
基于iTRAQ 技术分析镉胁迫下泽兰实蝇雄性附腺差异蛋白的表达*
2022-04-07 12:56:32桂华敏鲁武锋李丽芳吴国星
云南农业大学学报(自然科学) 2022年2期
桂华敏,鲁武锋,李丽芳,高 平,吴国星,高 熹
(云南农业大学 植物保护学院,云南 昆明 650201)
紫茎泽兰(Ageratina adenophoraSpreng.)是泽兰属多年生草本或亚灌木植物,是一种入侵性杂草,分布于中国多个省区[1]。泽兰实蝇(Procecidochares utilisStone)属双翅目(Diptera)实蝇科(Tephritidae),是恶性杂草紫茎泽兰的重要专食性天敌,在紫茎泽兰防治中起着重要作用。雌性泽兰实蝇把卵产在寄主紫茎泽兰的生长点上,通过幼虫产生的虫瘿来控制紫茎泽兰开花、结实及扩散[2-5]。泽兰实蝇可有效控制紫茎泽兰的生长及发育,并能抑制其光合作用及生物量的增加和分配[6-9],通过影响其生理作用从而有效抑制其蔓延危害。
随着经济快速发展,环境中的重金属镉污染越来越严重,如今镉已成为中国污染面积最大的重金属[10]。相关研究表明:泽兰实蝇对紫茎泽兰的寄生率与离矿中心距离成反比,说明在镉污染区范围内,泽兰实蝇对紫茎泽兰的防控作用低。昆虫的雄性附腺能产生生殖因子,具有交配效应功能。染镉的泽兰实蝇交配后,出现寿命短、交配能力下降和含卵量低等问题,且雌虫与染镉的雄虫交配后出现再次交配率显著上升的情况,表明重金属镉抑制和干扰泽兰实蝇的交配效应[11]。此外,镉胁迫对泽兰实蝇雄性附腺的长度和宽度均有显著影响,对雄性附腺基因的表达也有影响[11]。在生物体内,不同蛋白质相互协调行使其生物学行为,通过 Pathway 分析的显著性富集能确定差异蛋白参与的最主要生化代谢途径和信号转导途径;利用 WEGO 软件对差异蛋白的主要功能类别进行 GO 注释分析,可确定差异蛋白行使的主要生物学功能。因此,本研究以泽兰实蝇雄性附腺蛋白质为研究对象,利用iTRAQ技术分离并鉴定受镉胁迫后差异表达的泽兰实蝇雄性附腺蛋白质,分析其对重金属镉的响应机制,为探究重金属镉对泽兰实蝇雄性附腺的毒性效应及其分子机制提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 泽兰实蝇种群的建立
在塑料大棚内大批量种植紫茎泽兰以建立泽兰实蝇种群,以扦插的方式将未经重金属污染的野生紫茎泽兰扦插,种植在养虫笼内。待紫茎泽兰成活后接入野生泽兰实蝇成虫,以便让其在紫茎泽兰上产卵。
1.1.2 样品制备
将实验室中的泽兰实蝇分为对照组(CK)和重金属镉处理组(100 mg/kg)[11]。镉处理组每个月向紫茎泽兰浇灌1 次重金属镉,以清水作为对照,浇水时一次浇透,浇到盆底漏水为止。当重金属镉初次浇灌1 周后,陆续不间断放入已交配但未经镉污染的雌性泽兰实蝇,用于产卵。卵孵化成幼虫后寄生于紫茎泽兰,将形成透明窗口的虫瘿剪下并放入扎好透气孔的广口瓶内;将广口瓶置于温度24~26 ℃、湿度70%的恒温恒压光照培养箱内,随时观察泽兰实蝇的羽化情况并做好标记[11]。
1.1.3 雄性附腺蛋白质的提取
当有成虫羽化时,分别收集当天交配过的染镉和未染镉的泽兰实蝇雄虫置于PBS 中解剖得到雄性附腺,每组解剖300 头。
1.2 试验方法
1.2.1 蛋白提取和样品处理
每个样品取适量进行提取试验。向样品中加入蛋白裂解液[7 mol/L 尿素、2 mol/L 硫脲、4%十二烷基苯磺酸钠(SDS)、40 mmol/L Tris-HCl、pH 为8.5 的1 mmol/L 苯甲基磺酰氟(PMSF)和2 mmol/L 乙二胺四乙酸(EDTA)],混匀后冰上孵育5 min,加入终浓度为l0 mmol/L 的二硫苏糖醇(DTT),冰浴超声15 min,然后13 000 r/min、4 ℃离心20 min,取上清装入新的离心管中;向离心管中加入4 倍体积的冷丙酮,在-20 ℃静置过夜。离心收集蛋白沉淀,空气中晾干。加入8 mol/L 尿素和100 mmol/L TEAB (pH 8.0)溶液重新溶解蛋白,加入DTT 至终浓度为10 mmol/L,56 ℃水浴30 min 进行还原反应;加入碘乙酰胺(IAM)至终浓度为55 mmol/L,暗处室温放置30 min 进行烷基化反应。用Bradford 法测定可溶性蛋白浓度[12]。
1.2.2 iTRAQ 标记、HPLC 分离及质谱鉴定
将蛋白样品进行酶解处理,酶解的肽段用同位素标记相对和绝对定量技术(iTRAO)标记;运用Ultimate 3000 HPLC 系统(Thermo DINOEX,USA)对肽段样品进行分级分离,色谱柱为Durashell C18 柱(5 μm,100 Å,4.6 mm×250 mm);使用TripleTOF 5600+液质联用系统(SCIEX,USA)对分离的肽段进行质谱检测,用ProteomicsTools 软件[13]将格式转换成“.mgf”后进行Mascot 查库鉴定。肽段所代表的蛋白量依据其标记后的同位素标签进行定量,统计处理组和对照组样品的蛋白量,最终获得差异表达的蛋白。
1.2.3 蛋白质鉴定及生物信息学分析
质谱数据使用ProteinPilotTMV4.5 (AB Sciex 5600 plus)数据库搜索[14]。对ProteinPilot 的鉴定结果进一步过滤,若 unused score≥1.3 (即可信度水平在 95%以上),即可认为至少包含1个 unique肽段的蛋白为可信蛋白。利用重复样品间两两比值的均值经过中位数归一化后作为待比较样品的差异倍数;利用重复样品间两两比较单样本 Student’st检验P值的最小值作为待比较样品间的显著性差异检验P值;根据差异倍数和P值筛选差异蛋白。当差异倍数达到 1.5 倍及以上(即up_regulate≥1.5 和down_regulate≤0.67)且经过显著性统计检验其P≤0.05 时,视为显著差异蛋白。利用 WEGO 软件对鉴定出差异蛋白的主要功能类别进行 GO 注释分析,通过 Pathway 分析确定蛋白质参与的最主要途径;为充分挖掘定量数据包含的生物信息学信息,将鉴定出的差异蛋白进行GO 和KEGG 数据库比对。
2 结果与分析
2.1 泽兰实蝇雄性附腺总蛋白及差异蛋白的鉴定
脱盐后的泽兰实蝇雄性附腺样品经质谱分析及Mascot 查库,共鉴定到3 661个蛋白,最终有统计学意义的差异蛋白质共 58个(P<0.05),表达上调的蛋白38个,表达下调的蛋白20个(图1)。在这些差异表达的蛋白中,表达量上调和下调倍数前10 的蛋白见表1。

表1 泽兰实蝇雄性附腺镉诱导组与对照组排列前10 的差异蛋白Tab.1 The top ten differences in protein analysis on the male accessory gland in Procecidochares utilis with cadmium induced and control group
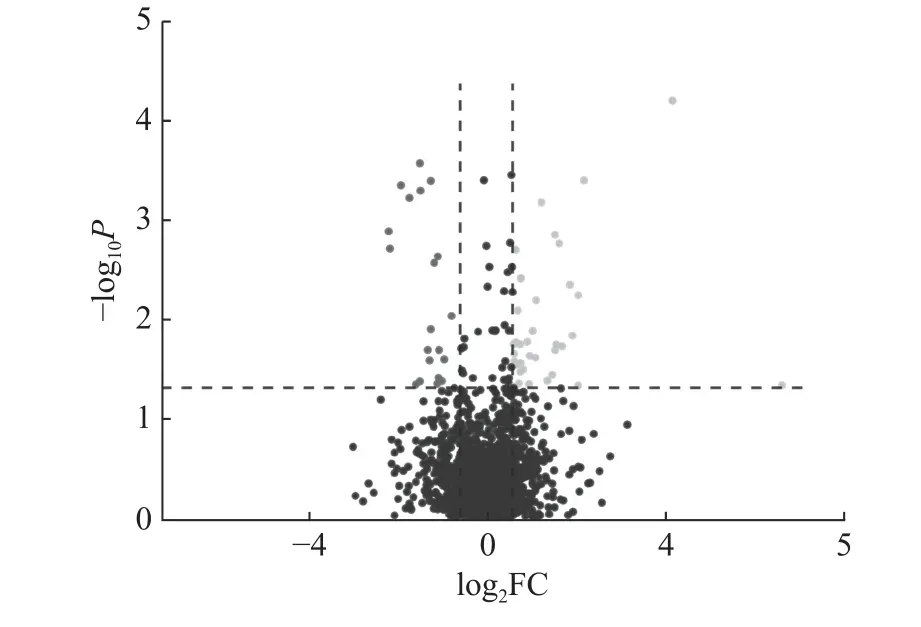
图1 染镉和未染镉组中差异蛋白占总定量蛋白比例的火山分布图Fig.1 Volcano distribution map of the proportion of different proteins account for total quantitative proteins
2.2 差异蛋白的GO 注释
由图2 可知:染镉诱导后的差异蛋白在生物学途径中主要参与细胞过程和代谢途径。从细胞组分中可看出:染镉诱导后产生变化的差异蛋白功能主要为细胞、细胞器部分、细胞器和大分子复合体;而在差异蛋白的分子功能注释中可看出:染镉诱导后产生变化的差异蛋白主要功能为催化活性、结合、结构分子活性和分子转换活性,其中以结合和催化类蛋白最多,分别占功能差异蛋白的48.27%和 46.55%。

图2 差异蛋白的GO 注释Fig.2 GO functional annotation for differential protein
2.3 差异蛋白的Pathway 代谢通路注释
由图3 可知:排名前10 的Pathway 代谢功能类型在所有上调和下调差异蛋白质中有所不同。排名前3 的上调差异蛋白Pathway 功能为代谢途径、微生物在不同环境中的代谢和核糖体,排名前3 的下调差异蛋白Pathway 功能为代谢途径、溶酶体和致病性大肠杆菌感染;且与下调差异蛋白注释结果相比,代谢途径在上调差异蛋白质功能注释结果中的比例明显上升。
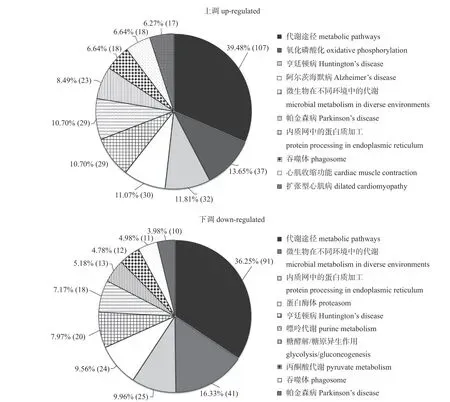
图3 染镉和未染镉泽兰实蝇雄性附腺中上调/下调差异蛋白质 Pathway 代谢通路Fig.3 Pathway of up-regulated/down-regulated proteins in the accessory glands of P.utilis with cadmium infected and non cadmium infected
2.4 差异蛋白的富集分析
2.4.1 差异蛋白的GO 富集分析
由图4 可知:差异表达的蛋白在生物学过程中主要富集于分解代谢过程、有机酸代谢过程、羧酸代谢过程和酮酸代谢过程等,在分子功能方面主要集中富集于金属离子结合、离子结合和阳离子结合过程,在细胞组分中主要富集于细胞内部分和细胞质。

图4 差异蛋白的 GO 富集分析Fig.4 GO enrichment analysis of different proteins
2.4.2 差异蛋白的KEGG 富集分析
由图5 可知:差异蛋白主要参与的途径包括代谢途径、微生物在不同环境中的代谢、酪氨酸代谢和核糖体,说明这些途径在镉胁迫泽兰实蝇中起重要作用。其中,上调的差异蛋白18个,包括过氧化氢酶、酚氧化酶和胞质附子水合酶等;下调的差异蛋白5个,包括谷胱甘肽S-转移酶、脂肪酸合酶和β-半乳糖苷酶等(表2)。

表2 参与代谢途径数量最多的差异蛋白Tab.2 Differential proteins with the largest number of metabolic pathways

图5 差异蛋白 Pathway 富集分析气泡图Fig.5 Differential protein pathway enrichment analysis bubble chart
3 讨论
环境中的重金属主要通过昆虫的呼吸和取食及外表皮渗透等物理作用直接进入昆虫体内;也能通过改变植物化学物质成分间接影响昆虫生物习性和昆虫种群增长[15-18]。如在重金属镉的胁迫下,能引起白纹伊蚊幼虫、甜菜夜蛾和斜纹夜蛾等昆虫生理生化的一系列改变[19-21]。实验室前期研究发现:经试验处理的Cd2+、Pb2+或Zn2+均可通过紫茎泽兰传递并累积到泽兰实蝇老熟幼虫体内,引起老熟幼虫一系列生理变化[22]。镉胁迫后,泽兰实蝇雄性附腺的长度和宽度均发生明显变化,且不同浓度镉胁迫下泽兰实蝇雄性附腺发生不同程度的变化,镉胁迫同样对雄性附腺基因的表达产生影响,造成150 条基因差异表达[11]。基因水平上的差异最终将反映在蛋白质水平上,所以本研究利用iTRAQ 技术分析镉胁迫下泽兰实蝇雄性附腺差异蛋白的表达。
本研究共鉴定出3 661个蛋白,具有定量信息差异的蛋白58个,其中上调的蛋白38个,下调的蛋白20个。在这些差异表达的蛋白中,唯一肽段最多的上调蛋白有:丙酮酸羧化酶、嘌呤霉素敏感氨肽酶、过氧化氢酶(CAT)和Rab GDP 解离抑制剂α 等。其中,限速酶丙酮酸羧化酶存在于线粒体中,催化不可逆的反应;嘌呤霉素敏感的氨肽酶是一种大脑分泌的酶;CAT 是一类广泛存在于动物、植物和微生物体内的末端氧化酶,是生物防御系统的关键酶之一,它能催化细胞内过氧化氢分解,防止过氧化;也有研究表明:CAT 能将蚕在代谢过程中产生的有毒过氧化氢分解成水和分子氧从而达到解毒作用[23-24];Rab GDP 解离抑制剂α 是涉及神经传导的Rab 小G 蛋白的调节因子[25]。在差异表达的蛋白中,下调蛋白有蛋白质二硫键异构酶(PDI)、可溶性异柠檬酸脱氢酶(IDH)、脂肪酸合酶和谷胱甘肽S-转移酶(GST)等。其中,PDI 是1个多功能的应激蛋白,对病毒和细菌的侵染具有防御作用;IDH 是三羧酸循环的关键酶,在三羧酸循环和乙醛酸旁路的碳通量分配中起关键的调控作用;脂肪酸合酶是脂肪酸合成的关键限速酶,可影响机体内脂肪组织的合成代谢和分解代谢[26];GSTs是昆虫中常见的多功能氧化酶系,在昆虫对外源物质解毒代谢过程中发生作用[27]。当生物体处于逆境时,GST 常发挥二相代谢酶和抗氧化酶的双重功能,保护生物体免受逆境损害[28]。
研究发现:GST 在苹果蠹蛾对吡虫啉的解毒代谢中起主导作用[29];不同浓度镉处理后,GST 活性在棕尾别麻蝇幼虫和成虫体内均受到抑制[30];经Cd2+和Cu2+处理后,棕尾别麻蝇幼虫体内超氧化物歧化酶 (SOD)和CAT 活性受抑制,且抑制程度随处理浓度的升高而加强[31]。本研究发现:镉胁迫下CAT 表达量上调,这与低浓度Cd2+处理下中华稻蝗的 GST 活性被激活[19]类似,而重金属对昆虫抗氧化酶系统的影响因昆虫种类和重金属种类而异[31],故推测本研究中 CAT活性升高可能是昆虫为抵御由重金属引起的氧化损伤和增强解毒代谢而做出的应激反应[32]。镉能在体内富集,被富集后能显著影响泽兰实蝇的生长发育和化学防御;此外,镉会对泽兰实蝇的生殖功能造成一定的损坏,导致泽兰实蝇生殖力减弱[11],低浓度重金属对泽兰实蝇幼虫的影响小,高浓度则影响大,泽兰实蝇幼虫对重金属具有一定的抵御作用[22],故推测泽兰实蝇成虫也能利用自身的解毒或避毒机制抵御重金属危害。昆虫可通过抗氧化酶系统清除过量活性氧以减少镉对机体的损害[33],除消化道、马氏管、脂肪体和体壁外,昆虫的生殖腺也具有储存过量重金属的功能[22],由此分析泽兰实蝇雄性附腺可通过调控CAT 和GST 对重金属进行代谢解毒,从而延缓并减弱重金属造成的毒性损伤。
本研究对差异蛋白的GO 显著性分析发现:差异表达的蛋白在生物学过程中主要富集于分解代谢过程、有机酸代谢过程、羧酸代谢过程和酮酸代谢过程,这些代谢过程在生物学过程中起着关键作用;在分子功能方面主要集中富集于金属离子结合、离子结合和阳离子结合过程;在细胞组分中主要富集于细胞内部分和细胞质,说明差异表达的蛋白主要在细胞内发挥作用。通过 Pathway 显著性富集确定差异蛋白主要参与的途径有代谢途径、微生物在不同环境中的代谢、酪氨酸代谢和核糖体,说明这些途径在镉胁迫泽兰实蝇中起重要作用。在这些代谢途径中,上调蛋白18个,包括过氧化氢酶、酚氧化酶和胞质附子水合酶等,其中,酚氧化酶在昆虫免疫防御以及生理发育过程中起到不可或缺的作用[34],胞质附子水合酶是参与抗氧化反应的硫氧还原蛋白过氧化物酶;下调蛋白5个,包括谷胱甘肽S-转移酶、脂肪酸合酶和β-半乳糖苷酶等,其中,脂肪酸合酶在能量运输和储存、细胞结构和提供激素合成的中间物等多个方面发挥关键作用。
4 结论
本研究明确了镉污染可改变泽兰实蝇雄性附腺的蛋白质组,鉴定了镉胁迫泽兰实蝇雄性附腺后的差异蛋白,为进一步揭示重金属胁迫对泽兰实蝇雄性附腺蛋白质组的响应原理及生殖毒性损伤机理奠定了基础,为今后的基因克隆和功能研究提供了基础数据。
猜你喜欢
浙江农林大学学报(2022年6期)2022-12-07 03:08:04
云南化工(2021年7期)2021-12-21 07:27:34
中成药(2019年12期)2020-01-04 02:02:56
中成药(2018年12期)2018-12-29 12:25:56
中成药(2018年6期)2018-07-11 03:01:30
蔬菜(2016年8期)2016-10-10 06:49:12
湖南农业(2016年3期)2016-06-05 09:37:36
湖南农业(2016年3期)2016-06-05 09:37:35
湖南农业(2016年3期)2016-06-05 09:37:34
湖南农业(2015年5期)2015-02-26 07:32:26
