
工作记忆的神经振荡调控:基于神经振荡夹带现象*
2022-04-13 07:08王鑫麟邱晓悦翁旭初
心理科学进展 2022年4期
王鑫麟 邱晓悦 翁旭初 杨 平
·研究前沿(Regular Articles)·
工作记忆的神经振荡调控:基于神经振荡夹带现象*
王鑫麟 邱晓悦 翁旭初 杨 平
(华南师范大学脑科学与康复医学研究院; 广东省心理健康与认知科学重点实验室; 心理应用研究中心; 脑认知与教育科学教育部重点实验室, 广州 510631)
工作记忆的神经振荡机制研究是当前记忆领域的研究热点之一。那么, 神经振荡仅仅是工作记忆过程的伴随现象, 还是直接参与并调控了工作记忆的加工过程?已有研究发现, 大脑内部的神经振荡活动在外界节律性刺激的驱动下, 逐步与外界刺激节律相位同步化, 这一现象被称为“神经振荡夹带”。重复经颅磁刺激(repetitive Transcranial Magnetic Stimulation, rTMS)和经颅交流电刺激(transcranial Alternating Current Stimulation, tACS)干预研究基于此现象, 对大脑局部脑区施加节律性磁、电刺激, 进而调控工作记忆过程中特定频段的神经振荡活动、跨频段的神经振荡耦合或跨脑区的神经振荡相位同步, 为神经振荡参与工作记忆加工提供较为直接的因果证据。未来研究需考虑从脑网络的角度出发, 调控多个脑区之间的神经振荡活动, 进一步考察神经振荡对工作记忆的影响。此外, 还需注意探索和优化rTMS/tACS调控工作记忆的刺激方案, 并辅以客观的脑电记录, 提高该类研究的有效性和可重复性, 最终达到提高工作记忆能力的目的。
工作记忆, 神经振荡夹带, 重复经颅磁刺激, 经颅交流电刺激
1 引言
工作记忆(working memory, WM)是大脑高级认知功能的核心组成部分, 它将离散的信息进行暂时的存储和加工, 对指导个体正在(或即将)发生的认知行为具有重要意义(Baddeley, 2010)。工作记忆的神经机制一直以来备受研究者的关注, 其中有两个问题至关重要: 一是工作记忆有哪些脑区参与; 二是这些脑区在何时、以何种神经振荡方式参与工作记忆过程。针对第一个问题, 大量功能磁共振成像(functional magnetic resonance imaging, fMRI)研究已发现个体在进行工作记忆任务时会不同程度地引起背外侧前额叶皮层、顶叶和枕叶等区域的激活, 进而建立起不同脑区激活与工作记忆之间的联系(Ambrose et al., 2016; Christophel et al., 2018; Ester et al., 2009, 2015; Gayet et al., 2017; Xu & Chun, 2006)。同时, 非侵入性脑刺激技术(non-invasive brain stimulation, NIBS): 如经颅磁刺激(transcranial magnetic stimulation, TMS)和经颅直流电刺激(transcranial direct current stimulation, tDCS), 可通过对特定脑区施加抑制性或兴奋性的磁、电刺激, 暂时地干扰局部脑区的神经激活, 进一步推断特定脑区的神经激活和工作记忆任务之间的因果关系(Arciniega et al., 2018; Berryhill et al., 2019; Röhner et al., 2018; Silvanto & Soto, 2012)。然而, 该类研究强调脑区激活对于工作记忆过程的重要性, 很难对工作记忆进程中特定脑区的神经振荡活动以及多脑区之间神经活动同步性的动态变化过程进行检测, 因而无法进一步探究第二个问题。
与此同时, 利用脑磁图(Magnetoencephalographic, MEG)和脑电图(electroencephalographic, EEG)开展的神经电生理学研究具有良好的时间分辨率, 研究者可以通过研究工作记忆中的神经振荡现象来填补这一空白。大脑在执行工作记忆任务时需要大量神经元集群的参与, 神经振荡反映了大脑神经元集群持续性的、有节律的活动, 保证不同神经元集群之间能够进行高效率的信息交流(Bosman et al., 2012; Buzsáki et al., 2012)。神经振荡的频率由慢到快依次被分为delta振荡(0.5~4 Hz)、theta振荡(4~8 Hz)、alpha振荡(8~12 Hz)、beta振荡(13~30 Hz)和gamma振荡。gamma振荡又可分为低频(30~70 Hz)和高频(70~150 Hz) gamma振荡。已有的大量EEG和MEG研究发现, 大脑局部脑区的神经振荡活动、不同脑区间的神经振荡耦合(脑网络)均与工作记忆的加工过程密切相关(Bonnefond & Jensen, 2012; Honkanen et al., 2015; Roux & Uhlhaas, 2014): 如额叶皮层和特定视觉皮层的gamma振荡与工作记忆过程中客体特征信息的维持有关; 左侧颞叶theta-gamma振荡的互频耦合反映了个体记忆视觉信息的精确度。这些研究一定程度上可以阐明不同脑区是何时、以何种神经振荡方式支持工作记忆加工进程, 但同时也引出了一个更为重要的问题: 神经振荡仅仅是工作记忆加工的伴随现象, 还是直接参与并调控了这一过程?
为回答上述这一问题, 研究者通过调控局部脑区或不同脑区之间的神经振荡, 干扰神经网络的局部节点, 继而建立脑区−神经振荡−工作记忆行为之间的因果关系。近些年研究发现, 对大脑神经振荡的调控可以通过 “神经振荡夹带(Entrainment of neural oscillation)”技术实现, 它的原理是外界有节律的刺激施加于大脑, 与大脑特定脑区相应节律的神经振荡相互作用直至同步化, 进而调控大脑内部的神经振荡活动(Thut et al., 2011)。外界有节律的刺激包括: 节律性的知觉刺激, 如有节律的声音和图像; 节律性的非侵入性脑刺激, 如重复经颅磁刺激(repetitive transcranial magnetic stimulation, rTMS)和经颅交流电刺激(transcranial alternating current stimulation, tACS)。其中, 有节律的知觉刺激往往用于驱使大脑感知觉皮层产生特定节律的神经振荡(Garcia et al., 2017; Spaak et al., 2014)。而节律性的NIBS, 如rTMS和tACS两种技术, 则能够针对性地调控感兴趣脑区特定频段的神经振荡活动, 进而推断神经振荡在工作记忆中具体作用的因果关系(Reinhart & Nguyen, 2019; Vosskuhl et al., 2018)。
本文重点回顾了近年来节律性NIBS调控工作记忆神经振荡的相关研究。首先对节律性NIBS调制神经振荡的原理与方法进行介绍。然后, 根据先前研究发现的不同频段神经振荡在工作记忆中的作用, 系统梳理节律性NIBS调控神经振荡、进而影响工作记忆能力的研究进展, 为回答神经振荡是否直接参与工作记忆加工这一问题提供因果证据, 启发未来的研究方向。最后对节律性NIBS干预工作记忆研究的局限性和改进方案进行讨论。
2 对大脑神经振荡的调制: 神经振荡夹带现象
2.1 神经振荡夹带的基本原理
神经振荡夹带的基本原理:外界的节律性刺激可引起大脑相应脑区同步的神经振荡活动, 使大脑内部的神经振荡与外界节律性刺激产生相位锁定, 实现大脑神经振荡与外界节律性刺激的同步化(Thut et al., 2011; Lakatos et al., 2019)。在神经振荡夹带现象发生过程中, 受刺激脑区的神经振荡活动主要有两方面的改变: 一是大脑内源性神经振荡的相位与外界节律性刺激的相位愈发同步, 二是由相位同步导致刺激部位特定频率的神经振荡幅值增加。根据Hanslmayr等人提供的模型(Hanslmayr et al., 2019): 起初, 神经元集群处于基线状态(baseline), 即无规则的放电活动状态; 当节律性刺激施加于大脑后, 神经元集群的活动出现规则化, 其活动频率趋向于与外界节律性刺激同步化, 其活动相位也逐步向外界节律性刺激的频率相位移动, 进入节律同步化形成阶段(Build-up); 最后, 大脑神经振荡与外界节律性刺激两者之间的相位差值为零, 同步化程度最大, 该频率的神经振荡幅值也达到最大(fully entrained)。当节律性刺激移除后, 神经元集群的同步化现象不会马上消失, 而是会持续几个周期, 产生夹带回波(entrainment echo), 随后慢慢地恢复到基线状态。
2.2 神经振荡夹带的实现方法
神经振荡夹带现象可以通过节律性的NIBS实现, 主要包括rTMS和tACS两种技术。TMS利用瞬变的高强度脉冲磁场穿过颅骨, 诱导大脑皮层产生感应电流, 从而刺激神经元活动, 而节律性的rTMS则可影响神经元集群相应节律的神经振荡活动(Vosskuhl et al., 2018)。针对rTMS的神经振荡夹带效应, 具有代表性的一项研究是Thut等人(2011)进行的TMS-EEG实验。他们对被试大脑顶叶施加10 Hz的alpha-rTMS刺激, 并同步记录EEG信号(Thut et al., 2011)。结果发现, 相比于控制条件, alpha-rTMS能够显著诱导被刺激脑区产生同步的alpha节律神经振荡, 且随着大脑内部alpha振荡相位与外界alpha-rTMS相位逐渐同步, 被刺激脑区的alpha振荡幅值也逐渐增加。此外, 该研究通过设置rTMS伪刺激条件(TMS线圈抬起与头皮呈45°角, 避免直接的TMS磁场刺激)作为对照, 控制了节律性rTMS的听觉噪声诱导大脑产生alpha频段神经振荡的可能。该研究为节律性rTMS刺激诱发大脑神经振荡夹带观点提供了有力证据, 也为今后的rTMS实验设计和操作提供了极有价值的参考。
另一种节律性NIBS是tACS, 它通过在被试头部贴附电极, 施加微弱的节律性交流电刺激, 进而诱导神经元集群产生相同节律的神经振荡(Antal & Paulus, 2013)。tACS诱导的神经振荡夹带现象较为直观的证据来源于Helfrich等人(2014)的同步tACS-EEG实验, 研究者利用10 Hz的alpha- tACS刺激被试大脑后顶枕皮层, 结果发现, 相对于伪tACS刺激, 被刺激脑区alpha振荡的相位在alpha-tACS的驱动下逐渐与tACS刺激的相位趋于同步, alpha振荡能量增加(Helfrich et al., 2014)。此后, 关于tACS诱导产生的神经振荡夹带现象, 也陆续在其他认知任务中得到证实(Chander et al., 2016; Voss et al., 2014; Witkowski et al., 2016)。
3 利用NIBS夹带神经振荡调节工作记忆的研究证据
基于“神经振荡夹带”这一现象, 愈来愈多的研究者使用NIBS探究大脑神经振荡与工作记忆作用机制的具体问题。值得注意的是, 这些研究中, 仅有部分研究对节律性NIBS刺激诱导的神经振荡夹带效应进行了检测, 详见表1。下面将依据参与工作记忆神经振荡的频段(由慢到快), 对节律性NIBS夹带神经振荡调节工作记忆的研究证据进行详述。
3.1 夹带theta神经振荡调节工作记忆
3.1.1 额叶和顶叶脑区theta振荡在工作记忆中的作用
额叶theta神经振荡与大脑在工作记忆过程中对认知资源的需求和分配有关。最早由Jensen等人(2002)发现个体在对字母进行有序编码和维持过程中, 额叶的theta振荡显著增强, 且存在记忆负荷效应, 因此研究者认为额叶的theta振荡与记忆资源的需求分配相关(Jensen & Tesche, 2002)。同样的, Griesmayr等人(2010)发现, 相比于简单记忆任务, 当要求被试完成对记忆字母进行重新排序的任务时, 被试在工作记忆维持期间theta振荡在额叶尤其是背外侧前额叶(dorsolateral prefrontal cortex, DLPFC)的活动更为明显(Griesmayr et al., 2010)。这些研究可能表明前额叶并非直接参与记忆信息的存储, 而是更多地参与记忆信息的加工处理和认知资源的分配, 进而完成对记忆信息的维持。Beynel等人(2019)在延迟匹配字母排序任务中, 利用theta-rTMS在记忆编码前或记忆维持期间刺激被试左侧背外侧前额叶皮层(dorsolateral prefrontal cortex, DLPFC), 相比于伪刺激条件, 能够显著提高被试在高难度条件下的工作记忆准确率(Beynel et al., 2019)。此外, 近期一项rTMS研究发现, 在目标物和干扰物同时存在的工作记忆范式中, 利用theta-rTMS (相比于非节律性rTMS)在工作记忆维持期间刺激左侧DLPFC能够显著提高右侧视野目标信息的工作记忆容量, 说明额叶theta神经振荡能够调控大脑将更多的认知资源投入到目标信息的优先处理当中(Riddle et al., 2020)。另外, 近年有研究者还发现额叶theta神经振荡与工作记忆提取阶段新颖刺激的识别相关(Liang et al., 2017), 但对此目前尚未有因果论证的研究, 有待进一步探究。
顶叶theta神经振荡在工作记忆的信息存储和信息操纵中均扮演着重要角色, 具体表现为工作记忆维持期间顶叶theta振荡幅值会随着记忆负荷和记忆任务需求的增加而增加(Palva & Palva, 2011; Sauseng et al., 2010)。如Jaušovec等人(2014)在工作记忆广度测试中利用theta-tACS分别刺激被试的左侧顶叶(P3电极点)、右侧顶叶(P4电极点)和左侧额叶(F3电极点), 结果发现与伪刺激条件相比, 刺激顶叶能够显著提高被试的工作记忆存储能力, 且记忆存储能力的提高在刺激左侧顶叶时最为明显, 然而刺激额叶时并未发现上述效应, 该结果表明顶叶theta振荡在工作记忆存储和更新中发挥了重要作用(Jaušovec et al., 2014; Jaušovec & Jaušovec, 2014)。近来有研究者发现顶叶theta振荡与工作记忆过程中的信息操纵也有关。Albouy等人(2017)发现, 被试在执行对音符进行排序处理的听觉工作记忆任务时, 与仅记忆音符的普通任务相比, 在工作记忆维持期间利用theta-rTMS (相对于非节律rTMS)刺激左侧顶内沟(intraparietal sulcus, IPS)能显著提高被试的记忆准确率, 而且该研究同步记录的EEG信号显示, theta-rTMS刺激不仅增强了IPS的theta振荡能量, 也增强了左侧IPS与右侧DLPFC之间theta振荡的相位同步性, 同时这种增强与听觉工作记忆行为准确率呈正相关(Albouy et al., 2017), 该研究结果明确了IPS的theta振荡与听觉工作记忆信息操纵之间的因果关系。
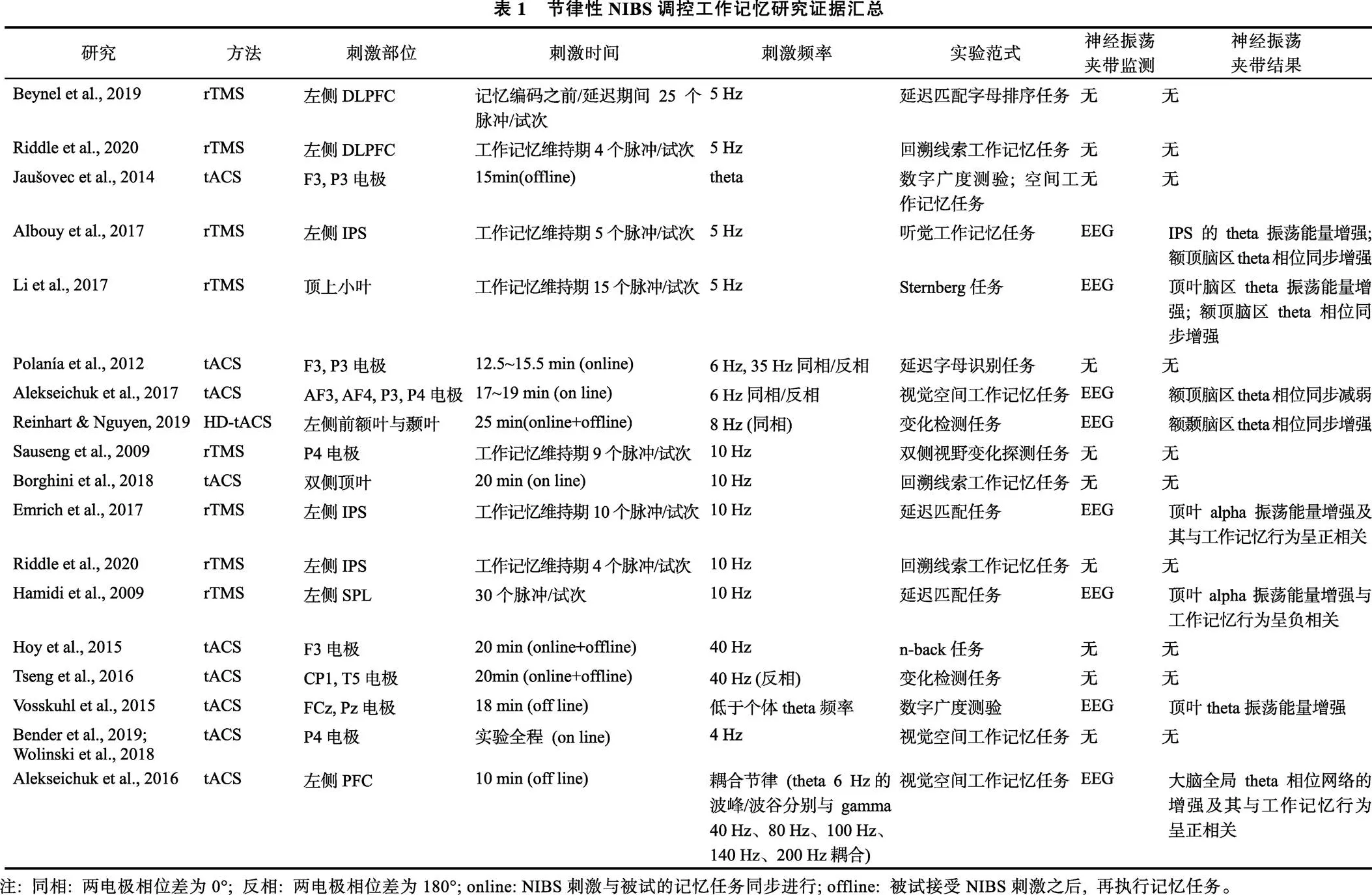
然而, 有研究者认为顶叶IPS脑区的 theta神经振荡并不直接参与工作记忆的信息操纵。Griffiths和Kumar (2017)认为IPS的theta-rTMS刺激增加了DLPFC和IPS之间theta振荡信号的同步性, 促进额顶脑区的信息传递, 加强了额叶对顶叶脑区的调控, 进而才促进了工作记忆信息操纵的行为表现(Albouy et al., 2017; Griffiths & Kumar, 2017)。类似的, Li等人(2017)在Sternberg记忆范式中, 在工作记忆维持期使用theta-rTMS刺激被试顶上小叶(superior parietal lobe, SPL), 并记录EEG信号。结果发现, theta-rTMS刺激同时增强了顶叶脑区theta振荡的能量和额顶脑区之间theta振荡的相位同步性, 使工作记忆行为准确率得到显著提高(Li et al., 2017)。因此, 调节额顶网络中任一脑区的神经振荡, 都可能影响脑网络中其他脑区的神经振荡, 故对该类研究结果的解释需谨慎, 同时也体现了同步EEG记录的重要性, 并启发研究者可从脑网络角度而非局部脑区变化去解释NIBS的干预结果。
3.1.2 额顶、额颞脑区theta振荡同步在工作记忆中的作用
除了单个脑区, 在工作记忆过程中, 不同脑区如何协调和整合记忆信息也受到研究者的广泛关注。以往研究发现, 额顶、颞顶脑区之间的theta振荡相位同步可以整合多个脑区的信息, 实现对工作记忆信息的加工和存储(Daume et al., 2017; Fell & Axmacher, 2011; Fries, 2015; Payne & Kounios, 2009)。例如, 额顶皮层之间theta振荡相位的耦合强度与工作记忆负荷(Payne & Kounios, 2009)、记忆信息操纵的难度(Sauseng et al., 2005)有关, 能够预测工作记忆容量(Kopp et al., 2006)。因此, 除了对单一的额叶或顶叶皮层施加theta节律的NIBS干预外, 大量研究利用tACS调节大脑功能网络中不同脑区之间theta振荡的相位同步性, 观察其对工作记忆能力的影响(Alekseichuk et al., 2017; Polanía et al., 2012; Tseng et al., 2018)。其中, Polanía等人(2012)分别利用相位不同步(相位差为180°)和相位同步(相位差为0°)的theta-tACS (6 Hz)同时刺激被试前额叶(F3电极点)与顶叶脑区(P3电极点), 结果发现, 相比于相位不同步的刺激, 施加相位同步刺激的被试工作记忆行为反应时更短(Polanía et al., 2012)。同样的, Alekseichuk等人(2017)在视觉空间工作记忆任务中, 发现利用相位不同步的theta-tACS刺激额叶和顶叶皮层, 会减弱额叶和顶叶脑区之间的theta相位连接, 导致被试工作记忆能力(击中率−虚报率)下降和反应时增加(Alekseichuk et al., 2017)。
此外, 针对真实世界物体的感知加工主要涉及腹侧视觉通路, 如颞叶皮层。根据工作记忆自上而下的调控机制, 颞叶脑区对于真实物体的加工和表征受到额叶的积极调控, 而这种跨脑区的调控可通过额颞脑区之间theta振荡的相位同步实现(Daume et al., 2017; Fell & Axmacher, 2011)。近来一项针对老年人的MEG研究发现, 与年轻被试相比, 老年被试前额和颞叶脑区之间theta振荡的同步化程度较弱, 研究者认为这可能是导致老年人工作记忆受损的重要原因。在此基础上, 研究者利用高精度的theta-tACS同时刺激被试的额叶和颞叶皮层, 并调控两脑区theta-tACS的相位差, 分别为0°(相位同步)和180°(相位不同步), 结果发现只有0°相位差的theta-tACS能够加强老年被试额颞脑区之间theta振荡的相位同步性, 并提高工作记忆的行为表现(反应时, 准确率) (Reinhart & Nguyen, 2019), 表明额颞网络的theta振荡相位同步在工作记忆跨脑区的信息传递中具有重要意义。
3.2 夹带alpha神经振荡调节工作记忆
顶枕皮层的alpha神经振荡与工作记忆中对干扰信息的抑制能力密切相关(Bonnefond & Jensen, 2012; Jensen et al., 2002; Klimesch et al., 2007; Rihs et al., 2007; van Dijk et al., 2010)。如Sauseng等人(2009)利用双侧视野工作记忆范式, 发现在工作记忆维持期间, 大脑顶枕叶出现明显的alpha振荡偏侧化效应, 与抑制干扰物相关的脑区alpha振荡能量升高, 且随着干扰物数目的增加而增加(Sauseng et al., 2009)。为进一步探究alpha振荡对工作记忆干扰信息的抑制作用, 他们利用alpha-rTMS (10 Hz)在工作记忆维持期刺激被试右侧顶叶(P4电极), 结果发现, alpha-rTMS (相对于15 Hz-rTMS刺激)刺激记忆项呈现视野的同侧脑区时(即抑制干扰物处理), 能够显著提高被试的工作记忆容量, 相反, 当rTMS刺激对侧脑区时(即抑制目标物处理), 则会降低被试的工作记忆容量, 从而验证了工作记忆维持期间顶枕脑区alpha振荡的干扰抑制效应。此后, 在老年人的认知研究中也得到类似的结果(Borghini et al., 2018; Grande et al., 2017), 例如, Borghini等人(2018)根据前人研究推测与年龄相关的认知衰退(如工作记忆准确率下降)可能与抑制干扰信息能力的下降有关, 研究采用有回溯线索和干扰信息的工作记忆任务, 发现在老年被试的右侧顶叶(P4电极)施加alpha-tACS刺激(相对于伪tACS刺激)能够显著提升老年被试的干扰抑制能力, 提高记忆准确率(Borghini et al., 2018)。不过, 上述研究对于刺激位置的选择主要基于脑电电极的P4位点, 其覆盖的具体脑区易受个体差异影响, 因而未能建立具体脑区alpha振荡与工作记忆之间的因果联系。
由于以往研究发现IPS脑区在工作记忆存储中扮演着重要角色(These & Thus, 2004; Todd & Marois, 2005; Xu & Chun, 2006), 且在工作记忆维持期间IPS的alpha振荡活动伴随着负荷效应(Grimault et al., 2009), 较多研究者利用功能磁共振成像精确定位刺激靶点, 探究IPS脑区alpha振荡在干扰抑制中的作用。例如, Emrich等人(2017)在延迟匹配任务中, 利用10 Hz的alpha-rTMS在工作记忆维持期间刺激被试左侧IPS脑区, 发现能够提高被试的工作记忆容量, 不过这种情况仅存在于基线状态下(未接受TMS干预)工作记忆容量较低的被试中(Emrich et al., 2017)。另外, Riddle等人(2020)对左侧IPS施加alpha-rTMS刺激(相对于非节律rTMS)也显著提高了被试的干扰抑制能力, 提升工作记忆容量, 证明了IPS脑区的alpha神经振荡在干扰抑制中的频率特异性和脑区特异性(Riddle et al., 2020)。
3.3 夹带gamma振荡调节工作记忆
gamma振荡(>30 Hz)与视觉工作记忆中客体信息表征的维持有关(Milner, 1974)。Howard等人(2003)通过颅内电信号记录的方式, 发现在Sternberg任务中, 被试枕叶、颞叶和额叶诱发了显著的gamma振荡活动, 且随着记忆负荷的增加而增强(Howard et al., 2003)。来自头皮EEG和MEG的证据也表明, 额叶脑区gamma振荡活动受记忆表征数目和复杂度的影响, 呈现出记忆负荷效应, 与工作记忆容量显著相关(Honkanen et al., 2015; Roux & Uhlhaas, 2014)。据此, 有研究者利用gamma-tACS刺激被试左侧DLPFC (F3电极), 发现在高记忆负荷条件下, 被试的工作记忆表现显著提高(Hoy et al., 2015)。然而, Santarnecchi等人(2015)采用同样的gamma-tACS刺激左侧额叶(F3电极), 却并未发现其对工作记忆表现有显著影响(Santarnecchi et al., 2016)。Pahor等人(2018)采用gamma-tACS刺激被试双侧额叶(F3和F4电极)时, 相较于伪刺激也未发现工作记忆表现的显著变化(Pahor & Jaušovec, 2018)。究其原因, 一方面tACS干预效果容易受任务难度和个体差异的影响, 而Santarnecchi等人没有调控工作记忆任务的难度, 如记忆项数量; 另一方面Pahor等人通过同步EEG信号发现gamma-tACS并未产生神经振荡夹带效应。此外, Tseng等人利用相位差为180°的gamma-tACS同时刺激左侧顶叶和颞叶, 发现可以提高视觉工作记忆的辨别力。该结果在一定程度上表明了在工作记忆过程中颞叶和顶叶皮层之间通过gamma振荡进行信息交流, 脑区之间的gamma振荡存在180°的相位时滞(Tseng et al., 2016), 但这种现象仅在工作记忆能力较差的个体中显著。因此, 在未来研究中, 需注意调整工作记忆的任务难度、被试分组、提高NIBS干预靶点定位的精准性以及结合同步的EEG进行神经振荡监测。
3.4 夹带跨频率振荡耦合调节工作记忆
除了单一频段的神经振荡, 不同频段神经振荡的相互耦合(Cross-frequency coupling, CPC)也在工作记忆中发挥着重要作用(Siebenhühner et al., 2016)。低频率的神经振荡(theta、alpha、beta)常常与更高频率的神经振荡(gamma)同时出现, 其中, 高频的神经振荡反映了神经元集群对外界信息的编码和表征(Jensen et al., 2007), 低频的神经振荡反映了脑区间的信息传递(Daume et al., 2017)。因此, 可以通过调控不同频段神经振荡的耦合验证其在工作记忆中的具体作用。
3.4.1 theta-gamma振荡在工作记忆中的作用
大脑额顶叶皮层的theta-gamma耦合与工作记忆信息的编码和存储有关(Fuentemilla et al., 2010; Rutishauser et al., 2010; Sauseng et al., 2019)。根据theta-gamma耦合模型: 个体gamma振荡的单个子周期代表单个记忆客体的神经表征, 单个theta振荡周期可以内嵌多个gamma振荡子周期。因此, theta振荡单个周期里波峰内嵌的gamma振荡子周期个数越多, 则个体表征的客体数量越多(Lisman & Jensen, 2013)。那么, 若通过神经振荡夹带的方法增加特定脑区theta振荡周期的长度, 则内嵌的gamma振荡子周期个数增加, 进而提高被试的记忆容量。近年的tACS研究结果也验证了以上假设(Bender et al., 2019; Vosskuhl et al., 2015; Wolinski et al., 2018), 研究者在空间工作记忆任务中, 分别采用低频(4 Hz)和高频(7 Hz)的theta-tACS刺激大脑右侧IPS, 结果发现较低频率(4 Hz)的theta-tACS能够显著提升被试的工作记忆容量。不过遗憾的是, 这些研究并未提供同步EEG记录的证据。
此外, theta-gamma耦合模型假设, 当gamma子周期内嵌在theta单个周期的波峰(神经元激活状态)时, 个体对客体信息的表征编码效率最高(Lisman & Jensen, 2013)。Alekseichuk等人(2016)利用tACS-EEG结合的技术, 在空间工作记忆任务中分别利用单节律theta (6 Hz)和theta-gamma复合节律(gamma子周期分别嵌套在theta单个周期波峰、波谷)的tACS刺激左侧前额叶(prefrontal cortex, PFC)。结果发现, gamma子周期嵌套在theta单个周期波峰的复合节律tACS刺激与单节律theta-tACS刺激均能够提高被试在工作记忆任务中的辨别力, 但前者优于后者; 而利用gamma振荡子周期嵌套在theta振荡单个周期波谷的复合节律tACS刺激左侧PFC, 则无此效应(Alekseichuk et al., 2016)。静息态EEG提供的证据也发现, 相比于其它条件, 左侧PFC在接受gamma振荡子周期嵌套在theta振荡单个周期波峰的复合节律tACS刺激后, PFC与其它脑区之间theta振荡的相位同步性显著增强。但由于头皮EEG对于高频gamma活动的检测能力有限, 复合节律的tACS在刺激部位是否成功地夹带诱导出相同模式的神经振荡, 还无法被完全证实, 未来可以考虑使用MEG等其它技术解决此类问题。不过, 上述研究为tACS模拟大脑神经振荡的活动模式提供了新思路, 也为利用复合频率的电刺激研究跨频耦合和认知功能之间的因果关系提供了新途径。
3.4.2 alpha-gamma振荡在工作记忆中的作用
以往的研究证据显示, 大脑后部顶枕脑区的alpha-gamma振荡耦合与外界信息处理门控有关, 它负责抑制与任务无关的干扰物进入视觉编码和记忆存储(Bonnefond & Jensen, 2013, 2015; Jokisch & Jensen, 2007)。alpha-gamma耦合模型认为, 多个gamma振荡子周期内嵌于alpha振荡单个周期的波谷, alpha振荡单个周期缩短, gamma子周期数减少, 工作记忆的表征受到抑制(Bonnefond & Jensen, 2013)。因此, alpha-gamma耦合强度的增加也意味着大脑对干扰物抑制程度的增加。例如, Bonnefond等人(2015)采用了Sternberg工作记忆任务, 并在记忆维持期间设置了强/弱两种干扰物信息, 结果发现在干扰物出现前, 后顶枕部皮层会出现显著的alpha-gamma振荡耦合, 且耦合程度随干扰物强度的增加而增强(Bonnefond & Jensen, 2015)。而当任务相关信息从感知编码进入到记忆系统中, 进行进一步加工时, 顶叶alpha-gamma振荡耦合则会减弱(Tzvi et al., 2018)。先前也有研究者发现, 利用10 Hz的rTMS在记忆维持阶段刺激顶上小叶后, 被刺激脑区的alpha振荡能量和alpha-gamma耦合程度都增强了, 且均与被试工作记忆行为表现的下降显著相关(Hamidi et al., 2009)。上述研究的一个启发在于采用alpha节律的NIBS干预, 不仅能调控alpha频率的神经振荡, 也能够引起alpha-gamma振荡耦合的变化。此外, 有研究通过静息态EEG发现个体alpha振荡的峰值频率(Individual alpha-band frequency, IAF)并不相同(Klimesch et al., 2003)。因此, 针对个体分别施加慢于、等于或快于IAF节律的NIBS干预, 似乎也是一种调控alpha-gamma振荡耦合的尝试。
3.4.3 beta-gamma振荡在工作记忆中的作用
beta-gamma振荡耦合被认为是视觉信息在工作记忆中整合和维持的基础(Bullmore & Sporns, 2009; Siebenhühner et al., 2016)。在前额叶皮层, 研究者采用皮层分层记录方法, 发现深层(layers 5、6)神经元的beta振荡和浅层(layers 2、3)神经元的gamma振荡存在耦合(Bastos et al., 2018; Miller et al., 2018)。具体而言, 在工作记忆编码与维持过程中, 前额叶深层beta振荡与浅层gamma振荡之间的耦合会减弱, 使得gamma振荡能量变强; 而无需加工信息时beta-gamma耦合程度则增强, gamma振荡能量逐渐被抑制(Miller et al., 2018)。同样地, Bastos等人(2018)发现前额叶深层和浅层神经元beta-gamma振荡的耦合强度在工作记忆维持期有所降低, 间接支持了以上观点(Miller et al., 2018)。然而, 现阶段利用节律性NIBS调控单一脑区不同皮层深度的beta-gamma神经振荡的研究还不是很多, 是未来值得探讨的一个研究方向。
针对于浅层皮层, 尚未有研究发现与工作记忆任务相关的局部脑区或跨脑区的beta-gamma振荡耦合现象, 也没有相应的NIBS干预研究, 仍有待研究者进一步探明。若单纯考虑beta振荡, 有研究者发现客体工作记忆中顶叶皮层(Lundqvist et al., 2011)和空间工作记忆中颞叶皮层(Proskovec et al., 2018)的beta振荡幅值会随着记忆负荷的增加而显著下降。此外, 额叶皮层的beta振荡幅值在记忆编码阶段降低(Hanslmayr et al., 2011), 这些均体现了beta振荡的抑制功能。有研究者进一步在记忆编码阶段, 对额下回施加beta-rTMS, 正如我们所预期的, beta-rTMS显著抑制了记忆信息的编码, 同时, 同步测量的EEG信号显示rTMS干预成功诱导了beta振荡的夹带效应, 且beta振荡幅值增强最大的被试记忆表现最差(Hanslmayr et al., 2014)。在其他的一些类似研究中, 研究者还发现在工作记忆维持期间, 视觉皮层和额顶叶皮层会诱发持续性的beta振荡相位耦合(Bullmore & Sporns, 2009; Siebenhühner et al., 2016), 此外, IPS与周围脑区之间的beta相位耦合强度会随着记忆负荷的增加而增加(Palva et al., 2010)。因此, 未来的研究可以尝试调控特定脑区的beta振荡, 探究其对工作记忆行为表现的影响, 并进一步探索beta-gamma耦合是否可以通过单一beta节律的干预进行调控。
4 总结与展望
综上所述, 节律性NIBS干预能够使被刺激脑区产生神经振荡夹带现象, 影响被试记忆表现, 进而为神经振荡活动的确参与调控了工作记忆的加工过程, 提供了较为直接的因果论证。值得注意的是, 相比于脑损伤研究, NIBS所验证的因果关系存在着一定程度的变异性。在脑损伤研究中, 特定脑区的损伤会使得个体的认知功能产生临时或永久的改变(Corrigan & Hammond, 2013), 研究者能够得到特定脑区和特定认知功能之间更为直接且确定的因果关系, 因此该类研究的可重复性高。而在NIBS研究中, 相同的NIBS刺激模式若应用到不同脑区、不同任务中, 对工作记忆行为的影响也会有所不同(Jaušovec & Jaušovec, 2014; Pahor & Jaušovec, 2018)。因此未来研究应着重考量以下几个方面, 以提高该类研究的有效性和可重复性。
4.1 rTMS和tACS的比较
节律性的rTMS和tACS均能诱发被干预脑区相同频段的神经振荡活动, 并通过同步测量EEG信号获得神经振荡夹带的证据(Witkowski et al., 2016)。然而, rTMS和tACS技术各有所长, 在实际应用过程中需要考虑它们的区别: 第一, 相比于tACS, 配有导航系统的rTMS具有较高的空间精度, 主要体现在配有导航系统的rTMS设备可结合个体的大脑结构像实现被干预脑区的精准定位, 而常规的无导航rTMS和tACS常基于EEG电极点位系统进行靶点定位。已有的研究表明, NIBS基于EEG电极点位系统进行靶点定位时, 干预效果不如基于个体结构像的靶点定位(Sack et al., 2008); 第二, 相比于tACS, rTMS具有较高的时间精度。从以往研究中可知, 节律性rTMS常对实验任务中某一特定阶段(如工作记忆维持期500ms时间窗)进行干预, 探究认知过程中某一具体阶段的神经振荡机制, 如顶叶theta振荡对工作记忆信息维持阶段的调控作用; 而tACS则更多对实验前或实验全程进行干预, 探究局部脑区神经振荡与认知功能的关系, 干预时长一般为20分钟(Helfrich et al., 2014; Witkowski et al., 2016); 第三, 相比于rTMS, tACS诱导刺激部位产生的神经振荡夹带效应更加集中在某一预设频率上。rTMS-EEG同步的研究发现, 特定节律的rTMS脉冲刺激不仅能够诱导刺激脑区产生特定频率的神经振荡活动, 还会在其它频段产生脉冲伪迹(Thut et al., 2011); 而tACS由于正弦电流严格限制在一个频率上, 除了预设刺激频率之外不会引起大脑其他频段的神经振荡活动(Herrmann et al., 2013), 因此对于rTMS刺激引起的神经振荡夹带效应需谨慎甄别; 第四, 相比于rTMS, tACS对多个脑区间神经振荡相位同步性的调控更加灵活。节律性rTMS干预可以促进靶点脑区与其他脑区同频率神经振荡的相位同步, 增强两个脑区之间的功能连接(Albouy et al., 2017)。然而, 具备双阳极的tACS可以同时对两个脑区施加特定频段同相位或反相位的节律性电刺激, 以增强或破坏两脑区之间的功能连接(Violante et al., 2017)。此外, tACS还可以灵活施加自定义的theta-gamma复合节律, 从而直接干预特定脑区的互频率耦合强度(Alekseichuk et al., 2016); 最后, 相比于rTMS, tACS没有明显的触觉和噪声感受, 这也是tACS更常用于儿童和老年人群体认知干预研究的原因之一(Antal et al., 2017)。
4.2 神经振荡调控优化策略
实验任务选择。NIBS调节大脑神经振荡的研究通常是在某一具体实验任务中, 这使得研究结果具有较强的任务依赖性, 如节律性材料(图片或声音)的呈现会诱导大脑特定脑区产生相同频段的神经振荡, 因此在选择实验任务时需考虑并排除任务本身对大脑内部神经振荡的影响。值得注意的是, NIBS与实验任务之间的交互作用也会对实验结果产生影响, 已有研究发现rTMS对任务表现的干扰效应在高难度任务中更加显著, 而对简单任务的行为结果影响不大(Barr et al., 2013), 类似的发现也存在于电刺激实验中(Hoy et al., 2015; Violante et al., 2017)。因此, 在设计实验时需要有效控制任务难度、设置对照组等。
关注被试个体差异。首先, 需考虑个体静息态或任务态EEG神经振荡的峰值频率, 如个体alpha振荡的峰值频率一般在8~12 Hz区间内变化。因此, 在实验前若基于个体静息态或任务态EEG神经振荡的峰值频率定义NIBS干预频率, 将有助于提升神经振荡的调控效果(Klimesch et al., 2003)。其次, 在干预前, 靶点脑区的活动状态也会影响NIBS干预效果。如研究者基于fMRI发现DLPFC激活高的被试在rTMS干预后行为改变显著, 而DLPFC激活低的被试则不受rTMS的影响(Sligte et al., 2011)。而且, 任务相关脑区的激活位点也存在个体差异, 因此基于个体fMRI激活的脑区进行靶点定位能有效提升NIBS干预效果(Sack et al., 2008)。此外, 在个体认知能力方面, 有研究发现NIBS干预能够提高低工作记忆容量被试的工作记忆表现, 而对于高记忆容量组被试的影响则不显著(Hsu et al., 2014)。最后, NIBS的神经振荡夹带效果也存在个体差异, 如Hamidi等人(2009)从群体水平并未发现alpha振荡的神经振荡夹带效应, 而从个体水平上发现alpha神经振荡夹带效应越强的被试其工作记忆行为表现越差(Hamidi et al., 2009), 因此对于该类研究结果需谨慎分析和解释。
严格设计刺激方案。NIBS在刺激靶点的选择方式、刺激强度、持续时间、线圈形状、线圈方向等方面的变化均可影响刺激效果。目前, 感兴趣脑区的定位方式共有四种, 按有效性从高到低依次排序为: 基于fMRI个体激活脑区、基于fMRI组分析激活脑区、基于个体结构像MRI和基于EEG电极10-20系统点位(Sack et al., 2008), 在实际应用中, 可依据实验条件选择最优方式。此外, 使用theta脉冲(theta burst stimulation, TBS)刺激被试运动皮层, 若延长刺激的持续时间(较对照组增加一倍), 会使本来对皮层兴奋性具有抑制作用的连续性TBS (cTBS)和对皮层兴奋性具有促进作用的间歇性TBS (iTBS)产生与预期结果相反的皮层兴奋性改变(Gamboa et al., 2010)。也有研究者发现, TMS线圈手柄方向平行于目标脑回时, rTMS刺激效果最好(Thut et al., 2011)。而且, 由于NIBS对大脑皮层兴奋性的影响在个体之间存在差异, 因此在设置刺激模式时需考虑被试特异性, 如在干预前测量被试的运动阈值等。
对照组设置和同步EEG监测。针对NIBS干预的频率特异性, 实验除了设计感兴趣频率的NIBS干预外, 应考虑增加其他频率或非节律的NIBS干预作为对照。此外, 要提高NIBS干预效果, 还需考虑干预前、中和后不同阶段同步EEG信号的采集和监控。实验前的静息态/任务态EEG测量可以提取个体化神经振荡信息, 指导NIBS干预的频率设置; 实验中和实验后EEG测量, 一方面可为节律性NIBS干预提供神经振荡夹带的证据, 另一方面也能探究全脑神经网络的变化, 通过建立NIBS干预引发神经振荡活动或神经振荡网络变化与行为表现变化的相关关系, 为神经振荡参与并调控认知行为提供进一步的因果论证。值得注意的是, 个体的大脑活动始终处于动态变化的状态, 那么如何让NIBS根据大脑状态的实时变化, 相应地调整刺激方案, 对特定神经活动实现准确、有效的调节, 今后还需对NIBS与脑电、脑机接口技术的融合进行优化, 开展相关研究。
最后, 除了提高神经振荡调控的效率, 针对工作记忆问题的研究, 当前研究多以局部脑区为刺激靶点进行研究, 但是对于研究结果的解释不一, 如利用tACS刺激记忆网络中的特定节点可改善工作记忆能力, 这背后的机制是由于该节点的特异性激活, 或是所施加的电流传播到与该节点直接相连的其他脑区, 引起其他脑区激活, 研究者很难给出定论。随着越来越多的理论把记忆涉及的脑区看作是一个网络, 而不是一个个独立的记忆“中心” (Watrous et al., 2013; Winocur & Moscovitch, 2011), 要实现对记忆能力的有效调节, 应该考虑从脑网络的角度促进多个大脑“节点”之间的相互作用, 而不是简单的局部刺激。此外, 针对节律性NIBS如何与大脑内源的神经振荡相互作用产生夹带效应这一问题, 目前研究主要是基于简单的计算模型和对神经元活动的间接测量进行推论, 关于NIBS如何影响大脑神经元活动这一关键问题仍待解决。因此, 若要更好地理解夹带效应对大脑神经元的影响, 未来有必要结合单细胞记录、电刺激或建立神经计算模型等多种方式进行更多的研究。
Albouy, P., Weiss, A., Baillet, S., & Zatorre, R. J. (2017). Selective entrainment of theta oscillations in the dorsal stream causally enhances auditory working memory performance.,(1), 193–206.e5. https://doi.org/10.1016/j.neuron. 2017.03.015
Alekseichuk, I., Pabel, S. C., Antal, A., & Paulus, W. (2017). Intrahemispheric theta rhythm desynchronization impairs working memory.,(2), 147–158. https://doi.org/10.3233/RNN-160714
Alekseichuk, I., Turi, Z., Amador de Lara, G., Antal, A., & Paulus, W. (2016). Spatial working memory in humans depends on theta and high gamma synchronization in the prefrontal cortex.,(12), 1513–1521. https://doi.org/10.1016/j.cub.2016.04.035
Ambrose, J. P., Wijeakumar, S., Buss, A. T., & Spencer, J. P. (2016). Feature-based change detection reveals inconsistent individual differences in visual working memory capacity.,(4), 1–10. https:// doi.org/10.3389/fnsys.2016.00033
Antal, A., Alekseichuk, I., Bikson, M., Brockmöller, J., Brunoni, A. R., Chen, R., … Paulus, W. (2017). Low intensity transcranial electric stimulation: Safety, ethical, legal regulatory and application guidelines.,(9), 1774–1809. https://doi.org/10.1016/j.clinph.2017. 06.001
Antal, A., & Paulus, W. (2013). Transcranial Alternating Current Stimulation (tACS).,(6), 1–4. https://doi.org/10.3389/fnhum.2013.00317
Arciniega, H., Gözenman, F., Jones, K. T., Stephens, J. A., & Berryhill, M. E. (2018). Frontoparietal tDCS benefits visual working memory in older adults with low working memory capacity.,(2), 1–12. https://doi.org/10.3389/fnagi.2018.00057
Baddeley, A. (2010). Working memory.,(4), 136–140. https://doi.org/10.1016/j.cub.2009.12.014
Barr, M. S., Farzan, F., Rajji, T. K., Voineskos, A. N., Blumberger, D. M., Arenovich, T., … Daskalakis, Z. J. (2013). Can repetitive magnetic stimulation improve cognition in schizophrenia? Pilot data from a randomized controlled trial.,(6), 510–517. https://doi.org/ 10.1016/j.biopsych.2012.08.020
Bastos, A. M., Loonis, R., Kornblith, S., Lundqvist, M., & Miller, E. K. (2018). Laminar recordings in frontal cortex suggest distinct layers for maintenance and control of working memory.,(5), 1117–1122. https:// doi.org/10.1073/pnas.1710323115
Bender, M., Romei, V., & Sauseng, P. (2019). Slow theta tACS of the right parietal cortex enhances contralateral visual working memory capacity.,(3), 477–481. https://doi.org/10.1007/s10548-019-00702-2
Berryhill, M., Arciniega, H., Jones, K., Stephens, J., & Gozenman, F. (2019). Who, what, where and how much: tDCS and training effects on working memory.,(2), 460. https://doi.org/10.1016/j.brs.2018. 12.496
Beynel, L., Davis, S., Crowell, C., Hilbig, S., Palmer, H., Brito, A., … Appelbaum, L. (2019). Site-specifics effects of online repetitive Transcranial Magnetic Stimulation (rTMS) on Working Memory (WM).,(2), 564. https://doi.org/10.1016/j.brs.2018.12.867
Bonnefond, M., & Jensen, O. (2012). Alpha oscillations serve to protect working memory maintenance against anticipated distracters.,(20), 1969– 1974. https://doi.org/10.1016/j.cub.2012.08.029
Bonnefond, M., & Jensen, O. (2013). The role of gamma and alpha oscillations for blocking out distraction.,(1), 20–22. https://doi.org/10. 4161/cib.22702
Bonnefond, M., & Jensen, O. (2015). Gamma activity coupled to alpha phase as a mechanism for top-down controlled gating.,(6), 1–11. https://doi.org/ 10.1371/journal.pone.0128667
Borghini, G., Candini, M., Filannino, C., Hussain, M., Walsh, V., Romei, V., … Cappelletti, M. (2018). Alpha oscillationsare causally linked to inhibitory abilities in ageing.,(18), 4418–4429. https://doi.org/10.1523/ JNEUROSCI.1285-17.2018
Bosman, C. A., Schoffelen, J. M., Brunet, N., Oostenveld, R., Bastos, A. M., … Fries, P. (2012). Attentional stimulus selection through selective synchronization between monkey visual areas.,(5), 875–888. https://doi.org/10.1016/ j.neuron.2012.06.037
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph theoretical analysis of structural and functional systems.,(3), 186–198. https://doi.org/10.1038/nrn2575
Buzsáki, G., Anastassiou, C. A., & Koch, C. (2012). The origin of extracellular fields and currents-EEG, ECoG, LFP and spikes.,(6), 407–420. https://doi.org/10.1038/nrn3241
Chander, B. S., Witkowski, M., Braun, C., Robinson, S. E., Born, J., Cohen, L. G., … Soekadar, S. R. (2016). tACS phase locking of frontal midline theta oscillations disrupts working memory performance.,(5), 1–10. https://doi.org/10.3389/fncel. 2016.00120
Christophel, T. B., Iamshchinina, P., Yan, C., Allefeld, C., & Haynes, J. D. (2018). Cortical specialization for attended versus unattended working memory.,(4), 494–496. https://doi.org/10.1038/s41593-018-0094-4
Corrigan, J. D., & Hammond, F. M. (2013). Traumatic brain injury as a chronic health condition.,(6), 1199–1201. https://doi. org/10.1016/j.apmr.2013.01.023
Daume, J., Gruber, T., Engel, A. K., & Friese, U. (2017). Phase- amplitude coupling and long-range phase synchronization reveal frontotemporal interactions during visual working memory.,(2), 313–322. https:// doi.org/10.1523/JNEUROSCI.2130-16.2016
Emrich, S. M., Johnson, J. S., Sutterer, D. W., & Postle, B. R. (2017). Comparing the effects of 10-Hz repetitive TMS on tasks of visual STM and attention.,(2), 286–297. https://doi.org/10.1162/jocn
Ester, E. F., Serences, J. T., & Awh, E. (2009). Spatially global representations in human primary visual cortex during working memory maintenance.,(48), 15258–15265. https://doi.org/10.1523/JNEUROSCI. 4388-09.2009
Ester, E. F., Sprague, T. C., & Serences, J. T. (2015). Parietal and frontal cortex encode stimulus-specific mnemonic representations during visual working memory.,(4), 893–905. https://doi.org/10.1016/j.neuron. 2015.07.013
Fell, J., & Axmacher, N. (2011). The role of phase synchronization in memory processes.,(2), 105–118. https://doi.org/10.1038/nrn2979
Fries, P. (2015). Rhythms for cognition: Communication through coherence.,(1), 220–235. https://doi.org/ 10.1016/j.neuron.2015.09.034
Fuentemilla, L., Penny, W. D., Cashdollar, N., Bunzeck, N., & Düzel, E. (2010). Theta-coupled periodic replay in working memory.,(7), 606–612. https://doi.org/ 10.1016/j.cub.2010.01.057
Gamboa, O. L., Antal, A., Moliadze, V., & Paulus, W. (2010). Simply longer is not better: Reversal of theta burst after-effect with prolonged stimulation.,(2), 181–187. https://doi.org/10.1007/ s00221-010-2293-4
Garcia, M., Miguel, A., & José, A. S. (2017). Binaural auditory beats affect long-term memory.,(1), 1124–1136. https://doi.org/10.1007/s00426- 017-0959-2
Gayet, S., Guggenmos, M., Christophel, T. B., Haynes, J. D., Paffen, C. L. E., van der Stigchel, S., & Sterzer, P. (2017). Visual working memory enhances the neural response to matching visual input.,(28), 6638–6647. https://doi.org/10.1523/JNEUROSCI.3418-16. 2017
Grande, G., Golemme, M., Tatti, E., Chiesa, S., Velzen, J., van, Bernardi Luft., … Cappelletti, M. (2017). P127 A combined EEG and alpha tACS study on visual working memory in healthy ageing.,(3), e77–e78. https://doi.org/10.1016/j.clinph.2016.10.248
Griesmayr, B., Gruber, W. R., Klimesch, W., & Sauseng, P. (2010). Human frontal midline theta and its synchronization to gamma during a verbal delayed match to sample task.,(2), 208–215. https://doi.org/10.1016/j.nlm.2009.09.013
Griffiths, T. D., & Kumar, S. (2017). Driving working memory.,(1), 5–6. https://doi.org/10.1016/j.neuron. 2017.03.031
Grimault, S., Robitaille, N., Grova, C., Lina, J. M., Dubarry, A. S., & Jolicoeur, P. (2009). Oscillatory activity in parietal and dorsolateral prefrontal cortex during retention in visual short-term memory: Additive effects of spatial attention and memory load.,(10), 3378– 3392. https://doi.org/10.1002/hbm.20759
Hamidi, M., Slagter, H. A., Tononi, G., & Postle, B. R. (2009). Repetitive transcranial magnetic stimulation affects behavior by biasing endogenous cortical oscillations.,(6), 1–12. https:// doi.org/10.3389/neuro.07.014.2009
Hanslmayr, S., Axmacher, N., & Inman, C. S. (2019). Modulating human memory via entrainment of brain oscillations.,(7), 485–499. https:// doi.org/10.1016/j.tins.2019.04.004
Hanslmayr, S., Matuschek, J., & Fellner, M. C. (2014). Entrainment of prefrontal beta oscillations induces an endogenous echo and impairs memory formation.,(8), 904–909. https://doi.org/10.1016/j.cub.2014. 03.007
Hanslmayr, S., Volberg, G., Wimber, M., Raabe, M., Greenlee, M. W., & Bäuml, K. H. T. (2011). The relationship between brain oscillations and BOLD signal during memory formation: A combined EEG-fMRI study.,(44), 15674–15680. https://doi.org/10. 1523/JNEUROSCI.3140-11.2011
Helfrich, R. F., Schneider, T. R., Rach, S., Trautmann- Lengsfeld, S. A., Engel, A. K., & Herrmann, C. S. (2014). Entrainment of brain oscillations by transcranial alternating current stimulation.,(3), 333–339. https://doi.org/10.1016/j.cub.2013.12.041
Herrmann, C. S., Rach, S., Neuling, T., & Strüber, D. (2013). Transcranial alternating current stimulation: A review of the underlying mechanisms and modulation of cognitive processes.,(5), 1–13. https://doi.org/10.3389/fnhum.2013.00279
Honkanen, R., Rouhinen, S., Wang, S. H., Palva, J. M., & Palva, S. (2015). Gamma oscillations underlie the maintenance of feature-specific information and the contents of visual working memory.,(10), 3788–3801. https://doi.org/10.1093/cercor/bhu263
Howard, M. W., Rizzuto, D. S., Caplan, J. B., Madsen, J. R., Lisman, J., Aschenbrenner-Scheibe, R., … Kahana, M. J. (2003). Gamma oscillations correlate with working memory load in humans.,(12), 1369–1374. https://doi.org/10.1093/cercor/bhg084
Hoy, K. E., Bailey, N., Arnold, S., Windsor, K., John, J., Daskalakis, Z. J., & Fitzgerald, P. B. (2015). The effect of γ-tACS on working memory performance in healthy controls.,, 51–56. https://doi.org/ 10.1016/j.bandc.2015.11.002
Hsu, T. Y., Tseng, P., Liang, W. K., Cheng, S. K., & Juan, C. H. (2014). Transcranial direct current stimulation over right posterior parietal cortex changes prestimulus alpha oscillation in visual short-term memory task.,, 306–313. https://doi.org/10.1016/j.neuroimage.2014.04.069
Jaušovec, N., & Jaušovec, K. (2014). Increasing working memory capacity with theta transcranial Alternating Current Stimulation (tACS).,(1), 42–47. https://doi.org/10.1016/j.biopsycho.2013.11.006
Jaušovec, N., Jaušovec, K., & Pahor, A. (2014). The influence of theta transcranial Alternating Current Stimulation (tACS) on working memory storage and processing functions.,(1), 1–6. https://doi.org/10.1016/j.actpsy. 2013.11.011
Jensen, O., Gelfand, J., Kounios, J., & Lisman, J. E. (2002). Oscillations in the alpha band (9-12 Hz) increase with memory load during retention in a short-term memory task.,(8), 877–882. http://www.ncbi.nlm.nih. gov/pubmed/12122036
Jensen, O., Kaiser, J., & Lachaux, J. P. (2007). Human gamma-frequency oscillations associated with attention and memory.,(7), 317–324. https://doi.org/10.1016/j.tins.2007.05.001
Jensen, O., & Tesche, C. D. (2002). Short communication frontal theta activity in human increases with memory load in a working memory task.,(8), 1395–1399. https://doi.org/10.1046/ j.1460-9568.2002.01975.x
Jokisch, D., & Jensen, O. (2007). Modulation of gamma and alpha activity during a working memory task engaging the dorsal or ventral stream.,(12), 3244–3251. https://doi.org/10.1523/jneurosci.5399-06.2007
Klimesch, W., Sauseng, P., & Gerloff, C. (2003). Enhancing cognitive performance with repetitive transcranial magnetic stimulation at human individual alpha frequency.,(5), 1129–1133. https://doi.org/ 10.1046/j.1460-9568.2003.02517.x
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG alpha oscillations: The inhibition-timing hypothesis.,(1), 63–88. https://doi.org/10.1016/ j.brainresrev.2006.06.003
Kopp, F., Schröger, E., & Lipka, S. (2006). Synchronized brain activity during rehearsal and short-term memory disruption by irrelevant speech is affected by recall mode.,(2), 188– 203. https://doi.org/10.1016/j.ijpsycho.2005.10.001
Lakatos, P., Gross, J., & Thut, G. (2019). A new unifying account of the roles of neuronal entrainment.,(18), R890–R905. https://doi.org/10.1016/j. cub.2019.07.075
Liang, T., Hu, Z., & Liu, Q. (2017). Frontal theta activity supports detecting mismatched information in visual workingmemory.,(10), 1–8. https://doi.org/ 10.3389/fpsyg.2017.01821
Li, S., Jin, J. N., Wang, X., Qi, H. Z., Liu, Z. P., & Yin, T. (2017). Theta and alpha oscillations during the retention period of working memory by rTMS stimulating the parietal lobe.,(9), 1–12. https://doi.org/10.3389/fnbeh.2017.00170
Lisman, J. E., & Jensen, O. (2013). The theta-gamma neural code.,(6), 1002–1016. https://doi.org/10.1016/j. neuron.2013.03.007
Lundqvist, M., Herman, P., & Lansner, A. (2011). Theta and gamma power increases and alpha/beta power decreases with memory load in an attractor network model.,(10), 3008–3020. https://doi.org/ 10.1162/jocn_a_00029
Miller, E. K., Lundqvist, M., & Bastos, A. M. (2018). Working Memory 2.0.,(2), 463–475. https:// doi.org/10.1016/j.neuron.2018.09.023
Milner, P. M. (1974). A model for visual shape recognition.,(6), 521–535. https://doi.org/ 10.1037/h0037149
Pahor, A., & Jaušovec, N. (2018). The effects of theta and gamma tacs on working memory and electrophysiology.,(1), 1–16. https:// doi.org/ 10.3389/fnhum.2017.00651
Palva, J. M., Palvaa, Monto, S., Kulashekhar, S., & Palva, S. (2010). Neuronal synchrony reveals working memory networks and predicts individual memory capacity.,(16), 7580–7585. https://doi.org/10.1073/pnas.0913113107
Palva, S., & Palva, J. M. (2011). Functional roles of alpha- band phase synchronization in local and large-scale cortical networks.,(9), 1–15. https:// doi.org/10.3389/fpsyg.2011.00204
Payne, L., & Kounios, J. (2009). Coherent oscillatory networks supporting short-term memory retention.,, 126–132. https://doi.org/10.1016/j.brainres.2008.09.095
Polanía, R., Nitsche, M. A., Korman, C., Batsikadze, G., & Paulus, W. (2012). The importance of timing in segregated theta phase-coupling for cognitive performance.,(14), 1314–1318. https://doi.org/10.1016/j.cub. 2012.05.021
Proskovec, A. L., Wiesman, A. I., Heinrichs-Graham, E., & Wilson, T. W. (2018). Beta oscillatory dynamics in the prefrontal and superior temporal cortices predict spatial working memory performance.,(1), 1–13. https://doi.org/10.1038/s41598-018-26863-x
Reinhart, R. M. G., & Nguyen, J. A. (2019). Working memory revived in older adults by synchronizing rhythmic brain circuits.,(5), 820–827. https://doi.org/ 10.1038/s41593-019-0371-x
Riddle, J., Scimeca, J. M., Cellier, D., Dhanani, S., & D’Esposito, M. (2020). Causal evidence for a role of theta and alpha oscillations in the control of working memory.,(9), 1748–1754.e4. https://doi.org/10.1016/j.cub. 2020.02.065
Rihs, T. A., Michel, C. M., & Thut, G. (2007). Mechanisms of selective inhibition in visual spatial attention are indexed by α-band EEG synchronization.,(2), 603–610. https://doi.org/10.1111/ j.1460-9568.2007.05278.x
Röhner, F., Breitling, C., Rufener, K. S., Heinze, H.-J., Hinrichs, H., Krauel, K., & Sweeney-Reed, C. M. (2018). Modulation of working memory using transcranial electrical stimulation: A direct comparison between TACS and TDCS.,(10), 1–10. https:// doi.org/10.3389/fnins.2018.00761
Roux, F., & Uhlhaas, P. J. (2014). Working memory and neural oscillations: Alpha-gamma versus theta-gamma codesfor distinct WM information?,(1), 16–25. https://doi.org/10.1016/j.tics.2013.10.010
Rutishauser, U., Ross, I. B., Mamelak, A. N., & Schuman, E. M. (2010). Human memory strength is predicted by theta-frequency phase-locking of single neurons.,(7290), 903–907. https://doi.org/10.1038/nature08860
Sack, A. T., Kadosh, R. C., Schuhmann, T., Moerel, M., Walsh, V., & Goebel, R. (2008). Optimizing functional accuracy of TMS in cognitive studies : A comparison of methods.(2), 207– 221. https://doi.org/doi:10.1162/jocn.2009.21126
Santarnecchi, E., Muller, T., Rossi, S., Sarkar, A., Polizzotto, N. R., Rossi, A., & Cohen Kadosh, R. (2016). Individual differences and specificity of prefrontal gamma frequency- tACS on fluid intelligence capabilities.,, 33–43. https://doi.org/10.1016/j.cortex.2015.11.003
Sauseng, P., Griesmayr, B., Freunberger, R., & Klimesch, W. (2010). Control mechanisms in working memory: A possible function of EEG theta oscillations.,(7), 1015–1022. https://doi.org/ 10.1016/j.neubiorev.2009.12.006
Sauseng, P., Klimesch, W., Doppelmayr, M., Pecherstorfer, T., Freunberger, R., & Hanslmayr, S. (2005). EEG alpha synchronization and functional coupling during top-down processing in a working memory task.,(2), 148–155. https://doi.org/10.1002/hbm.20150
Sauseng, P., Klimesch, W., Heise, K. F., Gruber, W. R., Holz, E., Karim, A. A., … Hummel, F. C. (2009). Brain oscillatory substrates of visual short-term memory capacity.,(21), 1846–1852. https://doi.org/10.1016/j.cub. 2009.08.062
Sauseng, P., Tschentscher, N., & Biel, A. L. (2019). Be prepared: Tune to FM-theta for cognitive control.,(5), 307–309. https://doi.org/10.1016/ j.tins.2019.02.006
Siebenhühner, F., Wang, S. H., Palva, J. M., & Palva, S. (2016). Cross-frequency synchronization connects networks of fast and slow oscillations during visual working memory maintenance.,(9), 15–30. https://doi.org/ 10.7554/eLife.13451
Silvanto, J., & Soto, D. (2012). Causal evidence for subliminal percept-to-memory interference in early visual cortex.,(1), 840–845. https://doi.org/10.1016/ j.neuroimage.2011.07.062
Sligte, I. G., Wokke, M. E., Tesselaar, J. P., Steven Scholte, H., & Lamme, V. A. F. (2011). Magnetic stimulation of the dorsolateral prefrontal cortex dissociates fragile visual short- term memory from visual working memory.,(6), 1578–1588. https://doi.org/10.1016/j.neuropsychologia. 2010.12.010
Spaak, E., de Lange, F. P., & Jensen, O. (2014). Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception.,(10), 3536–3544. https://doi.org/10.1523/JNEUROSCI. 4385-13.2014
These, M., & Thus, V. (2004). Capacity limit of visual short- term memory in human posterior parietal cortex.,(2003), 751–754. https://doi.org/10.1038/nature02466
Thut, G., Veniero, D., Romei, V., Miniussi, C., Schyns, P., & Gross, J. (2011). Rhythmic TMS causes local entrainment of natural oscillatory signatures.,(14), 1176–1185. https://doi.org/10.1016/j.cub.2011.05.049
Todd, J. J., & Marois, R. (2005). Posterior parietal cortex activity predicts individual differences in visual short- term memory capacity.,(2), 144–155. https://doi.org/10.3758/CABN. 5.2.144
Tseng, P., Chang, Y. T., Chang, C. F., Liang, W. K., & Juan, C. H. (2016). The critical role of phase difference in gamma oscillation within the temporoparietal network for binding visual working memory.,(3), 1–15. https://doi.org/10.1038/srep32138
Tseng, P., Iu, K. C., & Juan, C. H. (2018). The critical role of phase difference in theta oscillation between bilateral parietal cortices for visuospatial working memory.,(1), 1–9. https://doi.org/10.1038/s41598-017-18449-w
Tzvi, E., Bauhaus, L. J., Kessler, T. U., Liebrand, M., Wöstmann, M., & Krämer, U. M. (2018). Alpha-gamma phase amplitude coupling subserves information transfer during perceptual sequence learning.,(2), 107–117. https://doi.org/ 10.1016/j.nlm.2018.02.019
van Dijk, H., van Der Werf, J., Mazaheri, A., Medendorp, W. P., & Jensen, O. (2010). Modulations in oscillatory activity with amplitude asymmetry can produce cognitively relevant event-related responses.,(2), 900–905. https://doi.org/10.1073/pnas.0908821107
Violante, I. R., Li, L. M., Carmichael, D. W., Lorenz, R., Leech, R., Hampshire, A., … Sharp, D. J. (2017). Externally induced frontoparietal synchronization modulates network dynamics and enhances working memory performance.,, 1–22. https://doi.org/10.7554/eLife.22001
Vosskuhl, J., Huster, R. J., & Herrmann, C. S. (2015). Increasein short-term memory capacity induced by down-regulating individual theta frequency via transcranial alternating current stimulation.,(5), 1–10. https://doi.org/10.3389/fnhum.2015.00257
Vosskuhl, J., Strüber, D., & Herrmann, C. S. (2018). Non- invasive brain stimulation: A paradigm shift in understandingbrain oscillations.,(5), 1–19. https://doi.org/10.3389/fnhum.2018.00211
Voss, U., Holzmann, R., Hobson, A., Paulus, W., Koppehele- Gossel, J., Klimke, A., & Nitsche, M. A. (2014). Induction of self awareness in dreams through frontal low current stimulation of gamma activity.,(6), 810–812. https://doi.org/10.1038/nn.3719
Watrous, A. J., Tandon, N., Conner, C. R., Pieters, T., & Ekstrom, A. D. (2013). Frequency-specific network connectivity increases underlie accurate spatiotemporal memory retrieval.,(3), 349–356. https://doi.org/10.1038/ nn.3315
Winocur, G., & Moscovitch, M. (2011). Memory transformation and systems consolidation.,(5), 766–780. https://doi.org/ 10.1017/S1355617711000683
Witkowski, M., Garcia-Cossio, E., Chander, B. S., Braun, C., Birbaumer, N., Robinson, S. E., & Soekadar, S. R. (2016). Mapping entrained brain oscillations during transcranial Alternating Current Stimulation (tACS).,, 89–98. https://doi.org/10.1016/j.neuroimage.2015.10.024
Wolinski, N., Cooper, N. R., Sauseng, P., & Romei, V. (2018). The speed of parietal theta frequency drives visuospatial working memory capacity.,(3), 1–17. https://doi.org/10.1371/journal.pbio.2005348
Xu, Y., & Chun, M. M. (2006). Dissociable neural mechanisms supporting visual short-term memory for objects.,(7080), 91–95. https://doi.org/10.1038/nature04262
Modulating working memory related-oscillation via entrainment of neural oscillation
WANG Xinlin, QIU Xiaoyue, WENG Xuchu, YANG Ping
(Institute for Brain Research and Rehabilitation, South China Normal University; Guangdong Key Laboratory of Mental Health and Cognitive Science; Center for Studies of Psychological Application; Key Laboratory of Brain, Cognition and Education Sciences, Ministry of Education; Guangzhou 510631, China)
The study of the neural oscillation mechanism of working memory (WM) is one of the current research hotspots in the field of memory. However, it is yet not clear whether neural oscillations are only a concomitant phenomenon in the WM process, or if they are directly involved in and can help regulating the WM processing. Several studies have found that brain neural oscillatory activity could be driven by external rhythmic stimuli and gradually synchronizes with the phase of external rhythmic stimuli via a phenomenon known as “neural oscillatory entrainment”. Based on this, a lot of Repetitive Transcranial Magnetic Stimulation (rTMS) and transcranial Alternating Current Stimulation (tACS) intervention studies conducted by modulating neural oscillatory activity in specific frequency bands, phase–amplitude synchronization and phase synchronization between brain regions provided more direct causal evidence of the relationship between neural oscillatory activity and working memory processing. Future studies may look into modulating multiple brain nodes underlying WM by a network approach via rTMS/tACS. Besides, to improve the effectiveness and repeatability of rTMS/tACS intervention, new research in the field should also clarify how to effectively apply rTMS/tACS intervention, supplemented with objective EEG recording to monitor the neural oscillation entrainment.
working memory, oscillation entrainment, rTMS, tACS
2020-12-10
*广东省重点领域研发计划资助(2019B030335001); 国家自然科学基金青年科学基金项目(32100847); 教育部人文社会科学研究一般项目(21YJC190019)。
杨平, E-mail: yangp@m.scnu.edu.cn
B842
猜你喜欢
矿产综合利用(2021年5期)2022-01-17
矿产综合利用(2021年6期)2021-02-21
科学(2020年2期)2020-01-08
中国医学影像技术(2019年3期)2019-03-25
中老年保健(2019年7期)2019-01-13
中国现代医学杂志(2017年3期)2017-03-08
中国生物医学工程学报(2017年6期)2017-02-10
中国CT和MRI杂志(2016年11期)2017-01-18
中国煤炭(2016年9期)2016-06-15
安徽农业科学(2015年10期)2015-04-24
