
槟榔林下间作平托花生土壤细菌群落结构及多样性研究
2022-04-08 07:39:20颜彩缤胡福初张世青范鸿雁
中国土壤与肥料 2022年2期
颜彩缤,赵 亚,胡福初,芮 凯,陈 哲,张世青,范鸿雁*
(1.海南省农业科学院三亚研究院,海南 三亚 572000;2.海南省农业科学院热带果树研究所,农业农村部海口热带果树科学观测实验站,海南省热带果树生物学重点实验室,海南省热带果树育种工程技术研究中心,海南 海口 571100;3.海南省农业科学院植物保护研究所,海南 海口 571100)
槟榔(Areca catechu L.)是中国四大南药之一,棕榈科常绿乔木是海南第二大特色经济作物,其果实被当作咀嚼嗜好品,具有很高的经济价值[1]。槟榔原产于马来西亚,于热带、亚洲热带地区广泛栽培,在我国主要分布在海南、云南及台湾等地区[2]。随着槟榔种植面积扩大,槟榔黄化现象发生范围广、蔓延快,给产量造成了严重损失,据不完全统计,槟榔减产达10%~60%不等,每年因黄化现象损失20 亿元以上[3]。平托花生又称蔓花生、野花生,是一种热带亚热带豆科牧草,多年生宿根草本,生长能力强,可无性繁殖和种子繁殖,原产于巴西,主要用于人工观赏草坪、生物覆盖和果园生草栽培。本研究选用的‘热研12 号’平托花生是1991 年中国热带农业科学院热带牧草研究中心从哥伦比亚国际热带农业研究中心引进,其在热带亚热带地区能保持常绿并周年生长开花,广泛适应华南地区酸性红壤种植,不仅具有耐酸瘠及贫瘠土壤、耐旱和耐荫的特点,而且还具有增加土壤肥力、提高保水性能、改善园内小气候和控制杂草生长等作用,是生态果园优良的间作植物,间作时间越长保水性能越好[4-6]。
土壤细菌是土壤微生物类群的重要组成部分,主要参与有机物分解、生物地球化学循环等活动,几乎与土壤中所有生化反应都有关系,不仅在改善土壤肥力以及生态系统结构方面有显著作用[7],而且具有调节植物生长发育和防治病虫害等重要作用[8]。槟榔黄化发生严重,但目前尚未找到有效的防治方法,槟榔间作是重要的农业防治方法之一,可以预防和减缓该病害的发生。国外在槟榔园中混合种植了小豆蔻、菠萝、生姜、姜黄、香蕉、可可和黑胡椒等多种作物,我国仅单一间作香草兰、胡椒和糯米香茶等作物[9]。Castro 等[10]和Sujatha 等[11]研究发现槟榔园间作香草兰可提高土壤pH、土壤速效养分含量以及单位面积经济效益;吴凤芝等[12]提出间作具有显著增加土壤微生物群落多样性、减轻病害发生以及提高黄瓜产量的作用;王华等[13]研究槟榔不同株行距间作香草兰时发现,土壤细菌数量较香草兰单作显著提高,且细菌数量与有机质、碱解氮、有效铁和有效硼等含量呈显著正相关;戴晋等[14]的研究表明,辣椒小麦套种、再间作芝麻的种植模式可减少辣椒“日灼病”的发生率;冯晓敏[15]研究发现,燕麦间作豆科作物不仅对燕麦根部土壤微生物环境具有良好的调节作用,而且能增加燕麦产量。
目前,我国关于槟榔林下间作平托花生对土壤细菌群落结构及多样性的影响还尚未见报道。平托花生根系长度经测量平均值为10 ~20 cm,为探讨槟榔林下间作平托花生土壤细菌群落结构及其多样性,本试验以槟榔林下间作平托花生和槟榔单作地区10 ~20 cm 土层的土壤为研究对象,采用Illumina Miseq 高通量测序技术对土壤细菌群落V4 ~V5 区片段进行测序和生物信息学分析,从分子水平上揭示槟榔林下间作平托花生和槟榔单作土壤细菌群落结构及其多样性的变化,以期为深入研究槟榔林下间作平托花生对土壤细菌多样性的影响和防治槟榔黄化病提供一定的理论参考。
1 材料与方法
1.1 试验材料
供试槟榔品种为‘热研1 号’,于2010 年定植,种植规格为2 m×2.5 m;间作物平托花生品种为‘热研12 号’,2019 年初定植,平托花生于槟榔林下全覆盖种植,本试验平托花生不进行收割还田。
1.2 试验地概况
试验地位于海南省琼海市大路镇礼合村委会的槟榔种植园(110.47026°E,19.449614°N),海拔31 m,属热带季风及海洋湿润气候,年平均气温24 ℃,年平均降水量2072 mm,年平均日照2155 h,年平均辐射量118.99 kJ/m2。土壤为砖红壤,多为砂质粘壤土。
1.3 试验方法1.3.1 试验设计
试验设2 个处理,槟榔林下间作平托花生(BP)和槟榔单作(CK),每个处理3 个重复,分别编号为BP-1、BP-2、BP-3 和CK-1、CK-2、CK-3。
1.3.2 土壤样品采集
2019 年12 月分别在槟榔林下间作平托花生和槟榔单作试验处理区,在平托花生营养生长期采集土壤样品,每个处理重复3 次,每个重复采取五点取样法选择5 株槟榔,在距离槟榔茎基部约15 cm处的东西南北4 个点,拔开平托花生茎蔓后挖取10 ~20 cm 土层根际土壤,混匀后装入无菌瓶,密封好放入装有冰袋的保温箱带回实验室,在超净工作台将无菌瓶内土壤样品的杂根去除,分装于50 mL 灭菌离心管,-20℃下保存,用于高通量测序;与上述选点取样方法相同,取样后采用四分法去除多余土壤,装入密封袋带回实验室,过1 mm 筛,常温下保存,用于土壤理化因子分析。
1.3.3 土壤理化因子的分析
测定方法参考文献[16]。采用水合热重铬酸钾氧化法测定土壤有机碳;采用半微量凯氏定氮法测定全氮;采用碱解扩散法测定碱解氮;采用王水酸溶钼锑抗比色法测定全磷;采用双酸浸提钼锑抗比色法测定酸性土壤有效磷;采用碱熔—火焰光度法测定全钾;采用醋酸铵浸提—火焰光度法测定速效钾;采用铝盒烘干法测定干湿比;采用酸度计测定土壤pH。
1.3.4 土壤总DNA 的提取
使 用FastDNA®SPIN Kit for Soil 试 剂 盒 提 取样本土壤微生物基因组DNA,干冰运输,由上海天昊生物科技有限公司采用Illumina Miseq 测序平台对土壤细菌进行16S 扩增子绝对定量测序(V4 ~V5)。
1.4 数据处理
对原始数据进行质量控制,获得更为精准、高质量、可用于分析的序列;应用usearch、QIIME根据序列97%的相似度,将序列归并划分为多个OUTs。利用Mothur、R 计算丰富度指数Chao1 和ACE 以及多样性指数Simpson 和Shannon,并进行Alpha 多样性分析;同时进行细菌群落分布、聚类分析和细菌功能预测分析。用SPSS 20.0 独立t检验分析土壤理化因子差异性,用Canoco 5 构建细菌菌门与土壤理化因子的相关性,用SPSS 20.0 Spearman’s rho test 分析细菌菌属和土壤理化因子的相关性。
2 结果与分析
2.1 间作对土壤理化因子的影响
由表1 可知,槟榔林下间作平托花生后,土壤理化因子与槟榔单作有显著性差异。间作模式下酸性有效磷含量极显著提高95.99%;全磷含量显著提高33.33%;全钾含量显著提高32.64%;速效钾含量显著提高21.93%;有机碳、全氮和碱解氮无显著性差异;土壤pH 显著提高。

表1 土壤理化性质
2.2 不同种植模式下土壤细菌群落多样性变化
2.2.1 Alpha 多样性分析
Chao1 指数和ACE 指数侧重于体现群落丰富度,Shannon 指数和Simpson 指数兼顾群落均匀度,均与群落多样性成正比,结果见表2。槟榔间作模式较单作模式下的物种数目平均显著增加5.5%,Chao1 指数、ACE 指数分别显著提高4.95%、4.60%,表明间作后物种丰富度明显提高,而Shannon 指数、Simpson 指数均提高但未达到显著水平。物种覆盖率达到99%以上,说明土壤微生物样本序列检出率高。

表2 不同样品的α 多样性指数
2.2.2 Beta 多样性分析
本研究通过非度量多维尺度分析(NMDS)方法[17],观测样本之间的差异。每个点代表1 个样本,两点之间的距离越近,表明2 个样本之间的细菌群落结构相似度越高。图1 是对槟榔单作和间作模式下土壤中OTU 组成的分析,由图1 可知,间作与单作各自的3 个土壤样品分别聚类,说明两组土壤样品的细菌群落结构具有差异。
2.3 土壤细菌群落组成分析
2.3.1 土壤细菌群落结构在门水平的分析
两种槟榔种植模式下土壤细菌群落结构平均相对丰度大于1%的门类群共有12 个,相对丰度较高的门类群分别是变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetes)、奇古菌门(Thaumarchaeota)、拟杆菌门(Bacteroidetes)和芽单胞菌门(Gemmatimonadetes)等(图2)。结果表明,槟榔林下间作平托花生后并没有改变细菌门水平上的主要种类组成,但提高了变形菌门、放线菌门、浮霉菌门和芽单胞菌门等的相对丰度,其中变形菌门、放线菌门的相对丰度分别增加2.57%、2.84%;酸杆菌门、绿弯菌门、奇古菌门和拟杆菌门等的相对丰度均降低,尤其是酸杆菌门、绿弯菌门相对丰度降低幅度较大,分别降低4.61%、2.31%。在单作模式下酸杆菌门相对丰度最高,而间作模式下变形菌门相对丰度最高。
2.3.2 土壤细菌群落结构在属水平的分析
供试细菌菌群平均相对丰度大于1%的属类群共有15 个,详见图3。酸杆菌门的Gp1、Gp2、Gp4、Gp5、Gp6 和Gp7 亚 群 均 为 优 势 菌 属;其中,间作提高Gp6、Gp4 的相对丰度但降低了其余Gp 属的相对丰度,尤其是Gp6 增加幅度最大(为1.79%);Gp1、Gp2 和Gp5 比例分别降低2.79%、2.44%和0.14%;Gp6 在单作和间作模式下相对丰度均最高。相对丰度较高的还有亚硝化球菌属(Nitrososphaera)、Gaiella、芽单胞菌属(Gemmatimonas)、Pseudolabrys、硝化螺菌属(Nitrospira)和红游动菌属(Rhodoplanes)等菌属,其中间作提高Gaiella、芽单胞菌属和Pseudolabrys 的相对丰度,降低亚硝化球菌属、硝化螺菌属和红游动菌属的相对丰度;Gp3(1.3%)和Terrimonas(1.2%)在槟榔单作下为优势菌属。两种槟榔种植模式下无明确分类信息或分类名称(No_Rank)的菌属占25.09%~27.51%,无任何分类信息(Unassigned)的菌属占18.40%~17.43%。
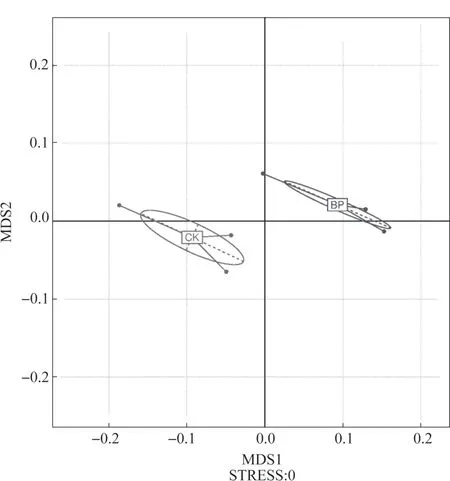
图1 非度量多维尺度分析图
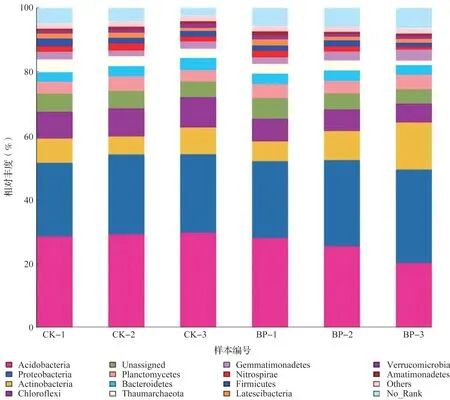
图2 土壤门水平上的细菌群落组成
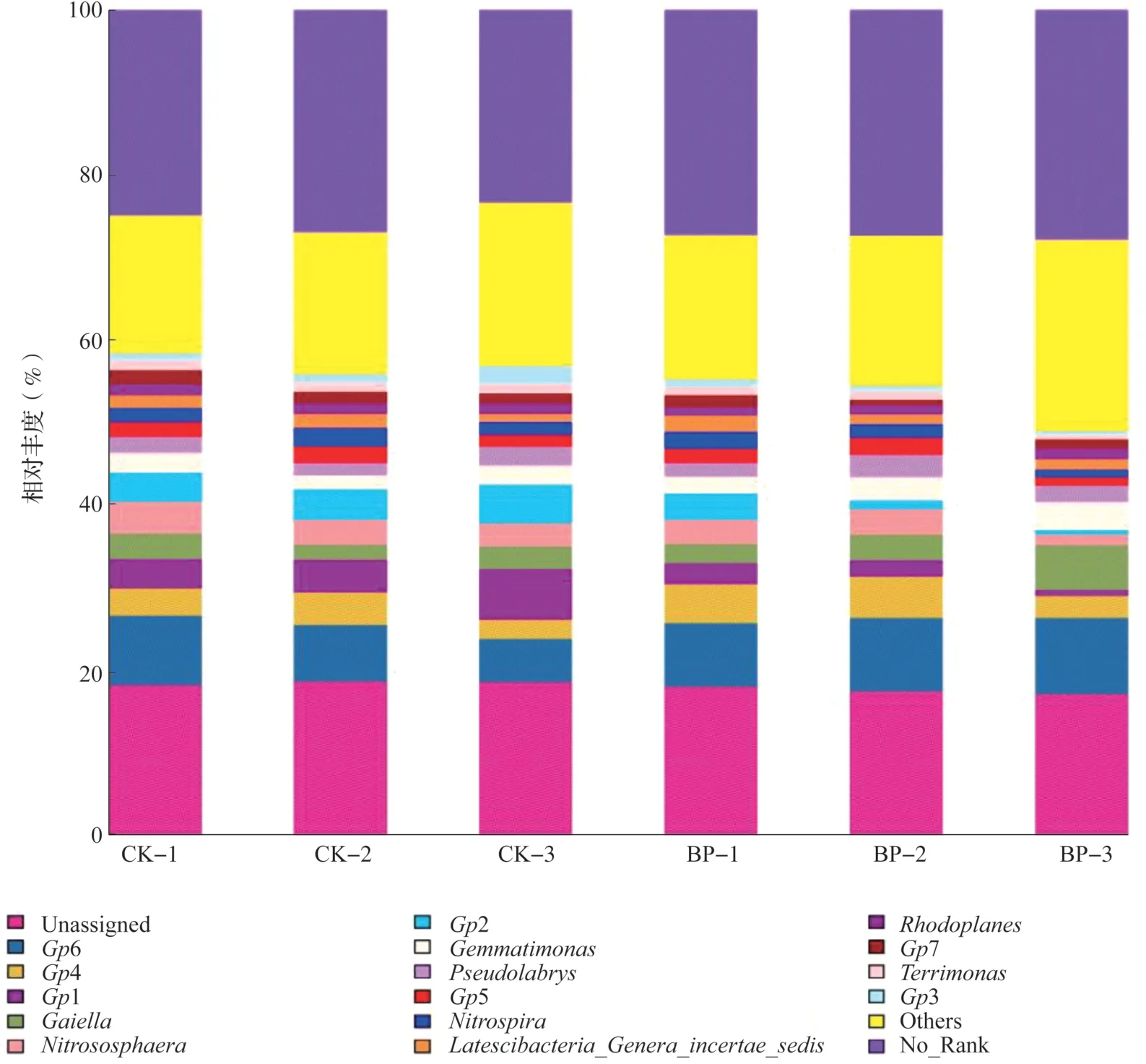
图3 土壤属水平上的细菌群落组成
2.4 土壤细菌与理化因子的相关性分析
2.4.1 门水平上土壤细菌与理化因子的相关性分析
在门分类水平上,对土壤总细菌中相对丰度大于1%的细菌进行冗余分析,以探究特定细菌与土壤理化因子的关系。总变异数为0.01839,解释变量占96.5%(调整后解释变化为82.3%),排序轴1 对土壤总细菌变异程度的解释度为83.96%,排序轴2 解释了8.71%的变异,对细菌群落结构的解释度总共高达92.67%(表3)。其中速效钾解释了最多的变异程度,解释度为76.2%,有机碳、全氮、干湿比解释度分别为3.5%、7.6%、9.1%(表4)。由图4 可见,放线菌门、芽单胞菌门、变形菌门与速效钾、有机碳、全氮和干湿比均呈正相关;酸杆菌门、绿弯菌门、拟杆菌门、疣微菌门(Verrucomicrobia)与速效钾和干湿比呈负相关。单作模式(CK)的土壤样品主要分布在第三象限全氮和有机碳含量延长线之间,说明CK 细菌群落结构差异主要是由全氮和有机碳含量的变化引起的;间作模式(BP)的土壤样品分布第一(BP-3)、第三(BP-1)和第四象限(BP-2),由于BP-1 与CK 聚类很近,因此作为异常值不做考虑,说明其细菌群落结构主要受速效钾和干湿比的影响。
2.4.2 属水平上土壤细菌与理化因子的相关性分析

表3 土壤因子与土壤细菌门水平RDA 分析结果
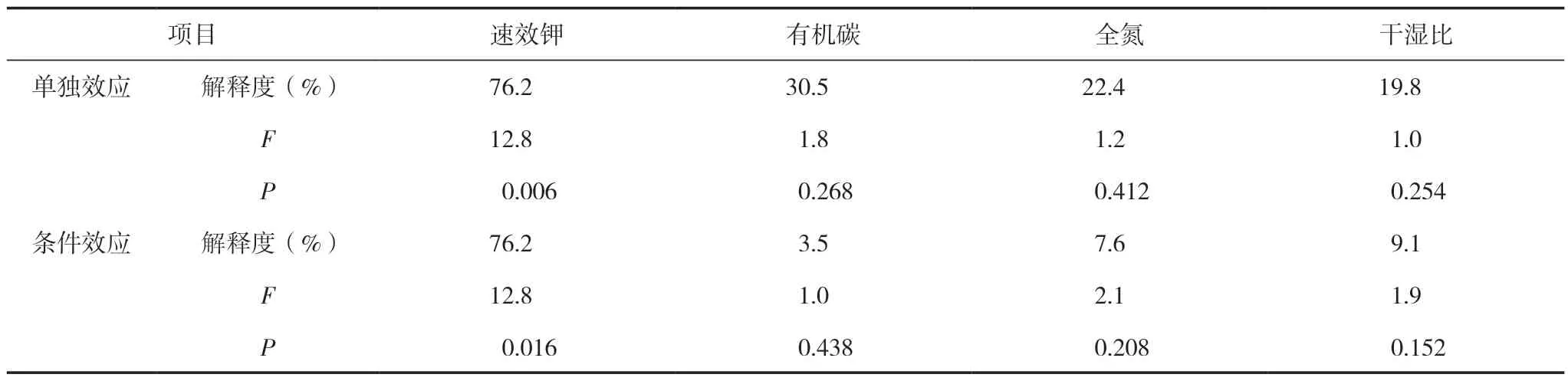
表4 主要土壤因子RDA 结果
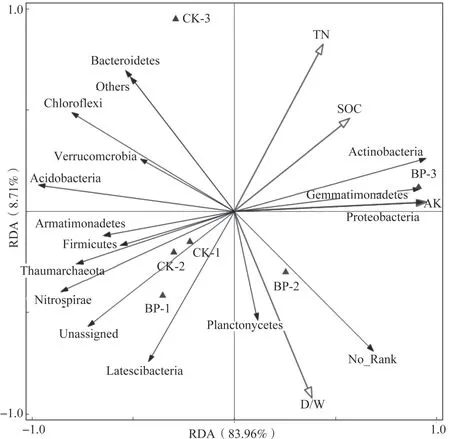
图4 门水平上土壤总细菌类群与土壤理化因子之间的关系
在属分类水平上,对土壤总细菌中相对丰度大于1%的细菌进行相关性分析,以了解优势菌属与土壤理化因子的相关性。亚硝化球菌属与有机碳、碱解氮呈极显著负相关;硝化螺菌属与全磷、全钾分别呈显著和极显著负相关;Gp1、Gp2 和Gp3 均与pH 呈极显著或显著负相关;Gp6 与pH 呈显著正相关;Gp4 与全氮呈显著负相关;红游动菌属与全氮呈显著正相关;Gaiella、芽单胞菌属和Pseudolabrys均与全钾呈极显著或显著正相关。

表5 属水平上土壤细菌与土壤理化因子相关性分析
2.5 土壤细菌菌群代谢功能预测
对两种槟榔种植模式的细菌菌群功能进行预测和分析,细菌基因序列中共发现来自6 类代谢通路的41 个子功能类群,选择相对丰度最高的top30 功能绘制热图。氨基酸代谢、碳水化合物代谢、膜运输、能量代谢是细菌群落的主要代谢功能群。单作和间作模式下菌群功能差异性分析结果见图5,间作平托花生后能量代谢的功能基因显著增加,影响苯丙素、类黄酮、甜菜红色素和吲哚生物碱等次生代谢产物的生物合成功能基因、辅助因子和维生素的代谢以及控制昼夜节律-植物的功能基因显著减少,说明间作除了提高细菌能量代谢功能外,其余代谢功能均降低。
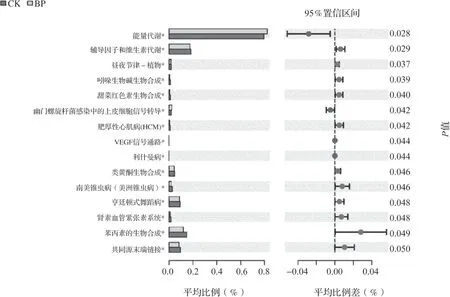
图5 t-检验差异KEGG 功能相对丰度条形图
3 讨论
3.1 间作对土壤理化性质的影响
磷和钾是植株生长发育所必需的基本元素,具有促进植物根系发育、防止植株瘦弱低矮、提高作物产量和品质以及抵抗不良土壤环境等作用,土壤全磷、全钾含量即反映磷和钾的贮量状况,土壤有效磷、有效钾容易被作物吸收利用,是体现土壤磷、钾养分供应水平的指标[18-21]。本研究中,槟榔林下间作平托花生显著提高了土壤酸性有效磷、全磷、全钾以及速效钾含量,尤其是土壤酸性有效磷含量极显著提高了95.99%(表1),说明间作平托花生后土壤磷、钾的贮量和供应能力均显著或极显著提高。初步分析是由于土壤pH 影响离子的选择性,在土壤pH 大于5.5 时,磷离子、钾离子与铝离子和Al(OH)x在竞争吸附位点中占优势,使溶液中的磷、钾易被土壤吸附[22-23]。本研究结果与其一致,单作模式下土壤pH 为5.47 而间作模式为5.72,说明间作平托花生能够提高土壤pH,从而导致部分养分含量增加,最终能够改善由于缺素造成的槟榔黄化现象。
3.2 间作对土壤细菌群落多样性的影响
前人在研究槟榔根际土壤微生物与槟榔黄化病的关系时发现,根际土壤细菌数量随着黄化程度增加呈减少的趋势,即根际细菌数量表现为健康槟榔>轻度黄化槟榔>中度黄化槟榔,细菌数量还与土壤有机质、全氮、有效磷呈显著或极显著正相关[24-25]。本研究通过间作平托花生发现,物种数目以及Chao1 指数、ACE 指数均显著提高(表2),说明间作平托花生可以提高土壤细菌数量及群落多样性,从而增强植株抗黄化能力。
3.3 间作对土壤细菌群落结构的影响
两组样本的土壤细菌群落结构存在一定差异(图2、3),各菌群相对丰度的不同反映了土壤环境的变化。在门水平上,两种模式下变形菌门、酸杆菌门、放线菌门、绿弯菌门均为优势菌群。变形菌门是反映土壤营养状况的重要指标之一[26],其相对丰度在营养较为丰富的土壤中更高[27],表现为随着土壤有机质含量增多而增多[28-29]。本研究中,槟榔间作模式下变形菌门的相对丰度提高了2.57%,说明间作平托花生提高了土壤营养丰富度。放线菌作为植物根际土壤的一种重要微生物,有促进植物生长发育、防治病害的作用[30-31],放线菌门的相对丰度与土壤碳氮比呈显著正相关关系[32],因此,间作平托花生后放线菌门相对丰度的显著提高,导致土壤碳氮比提高以及土壤营养结构优化。酸杆菌是广泛分布在土壤中的嗜酸菌[33-35],酸杆菌门的比例体现土壤的酸性条件[36-37],已有研究表明,酸杆菌门与土壤pH 呈极显著负相关[38],而且土壤pH 降低能够导致细菌群落多样性减少[39]。本研究间作模式下酸杆菌门相对丰度降低4.61%,pH 显著增加(表1),说明槟榔林下间作平托花生能够增加土壤细菌群落多样性以及改良土壤酸化状况;在间作模式下优势亚群中Gp6、Gp4 增加,Gp1、Gp2 和Gp3 菌群均减少;pH 与Gp1、Gp2 和Gp3 呈 显 著 负 相 关 但 与Gp6 呈显著正相关(表5),说明土壤环境pH 主要受到Gp1、Gp2、Gp3 以及Gp6 菌属的影响,这与前人得出的Gp1、Gp2 和Gp3 的相对丰度在酸性土壤中较高[40]的研究结果一致。绿弯菌门在贫瘠土壤条件下更具优势,与全氮和碱解氮呈极显著负相关[41],与本研究结果不一致,间作平托花生后土壤细菌中绿弯菌门的相对丰度降低2.31%,全氮含量降低但差异不明显(表1),绿弯菌门与全氮含量呈正相关,考虑平托花生为豆科作物,具有固氮作用;对两种模式下土壤优势细菌菌属的相对丰度进行比较,发现具有显著差异的还有Terrimonas 属,Terrimonas 属为拟杆菌门(Bacteroidetes)、鞘脂杆菌纲(Sphingobacteriia)、鞘脂杆菌目(Sphingobacteriales)、泉发菌科(Crenotrichaceae),是一类能够降解蒽类物质的好氧反硝化细菌[42]。本研究中槟榔单作模式下Terrimonas 的相对丰度达1.2%,说明间作平托花生后土壤环境发生改善,从而抑制Terrimonas 细菌生长,本研究在属水平下的No_Rank 和Unassigned 菌属在两种模式所占比例分别为25%以上和17%以上,说明槟榔林下土壤还存在大量需要进一步鉴定研究的未知或稀有细菌。
3.4 土壤细菌与理化因子的相关性分析
从相关性分析(图4,表5)得出速效钾、有机碳、全氮和干湿比是影响土壤细菌门结构的主要环境因子。速效钾解释度最高,其含量与放线菌门、芽单胞菌门和变形菌门呈极显著正相关,与酸杆菌门、绿弯菌门、拟杆菌门、疣微菌门以及Gp1、Gp2 呈显著负相关;Gaiella、芽单胞菌属和Pseudolabrys 均与全钾呈极显著或显著正相关。因此,钾元素为影响土壤细菌群落结构变化的主要理化因子。与氮有关的菌属为亚硝化球菌属、Gp4 菌属和红游动菌属,亚硝化球菌属与碱解氮呈极显著负相关,红游动菌属与全氮呈显著正相关,Gp4 菌属与全氮呈显著负相关,间作均降低了亚硝化球菌属、红游动菌属的相对丰度,但提高了Gp4 菌属的相对丰度,说明间作平托花生具有良好的固氮效果,并且可以改变土壤细菌群落结构。单作模式细菌群落结构差异主要由全氮和有机碳含量的变化引起,间作模式细菌群落结构主要受速效钾和干湿比的影响。本研究结果表明,影响两种模式下土壤细菌群落结构的主要理化因子多数差异不显著,因此,有必要进一步开展间作模式的改良以及持续监测。
3.5 土壤细菌代谢功能预测
在揭示土壤细菌群落组成的基础上,进行菌群代谢功能预测(图5)能反映环境因子的变化,为打破传统耕作、实现持久耕作奠定基础。两组样本的土壤细菌群落主要代谢功能为氨基酸代谢、碳水化合物代谢、膜运输和能量代谢。能量代谢功能主要表现为细菌有机物分解或无机物氧化过程中释放的能量,通过底物磷酸化或氧化磷酸化合成ATP的过程[43],间作后细菌能量代谢功能增强的原因可能是细菌群落结构的改变,尤其是与化能异养菌、光能营养菌还有化能自养菌相对丰度密切相关;但化能异养菌如Terrimonas 和化能自养菌如硝化螺菌属相对丰度的减少还有待进一步研究;间作后苯丙素、类黄酮、甜菜红色素和吲哚生物碱的生物合成功能基因显著减少,其原因可能是苯丙素、类黄酮和甜菜红素等是在植物逆境胁迫下产生的次生代谢物质[44-46],而间作平托花生后改善了土壤环境因子;崔晓莹等[47]探究发现丛枝菌根真菌接种连作大豆根系后的辅助因子和维生素的代谢功能基因均减少;董艳辉等[48]研究表明藜麦连作后的辅因子和维生素代谢功能基因减少,与本研究结果一致;昼夜节律是植物通过转录-翻译反馈调节环路对环境中的信号如温度和光照进行整合从而适应外界环境的一种内在分子机制,几乎影响植物新陈代谢的方方面面[49-50],本研究间作后植物昼夜节律功能基因减少,原因可能是温度和光照等环境因子的改变对植株生物钟造成影响。综上,代谢功能基因增减与土壤理化性质以及环境因子改变密切相关,对槟榔种植具有良好的指导意义。
4 结论
间作模式下酸性有效磷、全磷、全钾和速效钾含量分别显著或极显著增加95.99%、33.33%、32.64%和21.93%,土壤pH 显著提高,槟榔林下间作平托花生后土壤质量以及酸化状况得到明显改善。
间作模式下细菌菌群数量增加5.5%,Chao1 指数、ACE 指数分别显著增加4.95%、4.60%,表明槟榔林下间作平托花生可有效提高土壤细菌丰富度以及多样性。
间作还能提高变形菌门、放线菌门、芽单胞菌门等的相对丰度,降低酸杆菌门、绿弯菌门、奇古菌门、拟杆菌门等的相对丰度;在属水平上提高Gaiella、芽单胞菌属、Pseudolabrys 的相对丰度,降低亚硝化球菌属、硝化螺菌属、红游动菌属、Gp3和Terrimonas 等的相对丰度,说明槟榔林下间作平托花生能改变土壤细菌群落特性。
槟榔林下土壤细菌结构差异主要是受速效钾、有机碳、全氮和干湿比等理化因子的影响。放线菌门、芽单胞菌门、变形菌门与速效钾、有机碳和全氮呈正相关;酸杆菌门、绿弯菌门、拟杆菌门、疣微菌门与速效钾和干湿比呈负相关;亚硝化球菌属与全碳、碱解氮呈极显著负相关;硝化螺菌属与全磷、全钾分别呈显著和极显著负相关;酸杆菌门中Gp1、Gp2 和Gp3 与pH 呈极显著或显著负相关;Gp6 与pH 呈显著正相关;Gp4 与全氮呈显著正相关;红游动菌属(属变形菌门)与全氮呈显著正相关;Gaiella(属放线菌门)、芽单胞菌属(属芽单胞菌门)和Pseudolabrys 属(属变形菌门)均与全钾呈显著正相关,表明土壤细菌影响土壤质量。
氨基酸代谢、碳水化合物代谢、膜运输和能量代谢为槟榔林下细菌群落的主要代谢功能群,间作平托花生明显改善了细菌群菌代谢功能,表现为能量代谢功能基因显著增加,苯丙素、类黄酮、甜菜红色素、吲哚生物碱等次生代谢产物的生物合成功能基因显著减少,辅助因子和维生素的代谢功能基因以及影响昼夜节律-植物的功能基因显著减少。代谢功能的变化能够反映土壤微环境的状况,为筛选良好的槟榔种植模式奠定理论基础。
猜你喜欢
经济林研究(2022年4期)2023-01-12 09:14:46
基层中医药(2021年11期)2021-06-05 06:54:38
河北果树(2020年4期)2020-11-26 06:05:18
读书文摘(2017年10期)2017-10-16 14:26:57
民族音乐(2016年1期)2016-08-28 20:02:52
新疆农垦科技(2016年2期)2016-08-21 13:50:18
现代农业(2016年3期)2016-04-14 12:35:28
土壤与作物(2015年3期)2015-12-08 00:47:01
新疆农垦科技(2014年10期)2014-02-28 19:21:12
中国现代中药(2012年6期)2012-10-30 01:38:18
