
林窗尺度对马尾松人工林林下灌草层优势种生态位的早期影响
2022-04-06 06:29刘江丽刘俊杰陈小红李贤伟
西北植物学报 2022年2期
刘江丽,吕 倩,刘俊杰,陈小红,2,范 川,2,李贤伟,2,3*
(1 四川农业大学 林学院,成都 611130;2 长江上游森林资源保育与生态安全国家林业局重点实验室,成都 611130;3 长江上游林业生态工程四川省重点实验室,成都 611130)
生态位可表征生态系统中物种间以及物种与环境之间的关系[1-2]。在生态系统中利用不同资源、生存在不同空间的物种因不存在资源和空间的争夺而能使群落得以稳定,当不同物种利用相同的资源和空间时,根据优胜劣汰原则,最终会形成以优势物种为主的稳定群落[3-4]。生态位宽度和生态位重叠作为生态位的两个主要指标,其均能在一定程度上反映群落结构及物种多样性,其中生态位宽度反映物种对资源和空间的利用情况,生态位重叠能反映物种间竞争的激烈程度及群落稳定程度[5-6]。目前已有的生态位相关研究主要集中在生态位季节性变化[7]、土壤种类对物种生态位的影响[8]、土壤细菌生态位[9]、气候变化对生态位的影响[10]及生态位模型[11]等方面,而开窗尺度对林下物种生态位影响的相关研究较少。
马尾松(Pinusmassoniana)具有耐贫瘠、速生、适应性强等特点,是中国主要造林树种之一,其人工林是中国南方面积最大的人工林[12]。人工林大多存在土壤肥力下降、生物多样性较低等问题,极大地降低了人工林生态系统的生态价值和经济价值[13]。因管理不善,长江流域低山丘陵区马尾松人工林普遍存在生长较差,林下植被群落结构简单等生态问题。目前马尾松人工林是中国南方退化人工林中面积最大的林分类型之一[14]。如何改善人工林林分结构及提高其生态效益,已成为恢复生态学研究的重要内容[15-16]。
林窗对林地更新发挥着重要作用[17]。采伐林窗后林下光照增多,林地裸露面积增大,直接导致地表温度升高,水分含量减少,间接影响林下植被的物种组成[18]、群落结构和生态服务功能[19-20]。不同大小的林窗对林下植被群落的影响程度不同[21],研究林窗尺度对林下植被更新的影响,对深入探索人工林近自然经营具有重要意义。前人研究主要集中于林窗对植物地下资源分配[22]、土壤微生物多样性[23]、植被更新[24]、植被初级生产量[25]和林分生态服务功能的影响[26]等,而关于林窗大小如何影响植物群落分布及优势种生态位的相关研究较少。
本研究以45 a生马尾松过熟林为研究对象,探究4种不同开窗尺度对马尾松林下植被组成及优势种生态位的影响,以期调整其林下群落结构,促进其充分发挥应有的生态效益和经济效益,为马尾松人工林近自然经营提供理论参考。
1 研究区概况和研究方法
1.1 研究区概况
本实验研究区位于四川省华蓥市东方红林场(30°07′ N~30°28′ N,106°37′ E~106°54′ E),海拔206.7~1 704.1 m,地处川东低山区。该地区属中亚热带湿润季风气候,具有气候温和、温差较大、雨量充沛、降雨不均等特点[27]。研究区多年平均气温17.2 ℃,平均降雨量1 087.84 mm[28]。土壤为黄壤,土层较瘠薄。未采取任何经营措施的马尾松林龄为45年,郁闭度0.6,平均胸径22 cm,平均树高15 m,林下灌木层主要为樟(Cinnamomumcamphora)、毛桐(Mallotusbarbatus)和枹栎(Quercusserrata),草本层主要覆盖蕨类。
1.2 样地设置及调查
2017年8月,在东方红林场选择海拔、坡度、坡向、坡位、林分密度及郁闭度等条件基本一致、经营历史相同的45 a生马尾松人工林,该林分生态功能处于退化状态,生产功能处于极低效状态,采用完全随机设计的方法设置50 m2(A)、100 m2(B)、200 m2(C)、667 m2(D)的4种椭圆林窗,林窗设于距离林缘10 m以上的林内,每一种处理重复3次,另设置3个20 m×20 m对照林分样地(CK),共设置15个研究样地,各样地间设有5 m缓冲带。林窗大小由激光测距仪测定,并对需采伐马尾松进行测量,计算出株数和蓄积量,林窗边界采用有色丝带进行标记。林窗内采伐的马尾松、毛竹(Phyllostachysedulis)、树干及枝条等残体全部移出林窗,林窗内植物再生主要依靠种子和土壤种子库。马尾松人工林样地概况见表1。
2018年10月,对样地中林下植物进行调查,采用五点取样法在每个样地布设5个3 m×3 m的样方作为灌木样方,再在样地内随机设置10个1 m×1 m的样方作为草本样方[29]。记录每个灌草样方中各种植物的种名、株数/丛数、冠幅、盖度等[30]。
1.3 数据处理与计算
1.3.1 物种丰富度指数计算物种丰富度采用Patrick物种丰富度指数(R)[31],计算公式如下:
R=(S-1)/lnN
式中:S为样方中出现的物种数,N为物种总数量。
1.3.2 植物重要值计算利用样地调查数据,计算出现在各样地灌草层中的物种的重要值[27](IV)。计算公式如下:
IV=(相对密度+相对频度+相对盖度)/3
1.3.3 优势种生态位宽度计算生态位宽度采用Shannon-Wiener生态位宽度指数[32],计算公式如下:
式中Bi为i物种的生态位宽度;Pij=nij/Ni,式中nij为物种i在资源j上的重要值,Ni表示物种i在所有资源上的重要值之和。Pij为物种i在资源j上的重要值占该种在所有资源上重要值之和的比例,r为资源位总位数,j为不同改造模式[33]。
1.3.4 优势种生态位重叠计算生态位重叠采用Levins[34]生态位重叠公式,计算公式如下:
式中Lih为物种i重叠物种h的生态位重叠指数;Lhi为物种h重叠物种i的生态位重叠指数;Pij、Ni和nij的定义及计算方法同上;B(L)为Levins生态位宽度指数,B(L)i和B(L)h的值域为[1/r,1];Lih和Lhi的值域为[0,1][34]。
采用Excel 2019和SPSS 20.0软件统计调查数据、计算相关指标及制图,利用方差分析(One-way ANOVA)检测不同尺度林窗下的物种丰富度指数间的差异显著性。优势种重要值和生态位宽度关系采用Pearson相关分析。
2 结果和分析
2.1 林窗尺度对马尾松人工林灌草层物种数量的影响
由图1可见,除B林窗灌木层植物种数与CK差异不显著,其余林窗灌木层物种数均显著高于CK(P<0.05)。CK、A、B、C和D林窗灌木层物种数分别为21种、28种、20种、35种和29种。各林窗草本层物种数均显著高于CK(P<0.05),除A、B林窗草本层物种数之间差异不显著,其余处理草本层物种数两两之间差异显著(P<0.05),CK、A、B、C和D林窗草本层物种数分别为6种、17种、16种、20种和11种,表明开窗增加了马尾松人工林灌草层植物种类。
2.2 林窗尺度对马尾松人工林灌草层物种丰富度的影响
由图2可知,各林窗灌草层的物种丰富度指数较CK均显著增加(P<0.05),各林窗灌草层物种丰富度指数两两之间均存在显著差异(P<0.05)。
A、B、C和D林窗灌木层物种丰富度指数分别为CK灌木层的1.2、1.1、1.5和1.3倍,A、B、C和D林窗草本层物种丰富度指数分别为CK草本层的2.3、2.5、2.6和1.6倍,C林窗灌草层的物种丰富度指数均为最大。
2.3 林窗尺度对马尾松人工林灌草层重要值及耐荫性的影响
由表2可知,CK林下的21种灌木隶属于15科18属;A林窗的28种灌木隶属于18科20属;B林窗的20种灌木隶属于14科15属;C林窗的35种灌木隶属于21科29属;D林窗的29种灌木隶属于16科21属。由表3可得,CK林下的6种草本隶属于5科6属;A林窗的17种草本隶属于10科17属;B林窗的16种草本隶属于10科16属;C林窗的20种草本隶属于14科20属;D林窗的11种草本隶属于5科10属。CK灌木层优势种为地桃花(Urenalobata)(7.7)、樟(15.1)、楤木(Araliachinensis)(7.0)、九节龙(Ardisiapusilla)(21.6)和柳杉(Cryptomeriajaponicavar.sinensis)(10.9),草本层有金星蕨(Parathelypterisglanduligera)(12.0)、蕨(Pteridiumaquilinumvar.latiusculum)(21.3)、芒(Miscanthussinensis)(16.2)等6个优势种;A林窗灌木层优势种有白花泡桐(Paulowniafortunei)(6.2)、樟(20.4)、楤木(13.9)和地桃花(7.6),草本层共有荩草(Arthraxonhispidus)(7.7)、蕨(16.2)、芒(7.5)和酢浆草(Oxaliscorniculata)(9.1)等6个优势种;B林窗灌木层优势种有菝葜(Smilaxchina)(7.7)、楤木(6.2)、地果(Ficustikoua)(13.6)、枹栎(7.0)、高粱泡(Rubuslambertianus)(6.1)等7种,草本层有狗脊(Woodwardiajaponica)(7.6)、金星蕨(8.6)、荩草(9.7)和芒(5.2)等8个优势种;C林窗灌木层优势种有紫麻(Oreocnidefrutescens)(6.0)、紫金牛(Ardisiajaponica)(7.2)、毛桐(Mallotusbarbatus)(9.3)和地桃花(10.2)等6种,草本层有淡竹叶(Lophatherumgracile)(10.4)、狗脊(6.9)、华南鳞盖蕨(Microlepiahancei)(10.1)等7个优势种;D林窗灌木层优势种有楤木(23.3)、九节龙(13.6)、梨叶悬钩子(Rubuspirifolius)(5.9)和樟(13.4),草本层共有荩草(9.1)、蕨(8.2)和芒(10.7)等5个优势种。就耐荫性而言,除A、B林窗灌木层喜光植物种类略少于CK,其余林窗下灌木层喜光植物种类均多于CK,且C林窗灌木层喜光植物种类最多有13种,而D林窗灌木层喜光植物种类略少于C林窗,可能的原因是开窗后光照增加导致喜光植物增多,而随林窗尺度增大,喜光植物间愈加激烈的竞争[35]形成了适宜耐荫植物生长的环境,此时耐荫植物逐渐取代喜光植物的优势地位。各林窗下草本层喜光植物种类均多于CK,喜光植物种类随林窗尺度增大而减少,C林窗下喜光植物种类数较多有5种,表明开窗有利于促进喜光植物的生长,且C林窗促进作用最明显。
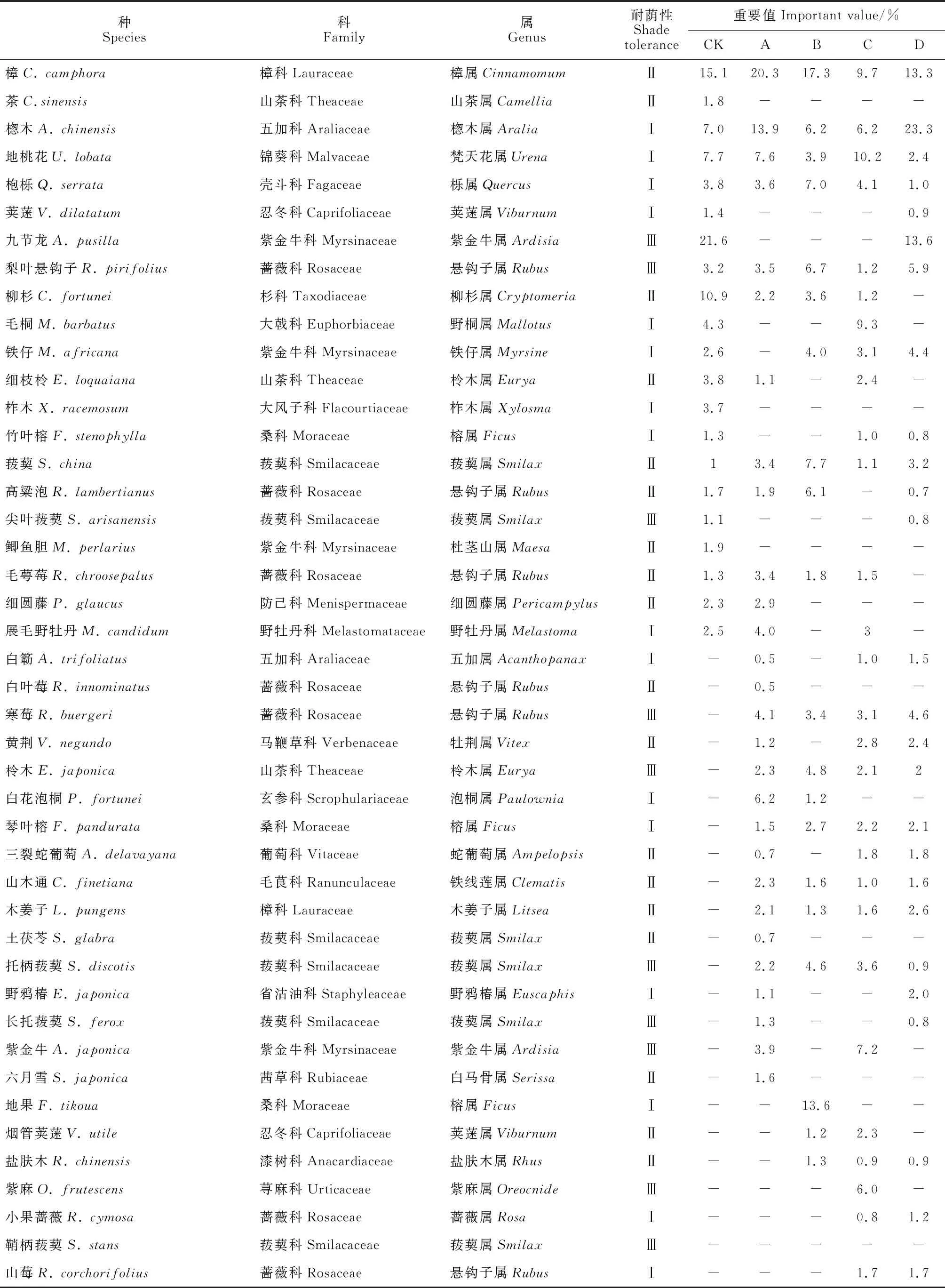
表2 马尾松人工林林窗下灌木层物种重要值及耐荫性
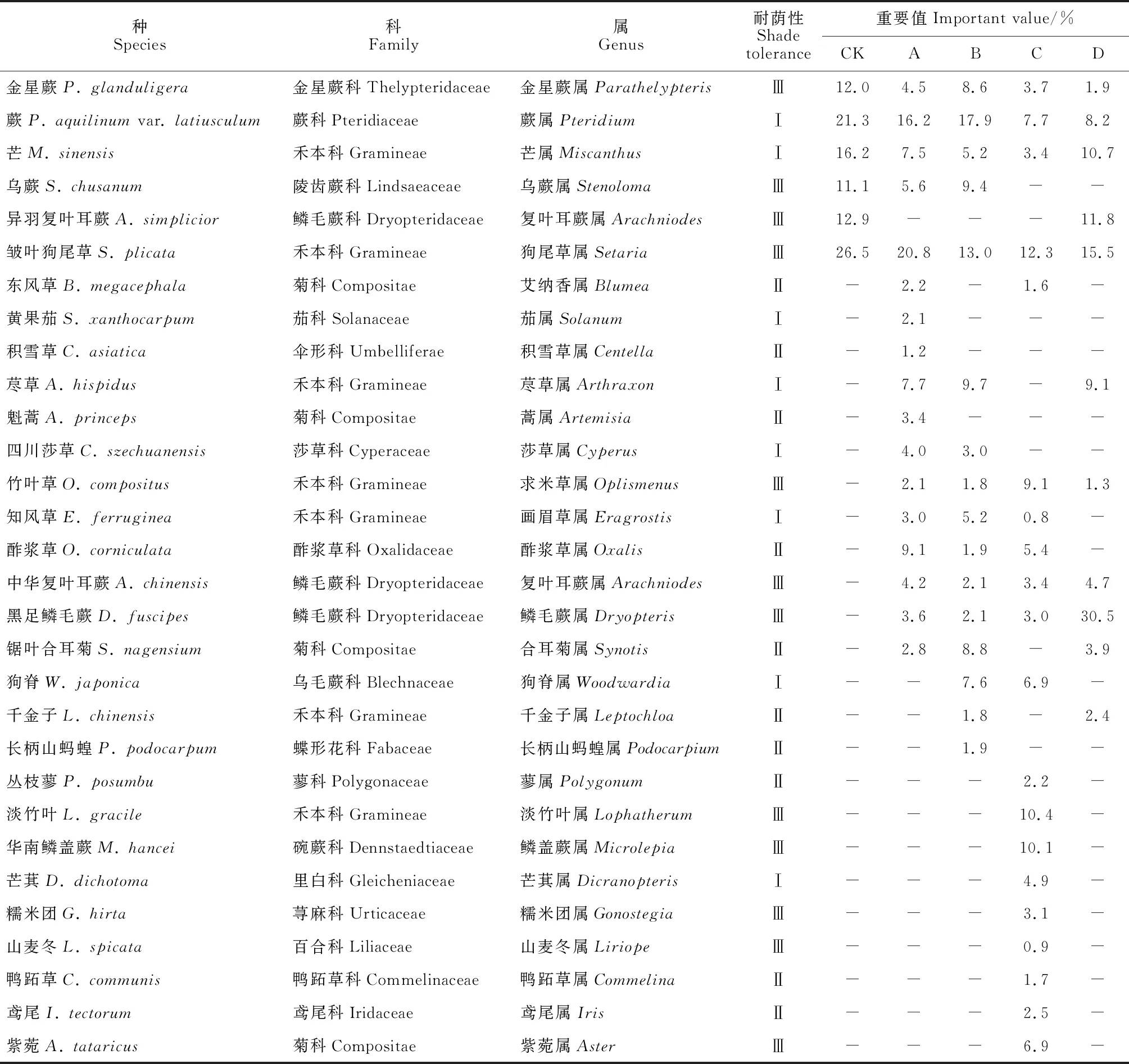
表3 马尾松人工林林窗下草本层物种重要值及耐荫性
续表2 Continued Table 2

注:Ⅰ.喜光;Ⅱ.中性;Ⅲ.耐阴; -.未发现。下同
2.4 林窗尺度对马尾松人工林灌草层优势种生态位宽度的影响
由表4可见,CK林下地桃花(0.343)、芒(0.368),A林窗地桃花(0.342)、荩草(0.359),B林窗枹栎(0.368)、荩草(0.368)和狗脊(0.357),C林窗地桃花(0.365)、狗脊(0.364),D林窗楤木(0.363)、荩草(0.367)和芒(0.346)生态位宽度均较大,且皆为喜光植物,说明开窗后林下光照增强导致喜光植物在灌草层中始终占据着优势地位,而适宜耐荫植物生长的荫蔽环境减少,耐荫植物竞争力不敌喜光植物。林窗下灌木层优势种生态位宽度平均值部分高于对照,部分低于对照,随林窗尺度增大,灌木层优势种生态位宽度平均值先减小后增大,且C林窗最小;各林窗下草本层优势种生态位宽度平均值均小于对照,其变化规律和灌木层一致。表明随林窗尺度的增大,灌草层优势种对资源的利用程度先降低而后利用更充分,C林窗各优势种对资源利用程度最低。各林窗下,灌木层优势种喜光植物地桃花和枹栎,草本层优势种喜光植物荩草和狗脊生态位宽度均较大。B林窗优势种地果、C林窗优势种紫麻、淡竹叶和华南鳞盖蕨的生态位宽度均为0,表明这4个种只存在于某个单一的资源位中。
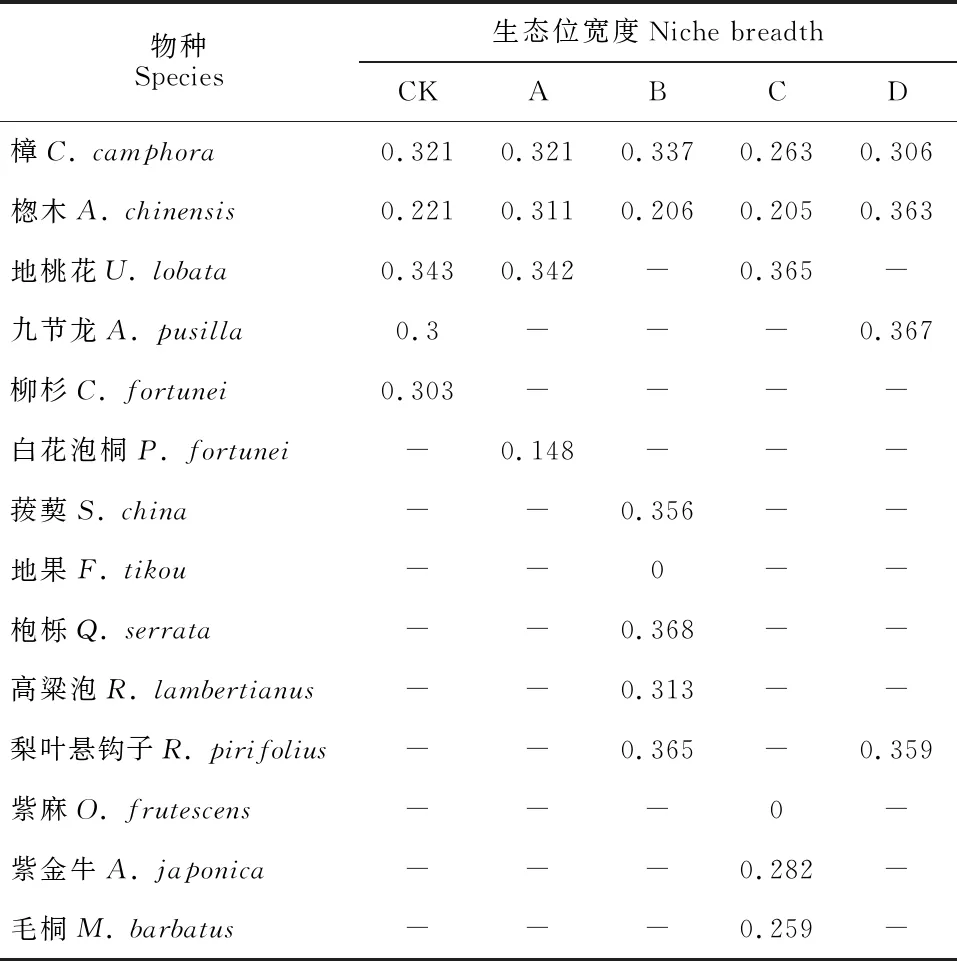
表4 马尾松人工林林窗下灌木层优势种生态位宽度
由表2~5可知,重要值与生态位宽度并不完全呈正相关关系。各林窗下灌木层优势种的重要值与生态位宽度在所有种中的排序大致相同,如CK灌木层的地桃花和樟的重要值分别为7.7%和15.1%,生态位宽度分别为0.343和0.321,而草本层却存在重要值与生态位宽度排序不同的情况,如CK草本层的芒重要值较小为16.2%,而其生态位宽度在优势种中最大为0.368。对各处理下灌草层优势种的重要值和生态位宽度进行Pearson相关性分析,结果表明(表6)两者无显著相关性(P>0.05)。
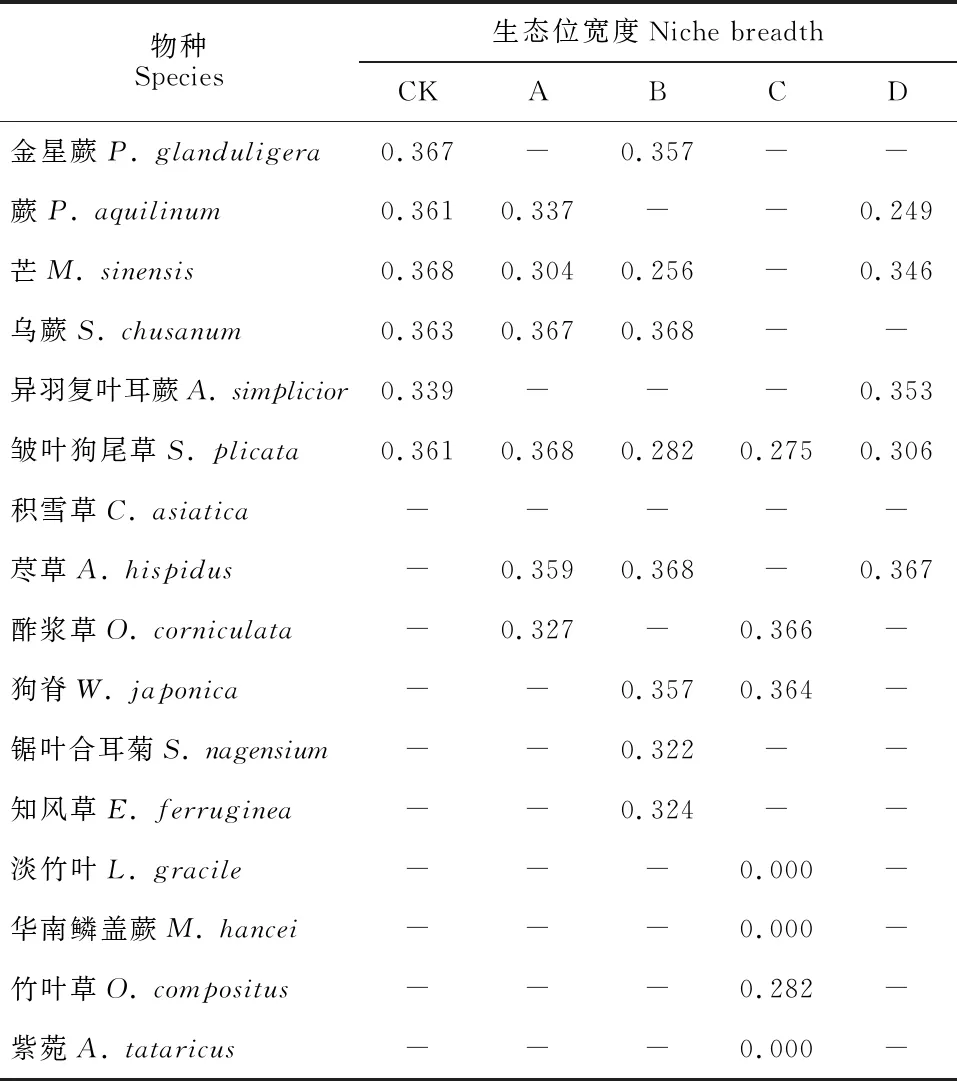
表5 马尾松人工林林窗下草本层优势种生态位宽度
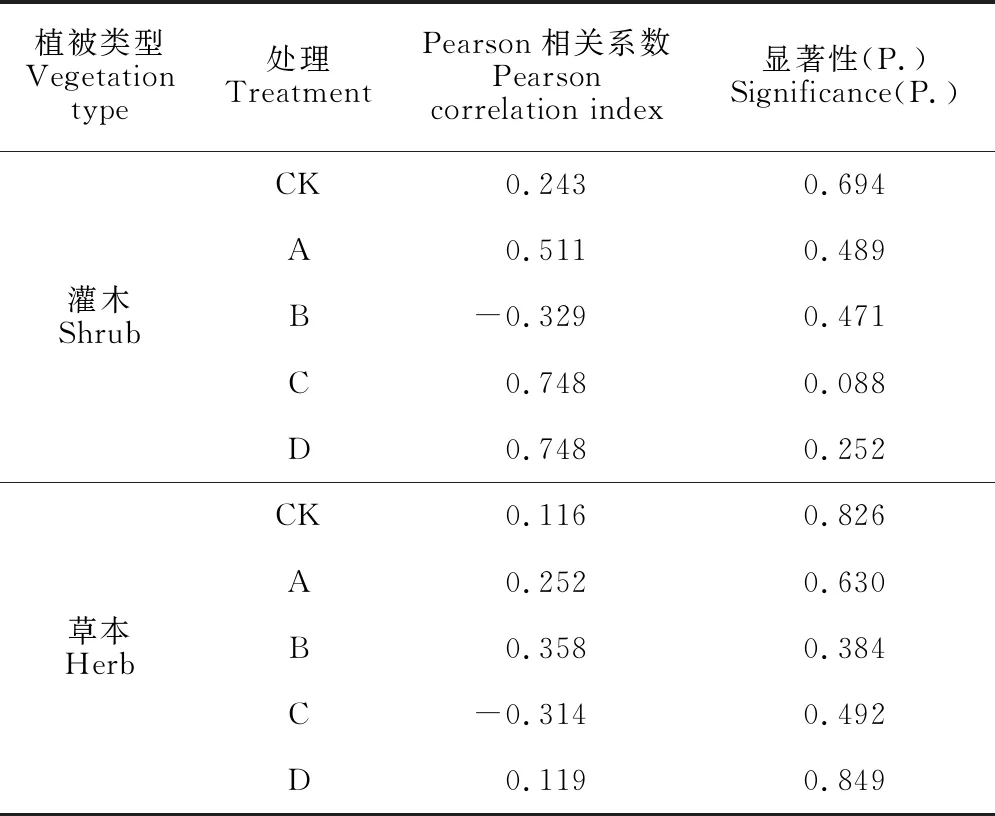
表6 马尾松人工林林下灌草层优势种重要值与生态位宽度的相关性系数
2.5 林窗尺度对马尾松人工林灌草层优势种间生态位重叠的影响
由表7和8可知,各处理林下灌草层优势种种间生态位重叠度值均在0.200以下,表明各处理下灌草层优势种间并未形成剧烈竞争,对生存空间的争夺现象不明显。其中,CK、A、B、C和D林窗灌木层优势种间生态位重叠度平均值分别为0.034、0.032、0.044、0.042和0.029,各草本层优势种间生态位重叠度平均值分别为0.054、0.035、0.042、0.048和0.024。由此可见,林窗下灌木层优势种间生态位重叠度平均值部分高于对照,部分低于对照,无明显规律,而草本层优势种生态位重叠度平均值均低于对照,表明开窗后林下草本层优势种间竞争减弱。各处理下,D林窗灌草层优势种间生态位重叠度平均值最小。CK灌木层优势种间生态位重叠度值高于0.080的种对数占总数的10.0%,其中最大为柳杉-九节龙(0.113);A林窗灌木层优势种间生态位重叠度值高于0.080的种对数占总数的8.3%,最大为樟-白花泡桐(0.080);B林窗灌木层优势种间生态位重叠度值高于0.080的种对数占总数的14.3%,最大为高粱泡-地果(0.184);C林窗灌木层优势种间生态位重叠度值高于0.080的种对数占总数的20.0%,最大为紫金牛-紫麻(0.182);D林窗灌木层优势种间生态位重叠度值均低于0.080,其中最大为九节龙-楤木(0.044)。CK草本层优势种间生态位重叠度值高于0.080的种对数占总数的3.3%,最大为乌蕨(Odontosoriachusana)-异羽复叶耳蕨(Arachniodessimplicior)(0.081);A林窗草本层优势种间生态位重叠度值均低于0.080,其中最大为皱叶狗尾草(Setariaplicata)-酢浆草(0.073);B林窗草本层优势种间生态位重叠度值高于0.080的种对数占总数的10.7%,最大为知风草(Eragrostisferruginea)-锯叶合耳菊(Synotisnagensium)(0.103);C林窗草本层优势种间生态位重叠度值高于0.080的种对数占总数的23.8%,最大为竹叶草(Oplismenuscompositus)-淡竹叶(0.182)、竹叶草-华南鳞盖蕨(0.182)和竹叶草-紫菀(Astertataricus)(0.182);D林窗草本层优势种间生态位重叠度值均低于0.080,最大为荩草-异羽复叶耳蕨(0.060)。在所有处理中,部分物种因较高的平均生态位重叠度而尤为突出,分别是A林窗白花泡桐、B林窗地果、C林窗紫麻、狗脊、竹叶草和酢浆草,表明它们占据着较多的资源,与其他物种竞争激烈。值得一提的是,B林窗地果、C林窗紫麻、C林窗淡竹叶、华南鳞盖蕨和紫菀与其他优势种间生态位重叠度值均为0,表明这5种植物与其他优势种在资源或空间利用上完全不存在重合部分。

表7 马尾松人工林林窗下灌木层优势种生态位重叠度
3 讨 论
3.1 林窗尺度对马尾松人工林林下灌草层物种丰富度的影响
丰富度指数表示局部地区均质生境中物种数量,许多研究都用其来指示物种多样性的变化[36]。本研究发现,开窗后马尾松人工林灌草层物种丰富度指数均显著提高,其随林窗尺度的增大呈现出先增大而后减小的变化规律,这与刘兵兵等[37]在杨桦次生林林窗下物种多样性研究中的发现类似。在四川南部开展的马尾松不同林窗尺度下植物多样性研究中,物种丰富度指数随林窗尺度的变化趋势和本研究大体一致[38]。可能的原因是林下植物生长主要与光照和水分等微环境有关,适当的林窗面积能保持林隙微环境及森林小气候,能提高生物多样性,而较大的林窗又接近无林地的微环境,不利于促进多种植物种类生长[37]。
3.2 林窗尺度对马尾松人工林林下灌草层生态位宽度的影响
生态位宽度表征物种对资源空间的利用程度[39]。本研究发现,各林窗下灌草层生态位宽度平均值随林窗尺度增大,呈现出先增大后减小的变化规律,且均在C林窗下降到最低值,可能的原因是C林窗极丰富的灌草层植物同时争夺空间和资源,导致各物种占据的平均资源和空间量最少。虽各林窗下灌草层中大多喜光植物生态位宽度较大,但部分林窗下耐荫植物也占据着优势地位,如A林窗皱叶狗尾草、B林窗梨叶悬钩子、C林窗紫金牛、D林窗九节龙和梨叶悬钩子的生态位宽度均较大,可能是因为占据优势地位的喜光灌木涨势迅猛为草本植物的生长创造了阴暗潮湿的环境。各林窗下喜光植物枹栎和荩草生态位宽度均较大,这与前人在华蓥市马尾松人工林开展的植物多样性研究结果类似[27],可能是本研究区各生态因子较适宜其生长。
重要值与生态位宽度是表征物种在群落中的地位的重要指标,但两者又有所不同,重要值代表物种在群落中的优势程度,而生态位宽度表示植物的生态位特征[40]。有研究表明,同一物种的重要值与生态位宽度在群落中有近乎相同的排序[41]。而本研究结果表明重要值与生态位宽度不存在明显相关性,这与陈霞等[42]研究结果相似,可能的原因是生态位宽度同时受重要值、种群个体分布和均匀度等多个因素的影响。
3.3 林窗尺度对马尾松人工林林下灌草层优势种间生态位重叠的影响
生态位重叠表征物种间利用相同资源和空间的程度[43]。生态位重叠较大的物种间可能因为争夺共同需要的资源而存在较大的竞争,也可能是因两者所需资源重叠过多占据了相近的生态空间而产生激烈竞争[44]。本研究结果表明各处理灌草层优势种间竞争较小,可能是因为本研究开展时间距开窗时间较近,林窗下灌草层植物群落未达到该条件下的稳定状态,此时资源和空间完全能够满足林下灌草层植物正常生长需要。各林窗草本层种间生态位重叠度较CK小,其中D林窗最小,这与已有的林窗对柏木人工林林下植被生态位影响的研究结果相似[45],可能是因为随林窗尺度进一步增大,丰富的林下资源能充分满足植物正常生长所需养分和光照等条件,此时竞争减弱。各处理中,C林窗灌草层种间高生态位重叠度所占比例最大,种间竞争最强,这可能与该林窗下植物种类最丰富有关。林窗尺度的增大使林下灌草层占据较多资源,竞争较强的物种由A林窗喜光植物白花泡桐、B林窗喜光植物地果变为C林窗耐荫植物紫麻和竹叶草,可能的原因是喜光植物的生长提供了适宜耐荫植物生长的阴暗潮湿环境。B林窗地果、C林窗紫麻、淡竹叶、华南鳞盖蕨和紫菀与其他物种间均不存在生态位重叠,可能是因为各物种的生物学习性和生态特征存在的差异使这5个种对资源和环境的利用与其他物种不存在相似性[46]。
4 结 论
开窗增加了林内光照,改变了林内微环境,促进了马尾松人工林林下灌草层植物发育,喜光植物较未开窗林分明显增多,林下灌草层优势种对资源和空间的利用程度先随林窗尺度增大而降低,后随林窗尺度进一步增大而逐渐变得充分,各优势种对资源和空间的竞争先增强而后逐渐减弱,表明开窗提高了马尾松人工林林下植物多样性,有利于其林下植物群落结构的改善,进而促进马尾松人工林更好地发挥生态效益。在各林窗中,200 m2林窗下植物物种最丰富,灌草层优势种中喜光植物均较多,此时植物对资源利用程度最低,优势种间对资源和空间的竞争最激烈,表明林下植物群落未达到稳定状态,可能与本研究调查时间距开窗仅1 a有关。综上,确定200 m2为提高马尾松人工林林下植物多样性、改善林下群落结构的最优林窗尺度。本研究开展野外调查时间距开窗时间较近,还处于试验早期阶段,植物群落未达到稳定状态,为探究开窗后马尾松人工林达到稳定状态时林下植被发育情况,后续可在该地区继续开展林窗下植物多样性及生态位相关研究。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
乡村科技(2021年1期)2021-12-03
农村实用技术(2021年6期)2021-12-01
新农业(2021年12期)2021-11-29
南方农业·中旬(2021年5期)2021-10-25
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
农民致富之友(2020年4期)2020-04-03
绿色科技(2019年5期)2019-11-29
农业与技术(2017年9期)2017-05-28
