
多巴胺受体激动剂、拮抗剂在药物成瘾中的应用基础研究进展*
2022-04-01 03:39:36李兵杜艳尉志文马栋贠克明苏红亮
中国药物滥用防治杂志 2022年2期
李兵,杜艳,尉志文,马栋,贠克明**,苏红亮**
(1.山西医科大学法医学院,山西 太原 030001;
2.司法鉴定科学研究院,上海市法医学重点实验室,司法部司法鉴定重点实验室,上海市司法鉴定专业技术服务平台,上海 200063)
药物成瘾(drug addiction)是由成瘾特性的药物引发的慢性、复发性脑疾病,其以强迫性的觅药和用药行为、越来越重的药物耐受现象,以及戒断后不可避免的负性情绪状态(如兴趣缺失、焦虑及易激惹)等为特点[1]。多巴胺系统在药物成瘾中发挥着重要作用,其中多巴胺受体(dopamine receptor)一直是研究者关注的焦点。多巴胺受体是与G 蛋白偶联的7 次跨膜受体,随着基因克隆技术的发展和应用,研究者先后克隆出了5 种可以被多巴胺激活的受体亚型。虽然多巴胺受体各亚型已被广大研究者所认知,但尚没有统一的书写格式,目前主要的中文书写格式有4 种(多巴胺DX 受体、DX 受体、多巴胺受体DX、DX 多巴胺受体),英文书写格式有8 种(dopamine DX receptor、DX receptor、dopamine receptor DX、DX dopamine receptor、DRDX、DXDR、DXR、DRX),本文统一采用“DX 受体”(X=1~5)的书写格式。
根据结构和功能上的不同,可以将多巴胺受体分为D1 样受体和D2 样受体两大家族,其中D1 样受体包括D1 受体和D5 受体,D2 样受体包括D2 受体、D3 受体和D4 受体。研究认为[2],D1 样受体主要激活突触后膜兴奋型G 蛋白偶联受体,从而引起细胞内环磷酸腺苷(cyclic adenosine monophosphate,cAMP)浓度升高;D2 样受体激活后主要作用于抑制型G 蛋白偶联受体,引起细胞内cAMP 浓度降低。在脑内,多巴胺受体各亚型的表达和分布存在明显差异,见表1。
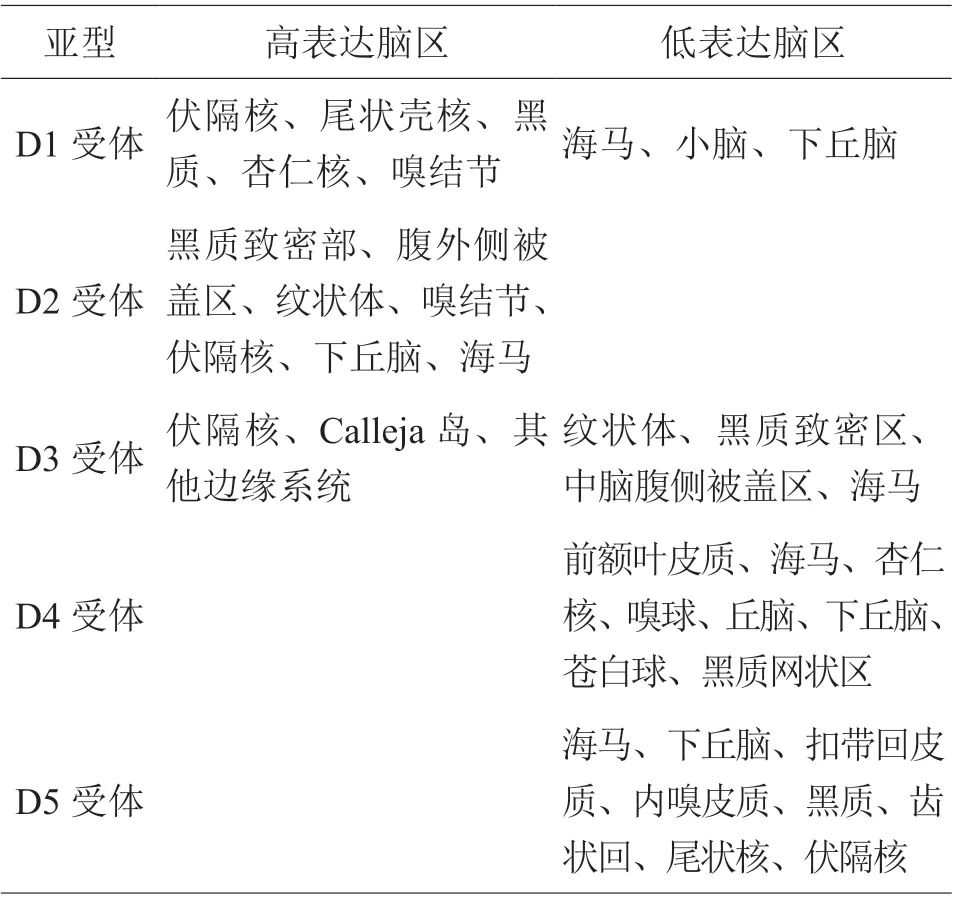
表1 多巴胺受体各亚型在脑内的表达和分布[3-6]
为了研究多巴胺受体各亚型在药物成瘾中分别发挥的具体作用,广大研究者选择应用众多针对多巴胺受体各亚型的激动剂、拮抗剂,高特异性的激动剂、拮抗剂逐渐在药物成瘾以及其他神经精神疾病的研究中显现出独特优势。据统计,目前常用的多巴胺受体各亚型的激动剂、拮抗剂见表2。
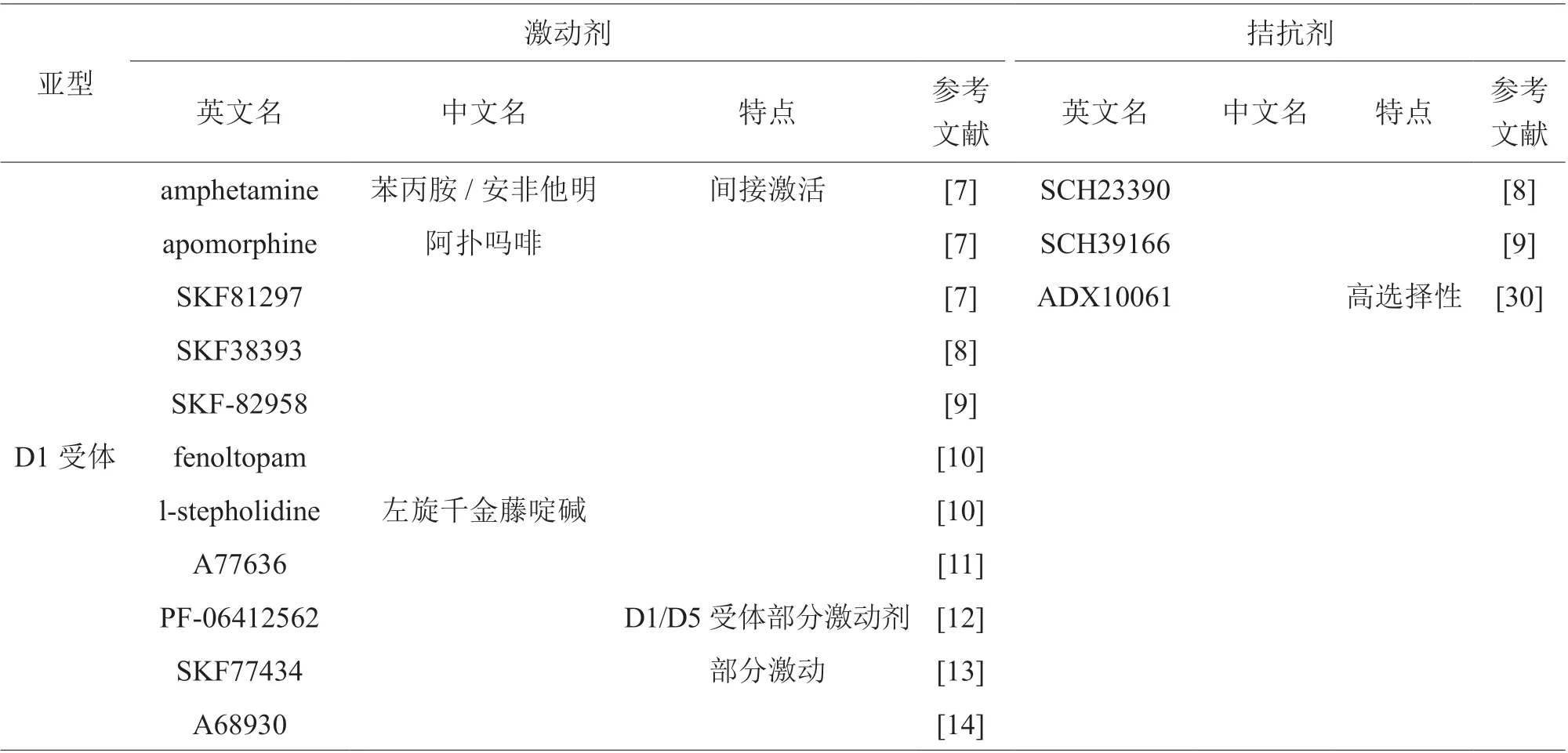
表2 常用的多巴胺受体各亚型的激动剂、拮抗剂
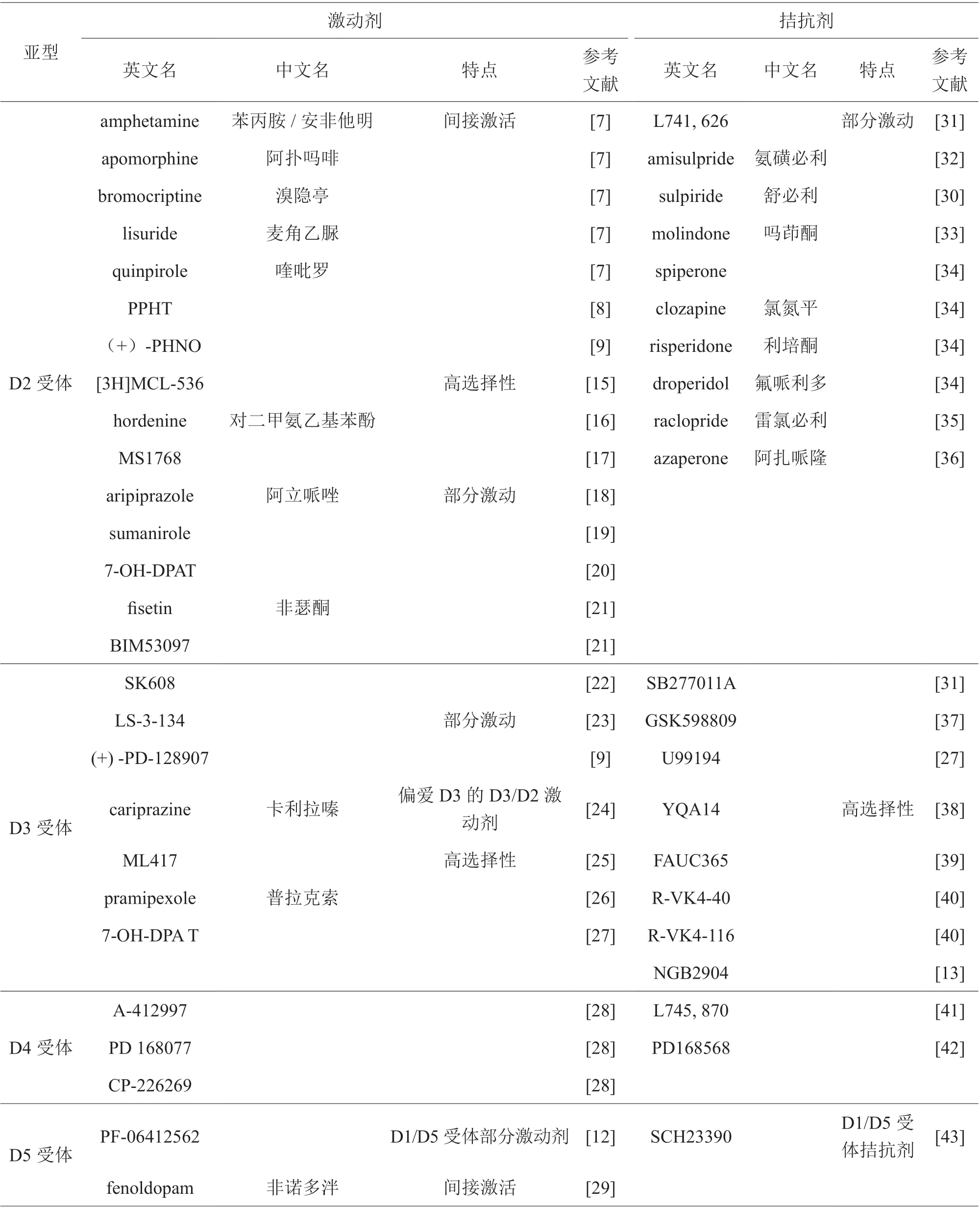
续表
1 多巴胺受体激动剂、拮抗剂在药物成瘾中的应用
药物成瘾的形成、巩固以及复发与脑内多巴胺受体有着密不可分的联系,多巴胺受体各亚型在这些过程中发挥着不同的作用。D1 受体、D2受体都参与了药物成瘾的奖赏、强化、增强运动、调控动机等,但它们的侧重点不同。D1 受体可能更多地参与动机调控,而D2 受体则更多地参与强化和依赖的形成[44]。D3 受体参与运动控制、认知、学习、奖励、情绪调节和社会行为等功能[45]。D4 受体在大脑中的分布有限,在成瘾药物复吸过程中具有独特的作用[46]。D5 受体可能更多地参与成瘾药物导致的运动活性增强[47]。选择性激活、阻断脑内多巴胺受体各亚型的信号传导,可能会揭示药物成瘾的潜在机制,开发出针对药物成瘾以及各种因素引起的复吸治疗方法,从而对药物成瘾的预防和治疗起到重要作用。
1.1 D1 受体激动剂、拮抗剂的应用
在不同的行为学模型中,D1 受体影响着成瘾的不同方面。D1 受体激动剂SKF81297 缩短了吗啡成瘾大鼠到达水迷宫平台的潜伏期,增加了大鼠跨越水迷宫平台的次数,同时增加了大鼠中脑导水管周围灰质(periaqueductal grey,PAG)中谷 氨 酸 脱 羧 酶67(glutamate decarboxylase 67,GAD67)的表达[8],以上研究结果表明D1 受体激活后可以通过增加GAD67 的表达水平来提高吗啡成瘾过程中的学习记忆能力。
GU SM 等[48]研究成瘾药物诱导的条件性位置偏爱(conditioned place preference,CPP)与多巴胺受体敏感性的实验中发现,D1 受体拮抗剂SCH23390 可减少甲基苯丙胺和可卡因诱导的CPP的表达,而且SCH23390 预处理可抑制多巴胺受体激动剂apomorphine 引起的运动活动增加,表明SCH23390 可通过抑制D1 受体减弱药物成瘾的发生,是治疗药物成瘾的潜在药物。肖琳等[49]研究也得到了一致的结果,其发现SCH23390 可以显著降低吗啡作用后小鸡CPP 的表达,从而证明D1受体参与了药物成瘾的形成。
1.2 D2 受体激动剂、拮抗剂的应用
与D1 受体激活不同的是,D2 受体部分激动剂Terguride 可剂量依赖性地减少海洛因固定比例和递增比例自主给药的最大反应次数,这种功能特异性的作用表明Terguride 能够阻止海洛因的强化,可能是治疗阿片成瘾的一种新的潜在药物[50]。然而,De Vries TJ 等[51]通过动物模型观察到,在静脉注射可卡因和海洛因的早期戒断中,D2 受体激动剂quinpirole 会导致可卡因和海洛因成瘾的快速恢复;对戒断早期运动活动的平行分析显示,quinpirole 的精神运动兴奋作用得到了强烈增强,并且用可卡因训练的动物停药三周后仍能观察到quinpirole 引起的觅药行为和行为敏化,这可能是由于quinpirole 的高反应性与动物觅药行为之间存在较强的时间相关性。
Milivojevic N 等[52]在研究具有D2 受体拮抗剂和D1 受体激动剂双重特性的LEK-8829 对大鼠可卡因自主给药影响的实验中发现,LEK-8829 减少了可卡因自主给药的摄入量,并减弱了可卡因的复吸,LEK-8829 对可卡因自主给药及复吸的影响与其具有较低的滥用潜力有关,表明LEK-8829可能是治疗可卡因滥用的候选药物。余化霖等[53]用D2 受体拮抗剂研究吗啡诱导的大鼠CPP 时却发现了相反的现象,在吗啡戒断后,大鼠眶额叶内微量注射D2 受体拮抗剂s(-)-Raclopride-(+)-tartrate salt,大鼠显示出比对照组更强的CPP 的表达,表明阻断眶额叶内的D2 受体会强化药物渴求的程度;通过与D2 受体激动剂组的数据相比,其认为这可能是由于眶额叶内的D2 受体对药物渴求起抑制作用,当D2 受体受到抑制时眶额叶对吗啡渴求行为的抑制被削弱,导致动物渴求行为明显增加。上述研究表明,不同脑区的多巴胺受体亚型在药物成瘾中发挥的具体作用可能不同。
1.3 D3 受体激动剂、拮抗剂的应用
D3 受体与多巴胺的结合能力是所有受体亚型中最高的,因此在研究药物成瘾中占有重要地位。Galaj E 等[13]在研究大鼠D3 受体和D1 受体在可卡因成瘾中的作用时发现,在觅药行为恢复或CPP 测试之前同时注射D3 受体拮抗剂NGB2904和D1 受体部分激动剂SKF77434 可显著减少大鼠杠杆按压的次数和在可卡因伴药侧花费的时间;在自主给药模型中,当给大鼠按递增比例给药时,单独注射NGB2904 无效,但却显著增强了SKF77434 的降低断点的效果,表明在可卡因成瘾中D3 受体和D1 受体之间存在着相互作用。Chen L 等[54]运用基因敲除小鼠的研究结果也证明了这一点,他们认为D3 受体和D1 受体通过调节(至少在一定程度上)大脑特定区域的细胞外信号 调 节 激 酶(extracellular signal-regulated kinase,ERK)的激活,在线索和可卡因奖赏属性之间的学习关联中发挥了作用。
Lv Y 等[38]在D3 受体拮抗剂YQA14 对吗啡成瘾影响的研究中发现,YQA14 不仅抑制吗啡诱导的行为敏化的获得,而且显著减弱吗啡诱导的行为敏化的表达,表明阻断D3 受体可抑制吗啡诱导的行为敏化,提示D3 受体在吗啡成瘾中起重要作用,可能成为临床治疗阿片成瘾的潜在靶点。Sun L 等[55]研究也表明,重复注射和单次注射YQA14(6.25 mg/kg、12.5 mg/kg 和25 mg/kg)均剂量依赖性地抑制了甲基苯丙胺诱导的行为敏化的获得和表达;然而,在CPP 试验之前单次注射YQA14 可剂量依赖性地减弱甲基苯丙胺诱导的CPP 的表达(重复注射不改变),虽然重复注射YQA14 对甲基苯丙胺诱导的CPP 的表达没有影响,却剂量依赖性地促进了甲基苯丙胺诱导的CPP 的消退,证明了D3 受体在甲基苯丙胺的奖赏作用中起着至关重要的作用。Hachimine P等[56]研究运用选择性的D3 受体拮抗剂SR 21502来研究可卡因诱导的大鼠CPP 表达能力,结果发现SR 21502 阻断了大鼠对可卡因伴药侧的偏好,减弱了可卡因线索诱导的奖赏效应,提示对D3 受体进行抑制可以减弱可卡因的成瘾作用,表明该化合物也可能是一种治疗可卡因成瘾的有效药物。
1.4 D4 受体激动剂、拮抗剂的应用
D4 受体在脑内的分布量与D3 受体相似,在药物成瘾的形成中起着重要的作用。Rivera A 等[57]研究发现,D4 受体激动剂PD168,077 可防止吗啡诱导的奖赏、多动和身体依赖的形成,这可能与D4 受体激活后和μ 阿片受体相互作用,减弱吗啡引起的黑质纹状体多巴胺通路的激活和黑质致密部多巴胺神经元的形态变化有关。Kim A 等[41]在研究D4 受体激动剂PD168,077 和选择性拮抗剂L745,870 对酒精自主给药和复吸的影响时发现,PD168,077 对应激诱导的复吸和线索诱导的复吸都没有影响,而L745,870 在高剂量下减少了酒精的摄入量,并且减少了应激诱导的复吸(对线索诱导的复吸没有影响),这些结果表明激动D4 受体对复吸没有作用,而阻断D4 受体可以减少应激诱导的复吸。同样,Yan Y 等[58]在研究L745,870 对尼古丁复吸行为影响的实验中也发现,L745,870 显著减弱相关线索或预先注射尼古丁诱导的复吸,表明D4 受体拮抗剂可能是一种治疗吸烟复吸的潜在药物。Mamiya T 等[59]研究发现,L745,870 可以在吗啡成瘾小鼠中通过抑制cAMP 系统的上调,降低吗啡成瘾的表达,表明D4 受体拮抗剂也可以作为一种治疗药物来减弱吗啡成瘾的发生。
关于D5 受体激动剂、拮抗剂在药物成瘾中的应用研究较少,有待进一步探索。
2 多巴胺受体激动剂、拮抗剂在药物成瘾中应用的影响因素
2.1 激动剂、拮抗剂对受体的选择性
由于多巴胺受体在药物成瘾中的作用复杂,大多数激动剂、拮抗剂不能严格区分相同家族的受体亚型,并且激动剂、拮抗剂均具有自己独特的药效特点,从而产生不同程度的激动、拮抗效应。以D3 受体的拮抗剂为例,每一种拮抗剂具有不同的抑制常数(inhibition constant,Ki,如KiBAK4-54=0.12、KiBP897=0.92、KiGSK598809=1.25、KiSR21502=4.2、KiR-VK4-116=7.4、KiSB-277011A=10.7),这些抑制常数指检测到50%抑制效果时拮抗剂的浓度,即反映拮抗剂对靶标的抑制强度,这个值越小说明抑制能力越强。在开展研究时,应尽可能地选择Ki 值较低的拮抗剂,这样才能保证更好的抑制效果,实验结果才更有说服力,以证明被抑制的受体在成瘾过程中的作用。
由于5 种多巴胺受体亚型识别相同的内源性配体——多巴胺,进而激活受体,但各亚型具有很高的序列同源性,以往大多数化合物不能严格区分各亚型,由此形成的配体对多巴胺受体的选择性激活机制还有待研究。特别值得关注的是,在2021 年初,我国两个研究团队采用单颗粒冷冻电镜技术,针对多巴胺受体亚型的药物开发取得重要进展,从原子分辨率层面解析了多巴胺受体亚型之间对配体(激动剂或拮抗剂)选择性识别、激活机理,以及与效应G 蛋白信号转导复合物选择性偶联的分子机制[60-61],为开发以多巴胺受体为靶标的高效选择性配体药物提供了重要的结构模式和理论依据,也为药物成瘾以及其他与多巴胺有关的神经精神疾病(如帕金森病、阿尔茨海默病、精神分裂症等)的治疗带来新的研究思想。
2.2 多巴胺受体基因敲除动物的应用
由于多巴胺受体各亚型结构上的相似性,某受体亚型的激动剂、拮抗剂或多或少会对其他多巴胺受体亚型(特别是同一家族)产生作用,会干扰对特定多巴胺受体亚型作用的研究。因此,研究者为了更好地开展实验,除了使用更具选择性的拮抗剂外,还倾向于使用多巴胺受体基因敲除小鼠作为实验动物来达到更直观的实验效果。多巴胺受体基因定向敲除后仅有对应的多巴胺受体的表达缺失,其他功能不受影响,其研究具有针对性强、特异性高的优势,可以真实地反映多巴胺受体在药物成瘾中的调控作用,对于揭示药物成瘾的神经生物学机制具有重要意义。因此,多巴胺受体基因敲除小鼠在研究多巴胺受体功能时的使用得到了广泛认可。
2.3 各种动物行为学模型的应用
在应用激动剂、拮抗剂的同时,动物行为学模型也是决定多巴胺受体各亚型在药物成瘾中发挥作用的关键因素,研究者往往根据需要探索的激动剂、拮抗剂对成瘾药物导致的特定行为特征的影响,从而选择适合的行为学模型。
目前,常用的动物行为学模型主要包括:条 件 性 位 置 偏 爱(conditioned place preference,CPP)、行 为 敏 化(behavioral sensitization,BS)和自主给药(self-adiminstration,SA),这些模型均是基于各种行为学理论以及成瘾机制,从各个方面模拟药物成瘾者的成瘾行为。以上三种模型各有其应用的优点和缺点:①CPP 是一种经典的反映成瘾性药物精神依赖性的反应性奖赏强化模型,实验动物不因受试药物是否妨碍运动机能(自发活动专门用于评价药物对运动机能的影响)而影响实验结果,其流程简单、方便快捷(不需要给动物做手术,只需要几个训练循环即可),已被证明对评估奖赏效应非常有效[62]。然而,CPP模型也有一定的缺点,如实验动物的给药过程是被动的,难以反映实验动物自身的需求;实验测试当天,实验动物可能会受到新环境的干扰而无法得到剂量依赖曲线。②BS 被认为是动机敏化,是从偶然性用药到强迫性用药的转变,具有良好的结构效度,易于操作,实验周期短,是研究药物成瘾机制、药物成瘾潜能和抗成瘾药效评价的良好模型[63-64],但该模型也无法排除由于连续性用药导致的与行为敏化无关的神经适应性改变,而这种改变可能对成瘾的形成产生促进作用,从而干扰该模型本身的假设[63]。③SA 模型是目前被广泛认可的模拟成瘾过程的最佳模型,在这个模型中动物可以通过不同的给药方式完成各种成瘾药物的自我给药行为。由于这个模型有非常好的结构有效性,常被用于研究成瘾行为的整个过程,如药物初始的奖赏作用,戒断(消退)和复吸阶段等[63,65]。但SA 模型在操作过程中需要麻醉实验动物,通过手术给脑内留置导管,这会对实验动物造成痛苦与伤害,干扰实验结果。
2.4 不同的给药方式
动物行为学模型在决定多巴胺受体各亚型的激动剂、拮抗剂发挥作用的过程中,所研究的成瘾药物和多巴胺受体各亚型的激动剂、拮抗剂的给药方式也是决定研究成功与否的重要方面。总体而言,以往关于成瘾药物和激动剂、拮抗剂的给药方式大都采用整体给药方式(如口服给药、皮下注射、腹腔注射、静脉注射),仅少数研究采用局部给药方式(如脑立体定位颅内注射),特别是对于激动剂和拮抗剂。
在啮齿类动物中,口服给药最常见于研究酒精的成瘾特性,在尼古丁、可卡因、甲基苯丙胺、吗啡等研究中少见,这是因为即使此种给药方式不会对实验动物造成伤害,啮齿类动物很少通过其他给药途径饮酒,并且尼古丁、可卡因、甲基苯丙胺、吗啡等成瘾药物通过口服给药很难发挥药效[66]。皮下注射、腹腔注射、静脉注射在CPP和BS 模型中常用,完成注射简单、省时、便利,药物快速产生效应,但被动的给药操作很难真实体现实验动物对药物的需求,容易对实验结果的真实性产生一定的干扰。
此外,鉴于不同脑区各多巴胺受体亚型在药物成瘾中发挥的作用不同,通过使用脑立体定位核团微量注射各受体亚型的激动剂、拮抗剂,可以精准地研究在所关注的脑区核团该受体亚型在药物成瘾中发挥的具体作用,为研究成瘾药物的神经药理学、神经解剖学基础以及在整个成瘾过程中的作用提供了良好的手段[65]。
3 多巴胺受体激动剂、拮抗剂在其他神经精神疾病中的应用
选择性激活、阻断脑内多巴胺受体各亚型的信号传导,不仅对药物成瘾起作用,在其他神经精神疾病的预防和治疗中也具有重要作用。例如,D1 受体激动剂PF-3628 可能成为治疗人类认知障碍的新药物[67],D1 受体部分激动剂PF-06669571可能会成为帕金森病的新疗法[68];放射性标记的D2 受体激动剂[3H]MCL-536 可能成为诊断和治疗神经精神疾病的通用工具,可在帕金森病和精神分裂症患者体内定量D2 受体高亲和力位点[15];D2/D3 受体部分激动剂Cariprazine 可以显著减弱野生型小鼠的慢性不可预测的应激性快感缺乏样行为,表明Cariprazine 具有治疗抑郁症的作用[69];选择性D1/D5 受体部分激动剂PF-06412562辅助治疗精神分裂症具有安全、耐受性好的特点[12]。
4 总结
成瘾药物的作用环节大都集中于脑内多巴胺系统,应用多巴胺受体各亚型的激动剂、拮抗剂来研究多巴胺受体在药物成瘾中的作用效果显著,本文所综述的研究为寻找有效的药物成瘾新疗法提供了重要依据和借鉴意义。在这些研究的基础之上,今后还需要筛选更多种类的激动剂、拮抗剂,进一步探讨多巴胺受体及其信号传导通路在药物成瘾中的作用,揭示药物成瘾的神经生物学机制,从根本上防范日益加剧的成瘾药物滥用,减少成瘾药物滥用给公共安全带来的风险隐患,维护国家的长治久安。
猜你喜欢
中成药(2018年10期)2018-10-26 03:41:22
天然产物研究与开发(2018年6期)2018-07-09 06:01:46
海南医学(2016年8期)2016-06-08 05:43:00
中国社区医师(2015年14期)2015-12-24 00:37:31
医学研究杂志(2015年5期)2015-06-10 06:43:26
生殖医学杂志(2015年11期)2015-02-28 16:32:16
发明与创新(2015年29期)2015-02-27 10:39:38
中华皮肤科杂志(2014年4期)2014-12-19 12:55:43
中国神经精神疾病杂志(2013年1期)2013-03-11 20:23:36
祝您健康(1998年11期)1998-12-25 10:18:52
