
苋菜MDH 基因克隆及生物信息学分析
2022-04-09 06:31纪雅婷陈家兰郑友峰赖钟雄刘生财
园艺与种苗 2022年2期
纪雅婷,赵 梅,陈家兰,郑友峰,赖钟雄,刘生财*
(1.福建农林大学园艺植物生物工程研究所,福建 福州 350002;2.西北农林科技大学园艺学院,陕西 杨凌 712100)
苋菜属于石竹目苋科苋属一年生短日照的C4 草本植物[1],富含维生素和矿物质[2],是营养价值极高的蔬菜之一,且茎叶茂盛,生长速度快[1],容易适应土壤环境,对土地没有较严格的要求[3]。作为世界粮农组织推荐的蔬菜,其在世界各地均有种植[1]。
MDH基因广泛存在于动植物和微生物中,其在植物中的作用具有多样性,在光合作用、pH 的平衡和豆科根瘤功能中发挥重要作用[4],参与种子萌发、细胞生长、花粉发育、糖的积累、果实发育和成熟、植物抗逆性等[5]。对光合作用也有重要作用,可催化苹果酸发生的可逆的脱氢反应,在C3 途径叶绿体中苹果酸脱氢酶(malate dehydrogenase,MDH)催化草酰乙酸生成的苹果酸通过草酰乙酸/苹果酸穿越到细胞质中,并氧化产生NADH,为硝酸还原反应提供还原力,且为氨基酸合成提供碳骨架,在NADP-ME 型C4 植物叶肉细胞中,作为C4 光合途径的关键酶,MDH 将叶绿体中的草酰乙酸还原为苹果酸,苹果酸转运到维管束鞘细胞叶绿体中,脱羧释放CO2用于参与卡尔文循环,是参与C4 途径中重要的限速步骤[6-7]。可见MDH 对植物的光合作用有着关键影响,而C4 植物相较于C3 植物有着更高的光合效率,所以MDH基因在C4 植物的表达以及功能有研究意义。
目前人们已经从苹果[8]、小麦[8]、花生[9]、香蕉[10]、木薯[11]等植物中克隆出MDH 基因,已有对其在非生物逆境胁迫应答[12]和超量表达提高对铝毒的耐受性[13]等方面的研究,但对在C4 植物苋菜中MDH基因研究较少。因此,该研究对苋菜中的MDH基因进行克隆和生物信息学分析,为将来深入研究该基因功能奠定基础。
1 材料与方法
1.1 材料
使用福建农林大学园艺植物生物工程研究所提供的苋菜(Amaranthus tricolorL.)无菌试管苗作为研究材料。
1.2 方法
1.2.1 苋菜试管苗总RNA 提取及质量检测。使用多糖多酚植物总RNA 快速提取试剂盒(百泰克公司)对苋菜试管苗总RNA 进行提取。用琼脂糖凝胶电泳对RNA 完整性进行检测,后使用超微量核酸检测仪(Thermo 公司)测定RNA纯度和浓度。
1.2.2 cDNA 合成。使用Fermentas 公司RevertAidTMFirst-Strand cDNA Synthesis Kit 试剂盒合成cDNA 的第一链。
1.2.3 苋菜MDH基因引物设计及PCR 扩增。根据福建农林大学园艺植物生物工程研究所实验室建立的苋菜转录组数据库(SRA:SRR924089-SRR924092),从中查找MDH基因序列片段,通过NCBI 比对确定ORF 区域,然后根据序 列 设 计ORF 引 物(MDH-F:CGGCATGAGAACTC AAATGTT,MDH-R:CTCAATTGTTCTTCCTTGCGA)进行扩增。
1.2.4 目的片段的回收、克隆及测序。使用Biomiga 公司The Inventor of EZgeneTMand ViraTrapTMSystems Gel/PCR Extraction Kit 琼脂糖凝胶DNA 回收试剂盒,以pMD18-T为载体,转化到感受态细胞DH5 中,再挑取单克隆进行摇菌培养,通过菌液PCR 初步验证阳性克隆,再将菌液送交北京六合华大基因科技股份有限公司进行测序。
1.2.5 苋菜MDH基因序列生物信息学分析。使用DNAMAN7.0 进行序列拼接及氨基酸序列推导,再用在线软件对该基因进行生物信息学分析,并用MEGAX 软件邻解法对不同物种间MDH基因的关系进化树进行构建。
1.2.6MDH基因表达量分析。对苋菜进行蓝光处理和黑暗处理,并分别测其MDH基因FPKM 值,以及检测其在生长不同阶段MDH基因的RPKM 值,对MDH基因表达差异进行分析,利用EXCEL 软件绘制柱状图和折线图。
2 结果与分析
2.1 苋菜MDH 基因的ORF 克隆及其编码的蛋白质理化性质分析预测
根据构建的苋菜转录组数据库,筛选出苋菜MDH基因cDNA 片段,通过NCBI 对比确定ORF 区域,利用克隆技术获得苋菜MDH基因ORF 序列,提交至GenBank(登录号:KY353102)。
利用ProtParam 对苋菜中MDH 基因编码的蛋白质理化性质分析。结果显示,该蛋白质分子式为C1615H2618N438O494S12,由5177 个原子构成,包括344 个氨基酸,相对分子量为36459.91 Da,理论等电点(pI)8.65,带正电的氨基酸有38(Arg+Lys)个,带负电的氨基酸有34(Asp+Glu)个,总平均亲水性为-0.024,属于疏水蛋白,同时也是稳定蛋白(不稳定指数为37.96)。
对MDH基因编码蛋白进行生物信息学分析,结果显示,MDH基因在多肽链的存在结合位点,属于Ldh_1_C 超家族;该蛋白没有明显的信号肽,不属于跨膜蛋白,且在细胞膜外;亚细胞定位预测表明该蛋白位于叶绿体和线粒体中。苋菜MDH基因编码的蛋白质磷酸化位点(>0.5)总数为33,其中包括18 个丝氨酸,12 个苏氨酸和3 个酪氨酸。
蛋白二级结构预测分析显示,苋菜MDH蛋白中α 螺旋占43.6%,无规则卷曲占34.88%,β 折叠占5.81%(图1)。三级结构进行预测分析表明该蛋白由无规则卷曲结构,β折叠和α 螺旋构成(图2)。

图1 MDH 编码蛋白质二级结构预测
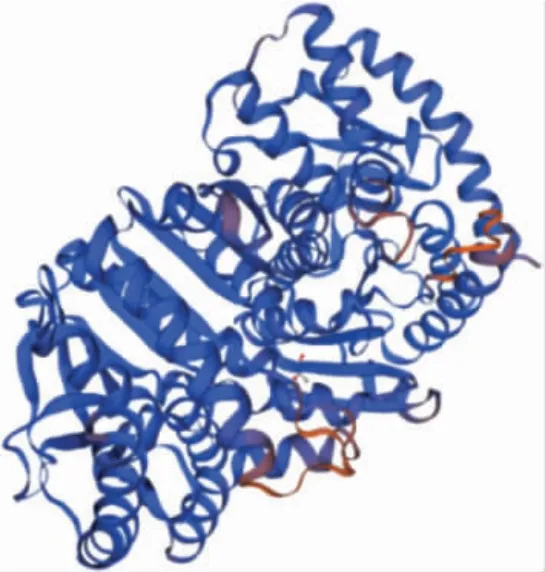
图2 MDH 编码蛋白质三级结构预测
2.2 苋菜MDH 基因编码的蛋白系统进化树构建
通过对苋菜MDH基因编码的氨基酸序列在线BLAST 的结果,选取不同同源性的物种氨基酸序列,用MEGAX 软件中NeighborJoining 法将Bootstrap 检验值设置为1000,建立其编码区氨基酸序列之间的系统关系树。根据MDH 氨基酸序列之间关系树的构建分析可知,苋菜MDH基因与藜麦聚类到一起,同源性较高,因苋菜与藜麦同属石竹目,亲缘关系较近,可知结果可信(图3)。
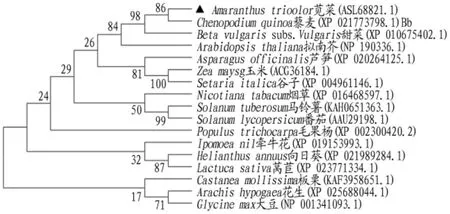
图3 MDH 编码蛋白质系统进化树
2.3 MDH 基因表达量分析
根据实验室提供的在蓝光处理与黑暗处理下苋菜转录组数据的MDH 基因FPKM 值、苋菜不同生长阶段中MDH 基因RPKM 值以及在叶片红绿部位的FPKM 值利用GraphPad Prism8 软件进行绘图,并进行MDH 基因表达量分析。由图4 可知,在黑暗处理下基因表达量高于蓝光处理,在苋菜幼苗期至壮苗期基因表达量出现了下降而花芽期到开花期基因表达量迅速增长,在叶片红色部位基因表达量远高于绿色部位,推测苋菜在黑暗条件下MDH基因表达更显著在花芽期至开花期表达量上升且在叶片红色部位表达更显著。

图4 苋菜MDH 基因相对表达量分析结果
3 讨论与结论
该试验通过克隆技术获得了苋菜MDH 基因ORF 序列,并提交GenBank 登录。蛋白系统进化树构建等各种生物信息学分析研究,得出苋菜中MDH 基因可能编码一种位于叶绿体和线粒体的非分泌蛋白,二级结构以α 螺旋为主,三级结构由无规则卷曲结构,β 折叠和α 螺旋构成,与藜麦有较近的进化关系,在黑暗条件下和叶片红色部位表达更显著且在开花期较其他生长时期更显著。
MDH 基因广泛存在于动植物以及微生物体内,并且参与光合作用多种生理活动,尤其是C4 途径中的关键基因之一。该试验对MDH 基因的亚细胞定位预测结果中MDH 可能编码的非分泌蛋白位于叶绿体及线粒体中,结合其他学者在研究小麦和谷子中苹果酸脱氢酶活性与光合的关系时,发现谷子的MDH 均与净光合速率成正相关,以及其在C4 植物中广泛存在,在草酰乙酸转化为苹果酸的过程中发挥作用[14],推测MDH 基因在苋菜中与苋菜的光合作用有关且可能发挥同样的作用。MDH 基因在生物体内的存在非常广泛,其中苋菜中MDH 氨基酸序列与藜麦同属石竹目,有较近的亲缘关系,有研究关注其在谷子[12]中可能参与了抗逆性胁迫的响应,也有研究其超量表达在苜蓿[13]、烟草[15]等植株中对提高铝毒耐性的作用,苋菜中MDH 基因与谷子、烟草等有一定亲缘关系,其在苋菜中是否有相似功能还有待研究。根据对其基因表达量分析,在黑暗条件下及叶片红色部位表达量更高推测其可能参与呼吸作用的作用机制,且可能更加有效地提高CO2利用率,在壮苗期表达量下降而在花芽期至开花期表达量上升,推测其可能对植物生殖生长发挥更大作用。对苹果酸脱氢酶的研究已有很长时间,且研究较深入,已有学者对其在国内外的研究进展进行追溯[16]。可见MDH 基因是一个具有重要功能,有着长久研究价值和意义的基因。
猜你喜欢
环球时报(2022-09-20)2022-09-20
中国现代医生(2022年21期)2022-08-22
中国现代医生(2022年21期)2022-08-22
课外语文·中(2022年1期)2022-02-16
老友(2021年6期)2021-07-01
百科知识(2021年2期)2021-02-24
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
科技新时代·e医疗(2011年8期)2011-12-23
