding technologies should be developed to improve the remobilization efficiency of K from vegetative organs to grain. Breeding dwarf or semi-dwarf wheat varieties is beneficial to improve KHI.
wheat; breeding; plant height; awn type; potassium harvest index

10.3864/j.issn.0578-1752.2022.07.003
2021-10-08;
2021-11-26
国家自然科学基金面上项目(31972497)、国家重点研发计划子课题(2017YFD0201702)
刘硕,E-mail:732361104@qq.com。通信作者田汇,E-mail:tianh@nwsuaf.edu.cn
(责任编辑 李莉)
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
今日农业(2022年13期)2022-09-15
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
山东农业科学(2022年6期)2022-07-09
福建农业学报(2022年3期)2022-05-24
节水灌溉(2022年2期)2022-03-04
当代陕西(2020年14期)2021-01-08
当代陕西(2019年12期)2019-07-12
吉林农业(2018年22期)2018-12-23
