
白桦脂醇对宫颈癌小鼠自然杀伤细胞杀伤力及淋巴细胞增殖活性的影响
2022-04-07 01:58罗芳高琴费志医
中国现代医学杂志 2022年6期
罗芳,高琴,费志医
(武汉科技大学附属普仁医院,湖北武汉430081)
宫颈癌的发病率居女性恶性肿瘤的第3 位,也是引起女性恶性肿瘤患者死亡的主要原因[1]。p38是促分裂原活化的蛋白激酶(mitogen-activated protein kinase, MAPK)的同工酶之一,有研究证实其过表达对宫颈癌发生、发展有促进作用[2]。有研究显示,免疫抑制在肿瘤发生、发展中起重要作用,肿瘤免疫已成为肿瘤治疗的新思路[3]。自然杀伤(natural killer, NK)细胞可以靶向识别肿瘤细胞,并诱导肿瘤细胞坏死。有研究显示,低水平的NK 细胞与宫颈癌患者预后不良和更差的化疗反应关系密切[4-5]。白桦脂醇是一种从桦树皮中提取的三萜类化合物,具有抗炎、抗菌和抗病毒活性。有研究发现,白桦脂醇能够诱导肺癌细胞凋亡[6],对乳腺癌也有抑制作用[7],但是其在宫颈癌中的作用尚不清楚。本研究主要分析白桦脂醇调控p38 途径对宫颈癌小鼠NK 细胞杀伤力及淋巴细胞增殖活性的影响。
1 材料与方法
1.1 材料
人宫颈癌细胞系HeLa(美国ATCC 公司)。SPF级BALB/C 雌性裸鼠,4 周龄,体重(12±1)g,购自北京维通利华公司,实验动物生产许可证号:SCXK(京)2015-0001;实验动物使用许可证号:SYXK(京)2019-0010。
1.2 主要试剂及仪器
DMEM 培养基血清和抗体(美国Invitrogen 公司),CCK-8 试剂盒(上海碧云天生物技术有限公司),白桦脂醇(美国Sigma 公司),p38 通路抑制剂SB203580(美国Selleck 公司),抗体、一抗和山羊抗免疫球蛋白G(IgG)二抗(1∶1 000 稀释,#ab6721)(美国Abcam 公司),硝酸纤维素膜(美国EMD Millipore 公司),ECL 显色试剂盒(美国Thermo Fisher公司),光学显微镜(美国Nikon 公司)。
1.3 实验分组
将DMEM 培养基中处于对数生长期的HeLa 细胞重新配置密度为1×107个/mL 的溶液。所有小鼠左侧前肢皮下注射0.2 mL 细胞溶液复制宫颈癌裸鼠模型,7 d 后触摸到瘤体生成则提示模型复制成功。随机选择45 只模型复制成功的小鼠分为模型组、白桦脂醇组和白桦脂醇+SB203580 组,每组15 只。另取15 只未做处理的小鼠作为对照组。模型复制成功后第8 天,白桦脂醇组和白桦脂醇+SB203580组小鼠使用白桦脂醇灌胃,剂量100 mg/kg,1 次/d[8]。白桦脂醇+SB203580 组小鼠通过腹腔注射SB203580来抑制p38 通路,剂量50 ng/kg,2 次/周。
1.4 方法
1.4.1 肿瘤生长情况每天观察肿瘤生长情况,在第28 天用游标卡尺测量皮下肿瘤长度和宽度,计算肿瘤体积,体积=[(长度×宽度)/2]3×0.5236。模型复制成功后第28 天颈脱臼处死小鼠,取出肿瘤组织并称重。
1.4.2 免疫组织化学染色将宫颈肿瘤组织固定于10%甲醛,石蜡包埋,切成4 μm 厚的切片。将切片放在涂有0.1%聚L 赖氨酸的载玻片上,二甲苯脱蜡,100%~70%梯度乙醇逐渐水合,磷酸盐缓冲液洗涤。随后用柠檬酸钠缓冲液(pH 6.0)处理,对组织切片进行热诱导修复。在室温下,将玻片用30%过氧化氢溶液处理10 min,封闭内源性过氧化物酶。磷酸盐缓冲液冲洗后,在室温下用5%胎牛血清白蛋白封闭玻片10 min,加入anti-Ki67(1∶100 稀释)抗体,在4℃条件下孵育过夜。磷酸盐缓冲液洗涤后,将每张玻片与特定的生物素化二抗在37℃条件下孵育60 min,用3,3′-二胺苯联苯胺和苏木精复染。显微镜下观察切片,用半定量法分析。染色强度=染色评分(0~3 分)×染色范围(0~4 分)。
1.4.3 Western blotting 检测p38 蛋白的表达宫颈癌组织研磨、裂解后4℃、12 000 r/min 离心5 min,收集总蛋白。通过8% SDS-PAGE 分离每个标本中等量蛋白质(50 μg),并将其转移到硝酸纤维素膜上。在室温下,将膜浸入5%脱脂牛奶2 h,封闭非特异性抗原。然后将膜与p38、pp38 一抗在4℃条件下孵育过夜,将膜与相应的辣根过氧化物酶耦联的二抗在室温下孵育1 h。使用化学发光试剂和Quantum One软件分析灰度,计算蛋白相对表达量。
1.4.4 脾脏指数小鼠处死后立即称重,分离并取出脾脏,称重,计算脾脏与体重的比值作为脾脏指数。
1.4.5 NK细胞杀伤力在无菌环境下收集脾脏细胞悬液,用DMEM 培养基重悬并调整细胞密度为2×106个/mL 作为效细胞。然后将HeLa 细胞也调整成相同密度作为靶细胞。在96 孔板中,按照效/靶比例25∶1 加入脾脏细胞悬液180 μL 和HeLa 细胞20 μL 作为实验细胞。37℃、5%CO2环境下孵育48 h,每孔加入10 μL CCK-8 溶液孵育2 h,用酶标仪检测450 nm 波长处的光密度(optical density, OD)值。分别设置200 μL 效细胞孔及200 μL 靶细胞孔,进行上述处理后测定OD 值。NK 细胞杀伤力={1-[(OD实验-OD效)/OD靶]}×100%。
1.4.6 淋巴细胞增殖实验按照1.4.5 中方法收集脾细胞,经过密度梯度离心后抽吸淋巴细胞层,调整细胞密度为1×106个/mL。将100 μL 细胞悬浮液添加到96 孔板中,孵育48 h 后每孔加入10 μL CCK-8溶液孵育2 h。采用酶标仪检测450 nm 波长处的OD值表示淋巴细胞增殖活力。
1.5 统计学方法
数据分析采用SPSS 19 统计软件。计量资料以均数±标准差(±s)表示,比较用方差分析,进一步两两比较用SNK-q检验。P<0.05 为差异有统计学意义。
2 结果
2.1 白桦脂醇对宫颈癌的抑制作用
模型组、白桦脂醇组、白桦脂醇+SB203580 组小鼠肿瘤体积比较,经方差分析,差异有统计学意义(P<0.05)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组肿瘤体积小于模型组(P<0.05),并且白桦脂醇+SB203580 组小于白桦脂醇组(P<0.05)。见表1。
模型组、白桦脂醇组、白桦脂醇+SB203580 组小鼠肿瘤质量比较,经方差分析,差异有统计学意义(P<0.05)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组肿瘤质量小于模型组(P<0.05),并且白桦脂醇+SB203580 组小于白桦脂醇组(P<0.05)。见表1。
表1 3组小鼠肿瘤体积和质量比较 (n=15,±s)
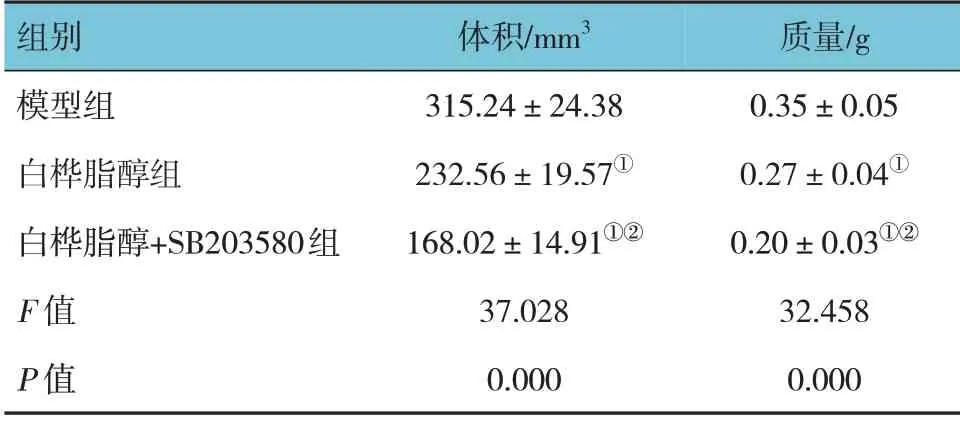
表1 3组小鼠肿瘤体积和质量比较 (n=15,±s)
注:①与模型组比较,P <0.05;②与白桦脂醇组比较,P <0.05。
组别模型组白桦脂醇组白桦脂醇+SB203580组F 值P 值体积/mm3 315.24±24.38 232.56±19.57①168.02±14.91①②37.028 0.000质量/g 0.35±0.05 0.27±0.04①0.20±0.03①②32.458 0.000
2.2 白桦脂醇对宫颈癌细胞增殖的影响
对照组、模型组、白桦脂醇组、白桦脂醇+SB203580 组免疫组织化学染色评分分别为(1.04±0.08)分、(9.23±0.86)分、(5.49±0.57)分和(3.15±0.24)分,经方差分析,差异有统计学意义(F=49.742,P=0.000)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组免疫组织化学染色评分低于模型组(P<0.05),并且白桦脂醇+SB203580 组低于白桦脂醇组(P<0.05)。见图1。
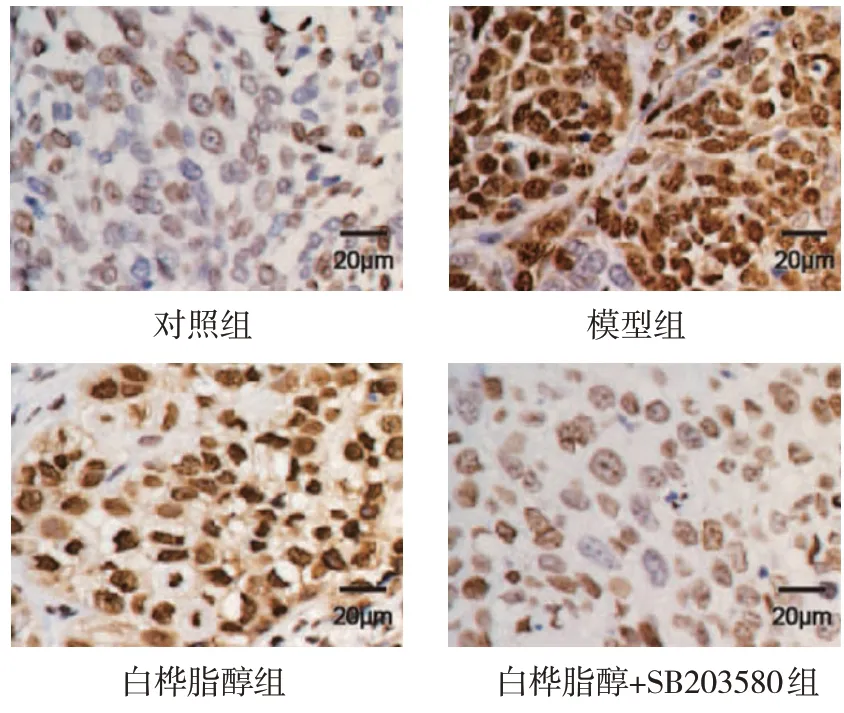
图1 白桦脂醇对宫颈癌组织中Ki67蛋白的影响(免疫组织化学染色)
2.3 白桦脂醇对宫颈癌组织中p38 蛋白磷酸化的影响
对照组、模型组、白桦脂醇组、白桦脂醇+SB203580 组p38 蛋白相对表达量分别为(0.24±0.02)、(2.28±0.19)、(1.27±0.10)和(0.42±0.04),经方差分析,差异有统计学意义(F=76.214,P=0.000)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组p38 蛋白相对表达量低于模型组(P<0.05),并且白桦脂醇+SB203580 组低于白桦脂醇组(P<0.05)。见图2。
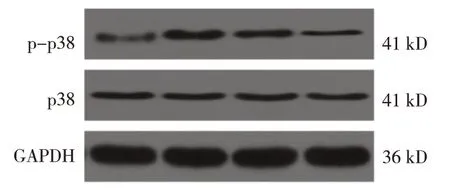
图2 白桦脂醇对宫颈癌组织中p38蛋白磷酸化的影响
2.4 白桦脂醇对宫颈癌小鼠脾脏指数的影响
对照组、模型组、白桦脂醇组、白桦脂醇+SB203580 组小鼠脾脏指数分别为(9.02±1.15)mg/g、(4.76±0.58)mg/g、(6.61±0.77)mg/g 和(8.25±0.92)mg/g,经方差分析,差异有统计学意义(F=36.527,P=0.000)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组脾脏指数高于模型组(P<0.05),并且白桦脂醇+SB203580 组高于白桦脂醇组(P<0.05)。
2.5 白桦脂醇对宫颈癌小鼠NK 细胞杀伤力和淋巴细胞增殖活性的影响
对照组、模型组、白桦脂醇组、白桦脂醇+SB203580 组小鼠NK 细胞杀伤力比较,经方差分析,差异有统计学意义(P<0.05)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组NK 细胞杀伤力高于模型组(P<0.05),并且白桦脂醇+SB203580 组高于白桦脂醇组(P<0.05)。见表2。
表2 4组小鼠NK细胞杀伤力和淋巴细胞增殖活性比较(n=15,±s)
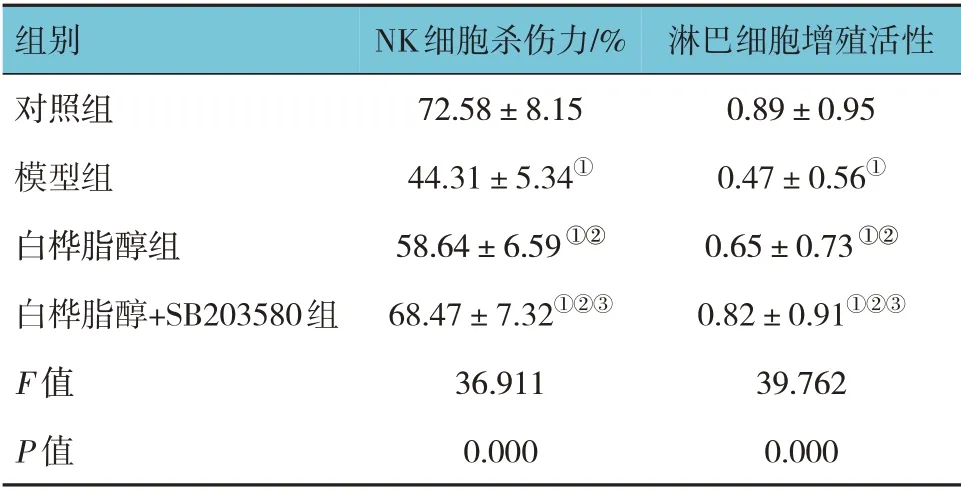
表2 4组小鼠NK细胞杀伤力和淋巴细胞增殖活性比较(n=15,±s)
注:①对照组比较,P <0.05;②与模型组比较,P <0.05;③与白桦脂醇组比较,P <0.05。
淋巴细胞增殖活性0.89±0.95 0.47±0.56①0.65±0.73①②0.82±0.91①②③39.762 0.000组别对照组模型组白桦脂醇组白桦脂醇+SB203580组F 值P 值NK细胞杀伤力/%72.58±8.15 44.31±5.34①58.64±6.59①②68.47±7.32①②③36.911 0.000
对照组、模型组、白桦脂醇组、白桦脂醇+SB203580 组小鼠淋巴细胞增殖活性比较,经方差分析,差异有统计学意义(P<0.05)。进一步两两比较结果:白桦脂醇组和白桦脂醇+SB203580 组淋巴细胞增殖活性高于模型组(P<0.05),并且白桦脂醇+SB203580 组高于白桦脂醇组(P<0.05)。见表2。
3 讨论
宫颈癌是女性常见恶性肿瘤,治疗方法包括根治性子宫切除术、化疗、放疗和同步放化疗。然而宫颈癌复发率为10%~50%,5年生存率约为65%[9]。近年来,中药治疗肿瘤取得了重大进展,寻找新的抑制宫颈癌的药物具有重要的现实意义。
白桦脂醇是一种五环羽扇豆型三萜类的天然活性化合物,存在于桦树的外皮中,具有抗菌、抗寄生虫、抗病毒、抗炎、抗蛇毒血清、内脏保护、抗糖尿病、抗动脉粥样硬化等作用[10-11]。近年来研究发现,白桦脂醇具有广泛的抗肿瘤活性,如通过抑制NFκB/c-Myc 通路,抑制乳腺癌细胞有氧糖酵解[12]。也有研究显示,白桦脂醇可通过调控Parkin 依赖性线粒体自噬,提高多药耐药肿瘤细胞对化疗的敏感性[13]。
为进一步分析白桦脂醇抑制宫颈癌的机制,本研究从p38 通路和免疫抑制方面分析了作用机制。p38 通路是与宫颈癌关系密切的通路,p38 蛋白磷酸化后,会调控一系列基因转录,促进细胞增殖和抗凋亡等[14]。本研究结果显示,白桦脂醇能够显著抑制宫颈癌组织中p38 蛋白磷酸化水平。此外,采用SB203580 抑制p38 通路后,会进一步提高白桦脂醇对宫颈癌细胞增殖的抑制作用。董蕊等[14]的研究表明,白桦脂醇可抑制p38 通路的激活,抑制黑色素瘤细胞合成黑色素。也有研究显示,白桦脂醇可通过调节p38 MAPK 通路,减弱T2 毒素引起的胸腺氧化应激反应[15]。结合既往研究和本研究结果,提示白桦脂醇抑制宫颈癌的作用机制与p38 通路密不可分,白桦脂醇可通过抑制p38 通路中p38 蛋白磷酸化来抑制宫颈癌细胞增殖。
白桦脂醇也具有关键的免疫调控作用,免疫抑制也是影响肿瘤增殖和转移的重要因素。NK 细胞是在骨髓或者脾脏中成熟的一种免疫细胞,可识别肿瘤细胞并分泌穿孔素等引起肿瘤细胞溶解[16]。宫颈癌患者体内NK 细胞杀伤力降低,并且宫颈癌组织微环境中浸润的NK 细胞水平和活性也降低[17]。有研究显示,活化的NK 细胞通过p38 信号杀死肝星状细胞[18]。脾脏是T 淋巴细胞成熟和分化的重要场所,有研究显示宫颈癌小鼠模型出现脾脏萎缩和免疫抑制[19]。宫颈癌患者体内CD3+和CD4+细胞比例均显著降低,功能也受到抑制,并使细胞逃离T 细胞的识别[20]。p38 可通过调控体内免疫抑制来抑制免疫炎症反应[21]。本研究结果表明,宫颈癌小鼠模型NK 细胞杀伤力降低,脾脏指数和淋巴细胞增殖活性也受到抑制。而白桦脂醇可显著提高脾脏指数和淋巴细胞增殖活性,并提高NK 细胞杀伤力,抑制p38 通路会进一步提高白桦脂醇对细胞免疫的促进作用。有研究显示,白桦脂醇提高成熟CD4+和CD8+细胞的百分比和绝对计数,增加其在脾脏中的细胞数,从而调节免疫细胞亚群[8]。本研究结果提示,白桦脂醇可通过抑制p38 通路,提高NK 细胞杀伤力和淋巴细胞增殖活性。
综上所述,在宫颈癌小鼠模型中,白桦脂醇通过抑制p38 通路,抑制宫颈癌细胞增殖;并且提高NK 细胞杀伤力和淋巴细胞增殖活性,提高机体免疫水平。但是白桦脂醇在宫颈癌中调控免疫作用的分子机制仍需要临床研究证实。
猜你喜欢
奥秘(2021年1期)2021-03-15
保健与生活(2020年5期)2020-03-20
保健文汇(2020年12期)2020-02-02
文苑(2019年20期)2019-11-20
中国保健营养(2019年7期)2019-10-21
文苑·经典美文(2019年10期)2019-10-16
作文周刊·小学二年级版(2019年20期)2019-07-03
中学生天地(B版)(2017年4期)2017-04-25
腹腔镜外科杂志(2016年11期)2016-06-01
华人时刊(2016年16期)2016-04-05
