
番茄苗期节间长度的遗传规律分析
2022-04-11 06:02:46刘文超李毅丰毛秀杰
中国果菜 2022年3期
张 宁,刘文超,李毅丰,王 帅,曹 霞,毛秀杰
(河北科技师范学院园艺科技学院,河北省特色园艺种质挖掘与创新利用重点实验室,河北秦皇岛 066004)
番茄(Solanum lycopersicumL.)为茄科茄属植物,营养丰富,用途广[1],且含有丰富的番茄红素,因此是世界上最重要的蔬菜之一。在生产过程中,受多种环境因素的影响,番茄苗期极易徒长[2],表现为植株细高、节间长[3]。明确番茄节间长度的遗传规律是解决番茄苗期徒长问题的关键[4]。
目前研究者对多种植物的株型和节间长度做了遗传分析研究,如甜瓜[4]、西葫芦[5]等植物的株型、节间长度是受两对主基因的控制,而黄瓜[6]、不结球白菜[7]的节间长度遵循主基因遗传模型。Sun 等[8]研究发现,野生番茄苗期节间长度可能受1 对或2 对主基因控制。虽然前人报道了番茄节间长度差异的现象,但未对番茄节间长度的遗传规律做深入研究。因此,研究短节间番茄育种材料的遗传表达,对培育丰产新品种、抗徒长番茄资源创新具有重要指导作用。为了获得番茄短节间的遗传规律,本文以长节间‘大黄’DH 番茄和短节间‘串黄’CH 番茄为亲本,同时构建6 个世代群体(P1、P2、F1、B1、B2、F2),分别对两个番茄品系苗期、结果期的主要形态进行调查及测定,并利用主基因+多基因进行番茄节间长度遗传规律的分析,为深入解析番茄节间长度的遗传机理提供理论基础。
1 材料与方法
1.1 试验材料
长节间‘大黄’DH番茄和短节间‘串黄’CH 番茄,河北科技师范学院园艺科技学院番茄组提供。
1.2 试验设计
以长节间‘大黄’DH 番茄和短节间‘串黄’CH 番茄为亲本,构建番茄6 个世代群体(P1、P2、F1、F2、B1、B2),其中B1为P1与F1回交,B2为P2与F1回交,F2为F1的自交。供试的P1、P2、F1、F2、B1、B2群体分别为131、107、102、694、156、220 株。2019 年1 月在日光温室中播种。
1.3 形态指标测定方法
两个品系采用随机区组设计,第1 片真叶出现后测定上胚轴长度(即从子叶到第一片真叶的间距),每隔6 d测定1 次,连续测定5 次,每次测量3 株,重复3 次。4~5片真叶展开时测定6 个世代的节间长度。植株高度为植株在自然状态下,其最高点至地面的垂直距离[9]。根据公式(1)计算节间长度。

1.4 统计与评价方法
通过极大似然法和IECM算法对番茄节间长度的遗传参数做出估计,然后对番茄节间长度的遗传模型进行测验:(均匀性测验),nW2(Smirnov 检验)和Dn(Kolmogorov 检验),从而选择番茄节间长度的最优遗传模型,并估计主基因和多基因效应值、方差等遗传参数[10]。
用SPSS 21.0、SEA-G6、Excel 2010 和南京农业大学遗传模型分析软件进行数据分析[11]。试验共进行3 次重复,根据Duncan’s 最小显著差异测验(P<0.05),使用方差分析对每次处理的结果与平均值之间的差异进行显著性分析。
2 结果与分析
2.1 番茄上胚轴长度的生长变化
由图1(见下页)可知,前6 d,番茄DH 上胚轴生长速度快于CH,第6 天以后2 个番茄品系的上胚轴长度增长速度均比较平缓。在第18 天2 个品系差异最大,第18 天以后上胚轴长度变化幅度趋于平缓。在0~24 d,DH 的上胚轴长度显著高于CH。
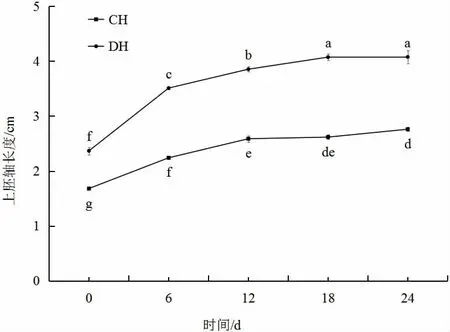
图1 DH、CH 番茄上胚轴长度的变化Fig.1 The change of hypocotyl length in DH and CH tomato
通过对苗期的两个番茄上胚轴长度的t 测验,由表1(见下页)可知,在苗期,DH 上胚轴长度的均值为4.068 mm,CH 上胚轴长度的均值为2.718 mm,t值为19.28(P=0<0.01);DH 的上胚轴长度极显著高于CH。
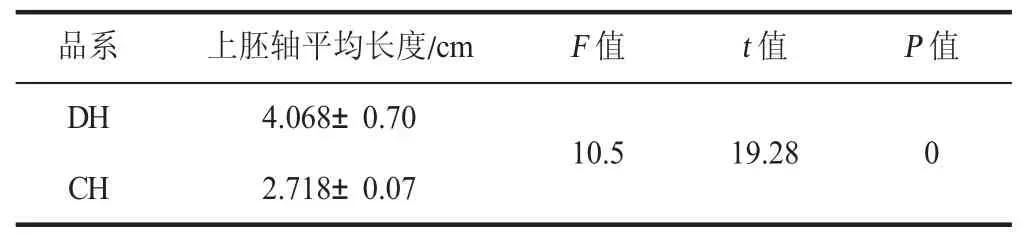
表1 两个番茄品系苗期上胚轴长度的比较Table 1 Comparison of the epicotyl length between two tomato strains at seedling stage
2.2 番茄苗期节间长度的遗传分析
2.2.1 节间长度的变异分析
对苗期的两个番茄品系的平均节间长度进行t 测验分析,结果见表2。由表2 可知,DH 的平均节间长度为2.62 cm,CH 的平均节间长度为1.86 cm,t值为12.15(P=0.01),DH 的平均节间长度显著高于番茄品系CH 的平均节间长度。

表2 两个番茄品系苗期平均节间长度的比较Table 2 Comparison of average internode length of two tomato strains at seedling stage
由表3 可知,P1、P2、F1、F2、B1、B2的变异系数分别为2.14%、2.52%、2.54%、0.88%、1.74%、1.59%。6 个世代遗传群体,变异系数比较相近,分离群体B1、B2、F2的节间长度呈连续分布(表3),表现偏正态分布(图2),因此,适合进行遗传模型分析。
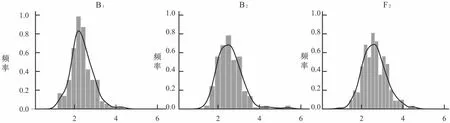
图2 B1、B2、F2 苗期节间长度的频率分布Fig.2 Frequency distribution of internode length of B1、B2 and F2 in seedling stages
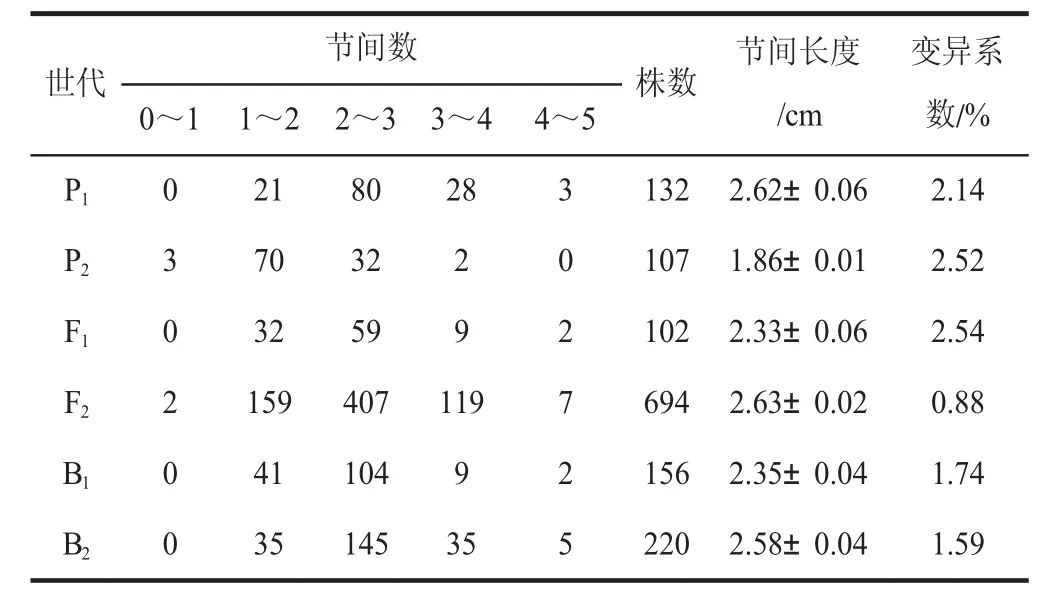
表3 DH 和CH 番茄6 个世代苗期节间长度的次数分布Table 3 Frequency distribution of internode length in 6 generations of DH and CH tomato at seedlings stage
2.2.2 节间长度遗传模型分析
对6 个世代群体节间长度进行遗传模型统计分析,获得5 类24 种番茄节间长度的遗传模型:1MG(1 对主基因)、2MG(2 对主基因)、PG(多基因)、MX1(1 对主基因+多基因)和MX2(2 对主基因+多基因)。
表4 显示了DH 和CH 番茄6 个世代节间长度的24种模型的极大似然数值和AIC值。依据最小AIC值的原则,在表4 中,选择出3 个苗期节间长度的候选模型:MX1-AD-ADI、MX2-ADI-ADI、MX2-ADI-AD。
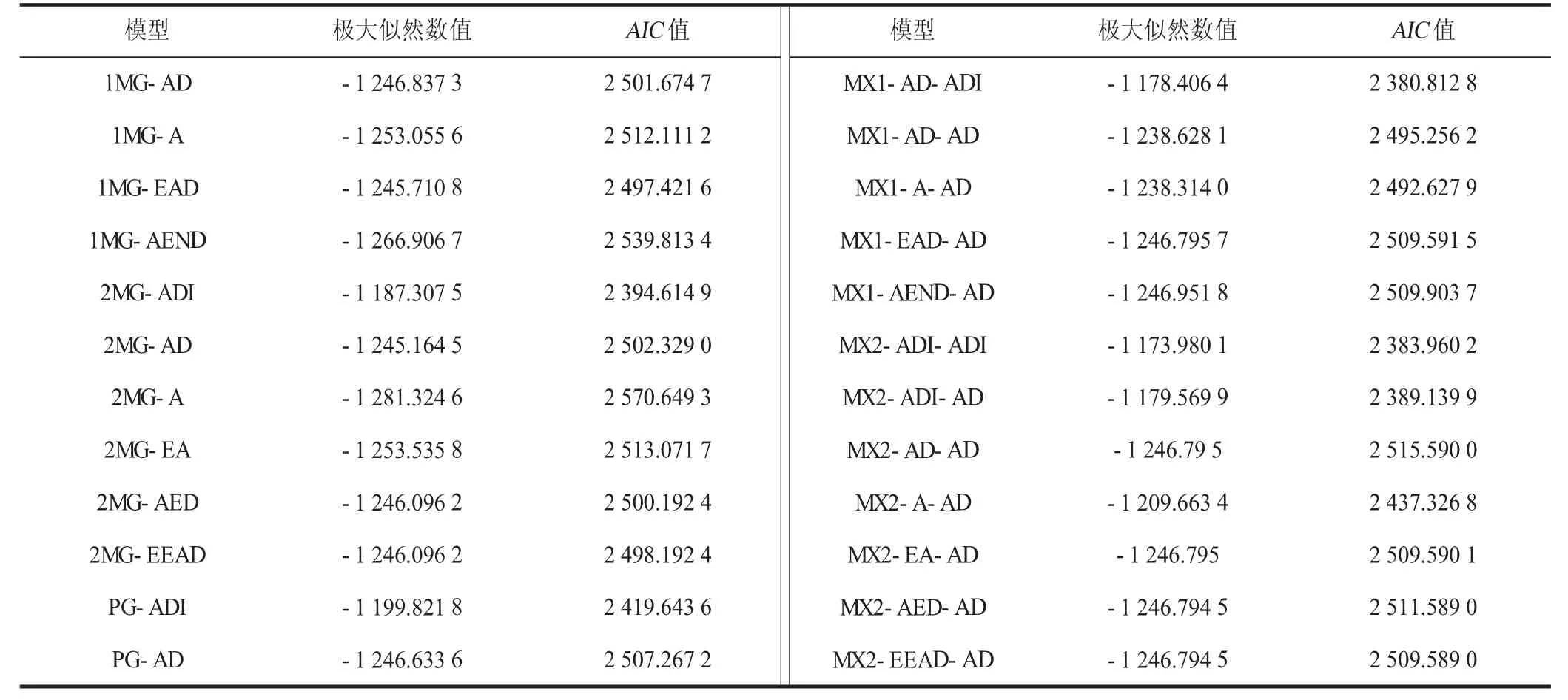
表4 DH 和CH 番茄6 个世代节间长度的24 种模型的极大似然数值和AIC 值Table 4 Maximum likelihood and AIC of 24 models of internode length for 6 generations of DH and CH tomato
2.2.3 节间长度的遗传模型的适合性检验
由表5(见上页)可知,番茄苗期节间长度的MX1-AD-ADI 遗传模型参数达到显著水平的个数为2个,分别为回交群体B1、B2的数值;MX2-ADI-ADI 遗传模型参数达到显著水平的个数为1 个,为回交群体B1的U32数值;MX2-ADI-ADI 模型参数达到显著水平的个数为6 个,回交群体B2的数值也达到极显著水平。依据模型参数达到的显著水平个数最少且AIC值最小为番茄苗期节间长度最适模型的原则[11],确定番茄苗期的节间长度的最优模型是MX2-ADI-ADI(2 对加性-显性-上位性主基因+加性-显性-上位性多基因)。

表5 DH 和CH 番茄6 个世代节间长度的适合性检验Table 5 Suitability test of internode length of 6 generations in DH and CH tomato
2.3.4 节间长度遗传参数的估计
由表6 可知,最适模型MX2-ADI-ADI 的一阶参数中,两对主基因的加性、显性效应值均相等;加性效应高于显性效应;两个主基因加性互作效应明显低于2 个显性基因互作效应。
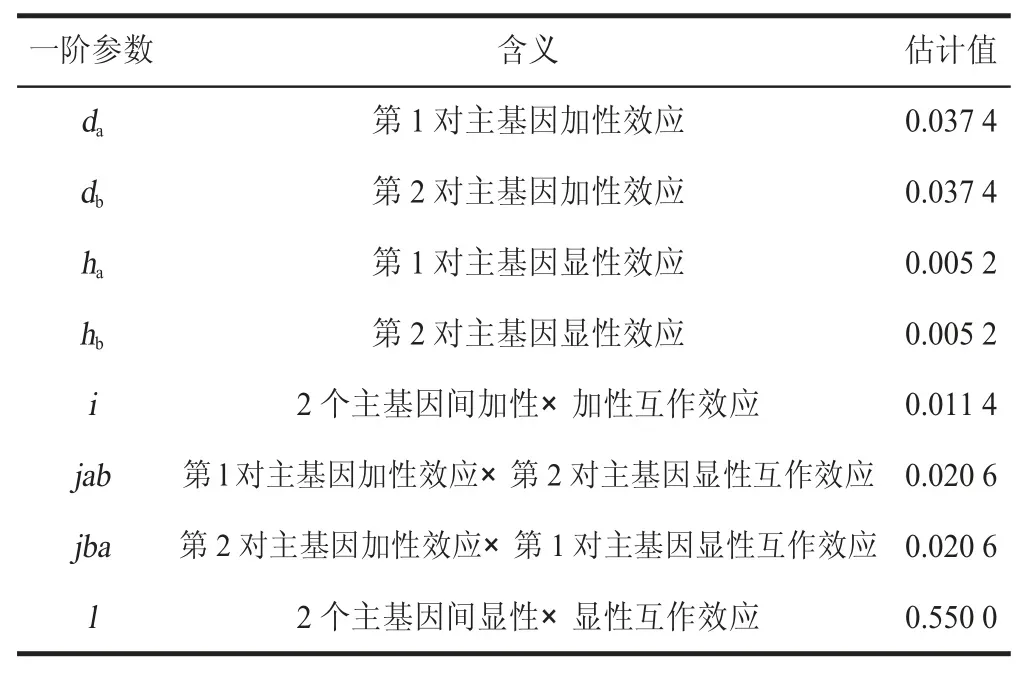
表6 最适遗传模型一阶遗传参数Table 6 Estimation of first-order genetic parameters of optimal genetic model
由表7 可知,最适模型MX2-ADI-ADI 的二阶参数中,B1、B2、F2表型方差分别为0.290 1、0.365 9、0.774 1,环境方差均为0.290 1,主基因方差分别为0、0.075 8、0.484 0,B1、B2、F2主基因的遗传率分别为100%、20.72%、62.52%;多基因方差、多基因的遗传率均为0;分离群体主基因遗传率均高于多基因遗传率。
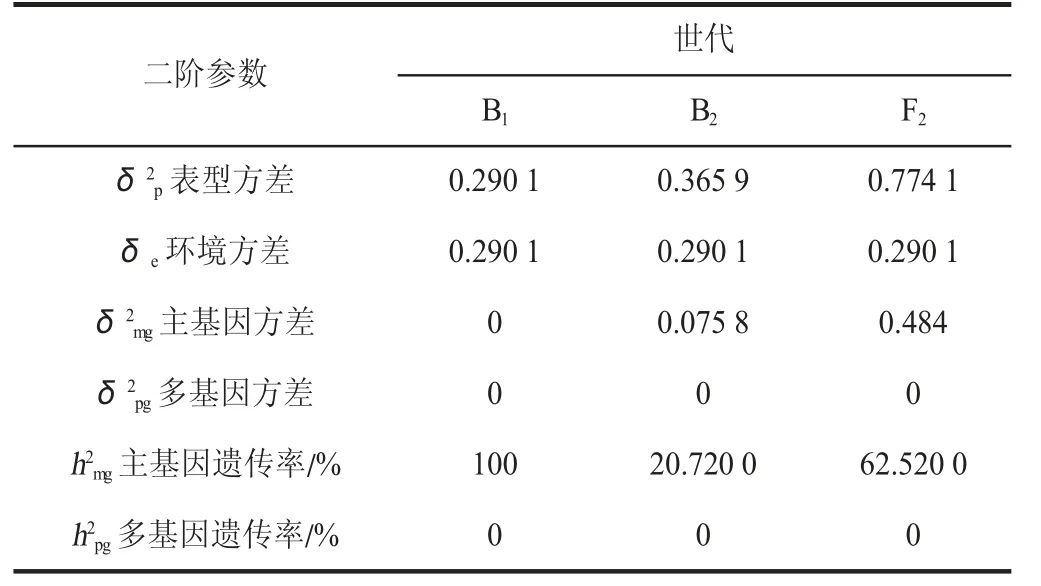
表7 最适遗传模型二阶遗传参数估计值Table 7 Estimation of second-order genetic parameters of optimal genetic model
3 小结与讨论
DH 番茄的上胚轴长度始终高于CH,第一片真叶出现后第18 天2 个品系差异最大。经t 测验,DH 上胚轴长度极显著高于CH,DH 节间长度显著高于CH。番茄CH的6 个世代群体的节间长度遗传模型为2 对加性-显性-上位性主基因+加性-显性-上位性多基因。遗传群体B1、B2、F2节间长度的主基因的遗传率分别为100%、20.72%、62.52%。
本试验所得出的番茄节间长度的遗传模型与前人研究结果不完全相同,如有学者发现野生番茄苗期节间长度可能受1 对或2 对主基因控制[11],这可能与材料遗传物质、试验测定时期、试验栽培环境等因素不同有关。在本试验中,利用6 个世代遗传群体研究发现番茄节间长度的遗传受2 对加性-显性-上位性主基因+加性-显性-上位性多基因控制,为后续的番茄节间长度QTL 定位和分子育种提供了理论基础。
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
农技服务(2023年2期)2023-03-15 00:43:08
热带海洋学报(2022年6期)2022-12-05 12:53:52
阜阳师范大学学报(自然科学版)(2022年1期)2022-04-02 12:35:16
园林科技(2020年2期)2020-01-18 03:28:18
中国稻米(2019年6期)2019-12-05 03:34:22
系统管理学报(2018年3期)2018-08-13 01:05:28
系统管理学报(2018年2期)2018-08-13 01:04:38
上海农业学报(2017年4期)2017-04-10 12:40:22
江西农业学报(2017年12期)2017-02-03 21:49:41
