
初生和成年黑藏羊肉品质与肌纤维特性差异分析
2022-03-30 02:29高占红侯生珍杨葆春王志有桂林生
畜牧兽医学报 2022年3期
周 力,高占红,侯生珍,杨葆春,王志有,桂林生
(青海大学农牧学院,西宁 810016)
黑藏羊俗称黑裘皮羊,以生产黑色二毛裘皮著称,是藏系绵羊中重要的经济类群[1]。当前黑藏羊产业由生产裘皮向肉用逐步转型,若能了解其肌纤维类型分布以及肉质等变化规律,对于保护地方品种资源,提高农牧民经济收入以及推动青海特色畜牧业发展均具有重要意义。长期以来,羊肉产品以高蛋白、低脂肪、低胆固醇、味道鲜美、富含维生素和矿物质特征而备受青睐。羊肉肉质主要受饲料种类、饲养方式以及遗传因素的影响[2],肉羊年龄也是一个重要方面[3]。研究发现,相对于50日龄羔羊,100日龄的羔羊肉中干物质、总蛋白质和粗蛋白质含量更高,而50日龄的羔羊肉则具有更强的持水力[4]。其他试验也表明,不同年龄组间牦牛背最长肌中过氧化氢酶、总超氧化物歧化酶、谷胱甘肽过氧化物酶和总抗氧化能力差异均不显著,但随着年龄的增长,丙二醛含量不断上升,4.5岁组比1.5岁组升高了53.80%[5]。由此可以看出,不同发育时期反刍动物肌肉中营养成分与抗氧化能力均存在一定差异性。
肌纤维类型是决定肌肉特性的重要因素,根据肌动蛋白重链上ATP酶对酸碱的稳定性可以将肌纤维分成3种类型,即Ⅰ型(慢速氧化型肌纤维)、ⅡA型(快速氧化型肌纤维)和ⅡB型(快速酵解型肌纤维)[6]。王丽莎等[7]指出,不同类型的肌纤维代表着不同的结构、功能及代谢特征。当肌肉中氧化型肌纤维所占比例较高时,肌肉细嫩多汁,系水力强,肉质也较好[8]。以土耳其地方绵羊品种为研究对象,发现肌纤维的形态特性,如肌纤维类型、数目和直径会对肉品质产生重要的影响[9]。日粮添加乳酸菌试验表明[10],可以通过提高氧化型肌纤维的比例改善肌肉色泽,提高肌肉嫩度,继而改善苏尼特羊肉品质。在肉羊上也进一步证实,肌球蛋白重链基因表达水平的变化能够反映肌纤维类型转化的程度[11]。由此可知,肉质特性受年龄的影响较大,且不同年龄的肌肉品质与其肌纤维类型以及MyHCs基因的mRNA表达量差异关系密切。
目前,现阶段有关黑藏羊的研究相对匮乏,仅有针对不同性别黑藏羊肉品质特性的差异分析[12]。基于此,本试验以自然放牧条件下的初生羔羊和成年黑藏羊为试验对象,通过对其肌肉常规营养成分、抗氧化能力、肌纤维组织学特性及其MyHCs基因的表达水平进行分析比较,进而为开展青海黑藏羊肉品质相关研究提供理论参考。
1 材料与方法
1.1 材料和试剂
试验选用青海省海北州贵南县自然放牧条件下发育正常、健康无病的初生黑藏羊公羔(平均体重(2.31±0.49)kg)和12月龄成年黑藏羊公羊(平均体重(36.58±1.26)kg)各5只。试验羊只屠宰后,采集其左侧胴体第12~13肋骨间背最长肌,液氮保存待用。
包埋剂(日本Sakura公司),二甲苯(广东光华科技股份有限公司),乙醇(天津富宇精细化工有限公司),总抗氧化能力、丙二醛含量,超氧化物歧化酶、谷胱甘肽过氧化物酶和过氧化氢酶活性试剂盒(南京建成生物工程研究所有限公司),RNA Prep Pure总RNA(动物组织)提取试剂盒D431,FastQuant cDNA第一链合成试剂盒(去基因组)KR106和SuperReal PreMix Color(SYBR Green)试剂盒等均购自北京天根生化科技有限公司。
1.2 仪器与设备
德国LEICA切片机(德国Leica公司);美国Bio-Rad伯乐凝胶成像分析系统(北京赛百奥科技有限公司);罗氏实时荧光定量PCR仪(北京众力挽生物科技有限公司);科华酶标仪(上海科华生物工程有限公司)。
1.3 方法
1.3.1 肌肉常规营养成分测定 水分(moisture)含量参照GB5009.3—2016进行测定;粗灰分(ash)含量参照GB5009.4—2016进行测定;粗蛋白(crude protein, CP)含量参照GB/T5009.5—2016进行测定;粗脂肪(ether extract, EE)含量参照GB 5009.6—2016进行测定。
1.3.2 肌肉抗氧化指标检测 抗氧化指标包括超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶活性,总抗氧化能力及丙二醛含量,具体操作步骤严格按照试剂盒说明书进行。
1.3.3 肌纤维组织形态学特性测定
1.3.3.1 样品采集和制片:将样品沿着肌纤维的横向方向切成0.5 cm×0.5 cm×1.0 cm大小的肉样,经过预冷的异戊烷干燥30 s后,将肉块放入冰冻切片机内进行切片(厚度约8 μm),再利用碱孵育ATPase对黑藏羊背最长肌进行染色。
1.3.3.2 肌纤维各项指标分析:利用显微镜彩图分析软件在20×10倍下观察并分析肌纤维组织学特性的相关指标。每个片子随机选取3~5个视野,使测定的肌纤维总数大于1 500根。1)肌纤维直径(muscle fiber diameter):测量每根肌纤维横截面最长两点距离,作为长轴,垂直于长轴的最短两点距离,作为短轴,多次测量求其平均值;2)肌纤维横切面积(muscle fiber cross-sectional area):圈出视野内各型肌纤维的横截面积,测多根取其平均值;3)肌纤维密度(muscle fiber density):计算每个视野的面积,并统计每个视野内肌纤维的根数,将其换算成每平方毫米的根数;4)肌纤维数量比例(number rate of muscle fiber):每种类型肌纤维数量除以肌纤维总数量即为肌纤维数量比例,公式如(1)所示:
(1)
5)肌纤维面积比例(area rate of muscle fiber):每种肌纤维总面积除以总肌纤维面积即为肌纤维面积比例,公式如(2)所示:
(2)
1.3.4MyHCs基因表达量测定 利用动物组织总RNA提取试剂盒提取黑藏羊肌肉中总RNA,利用1.5%琼脂糖和超微量核酸蛋白仪分别检测RNA完整性、浓度和纯度,将符合要求的RNA反转录成cDNA。按照SuperReal PreMix Color(SYBR Green)进行PCR扩增。qPCR反应体系为:1.6 μL cDNA模板,上、下游引物各0.8 μL,2×SuperReal Color PreMix 10 μL,RNase-free ddH2O 6.8 μL。反应条件为:95 ℃预变性 15 min,95 ℃变性 10 s,60 ℃退火/延伸 30 s,40个循环。引物由上海生工生物设计合成。内参基因(GAPDH)和目的基因(MyHCI、MyHCⅡA及MyHCⅡB)序列如表1所示。
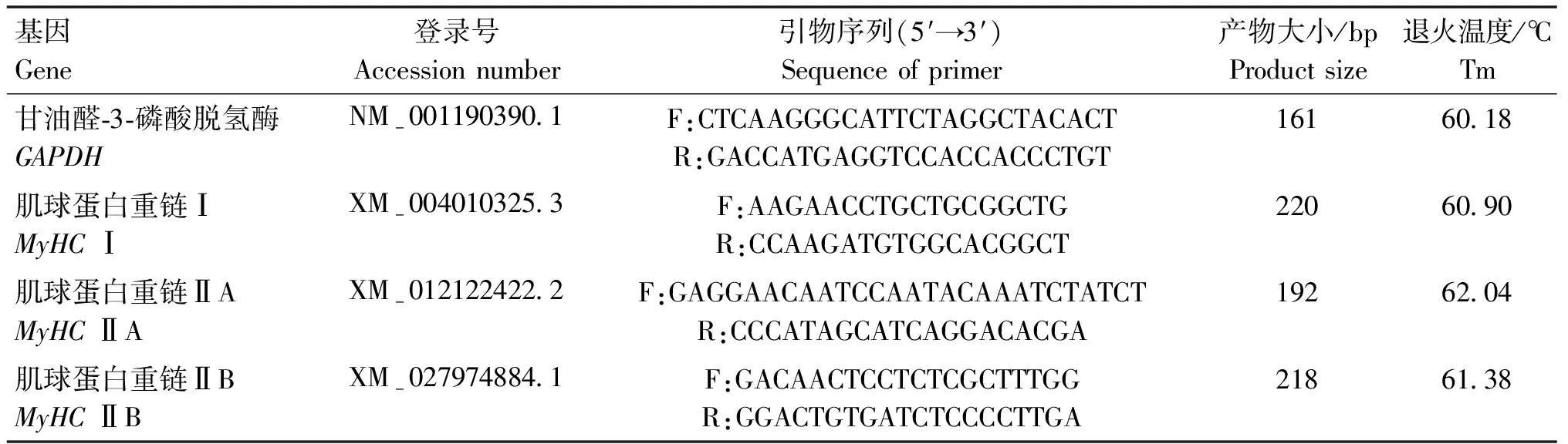
表1 引物序列
1.4 统计分析
本研究采用2-ΔΔCt法作为目的基因表达量的分析方法[13],试验数据经Excel(2020)初步计算处理,在初生羔羊和成年羊间的数据差异作独立样本t检验,试验结果均以“平均值±标准差(mean±SD)”表示,以P<0.05作为显著性判断的标准。
2 结 果
2.1 不同生长期黑藏羊背最长肌常规营养成分的比较
由表2可知,相较于初生羔羊,成年羊背最长肌中水分(moisture)、灰分(ash)、粗蛋白(CP)以及粗脂肪(EE)含量差异均不显著(P>0.05)。这说明黑藏羊肌肉中营养成分的含量受年龄影响较小。

表2 黑藏羊肌肉常规营养成分指标分析
2.2 不同生长期黑藏羊背最长肌抗氧化能力的比较
由表3可知,与初生羔羊相比,成年羊背最长肌中总抗氧化能力(T-AOC)、丙二醛(MDA)含量,超氧化物歧化酶(SOD)、过氧化氢酶(CAT)以及谷胱甘肽过氧化物酶(GSH-Px)活性差异均不显著(P>0.05)。这表明不同发育阶段对黑藏羊肌肉抗氧化能力影响不显著。

表3 黑藏羊肌肉抗氧化指标分析
2.3 不同生长期黑藏羊背最长肌组织形态学特性的比较
肌纤维直径分析结果如图1所示。初生羔羊背最长肌肌纤维直径Ⅰ型、ⅡA型和ⅡB型均显著小于成年羊(P<0.05)。由此可见,不同年龄阶段黑藏羊肌纤维直径Ⅰ型、ⅡA型和ⅡB型差异较大。
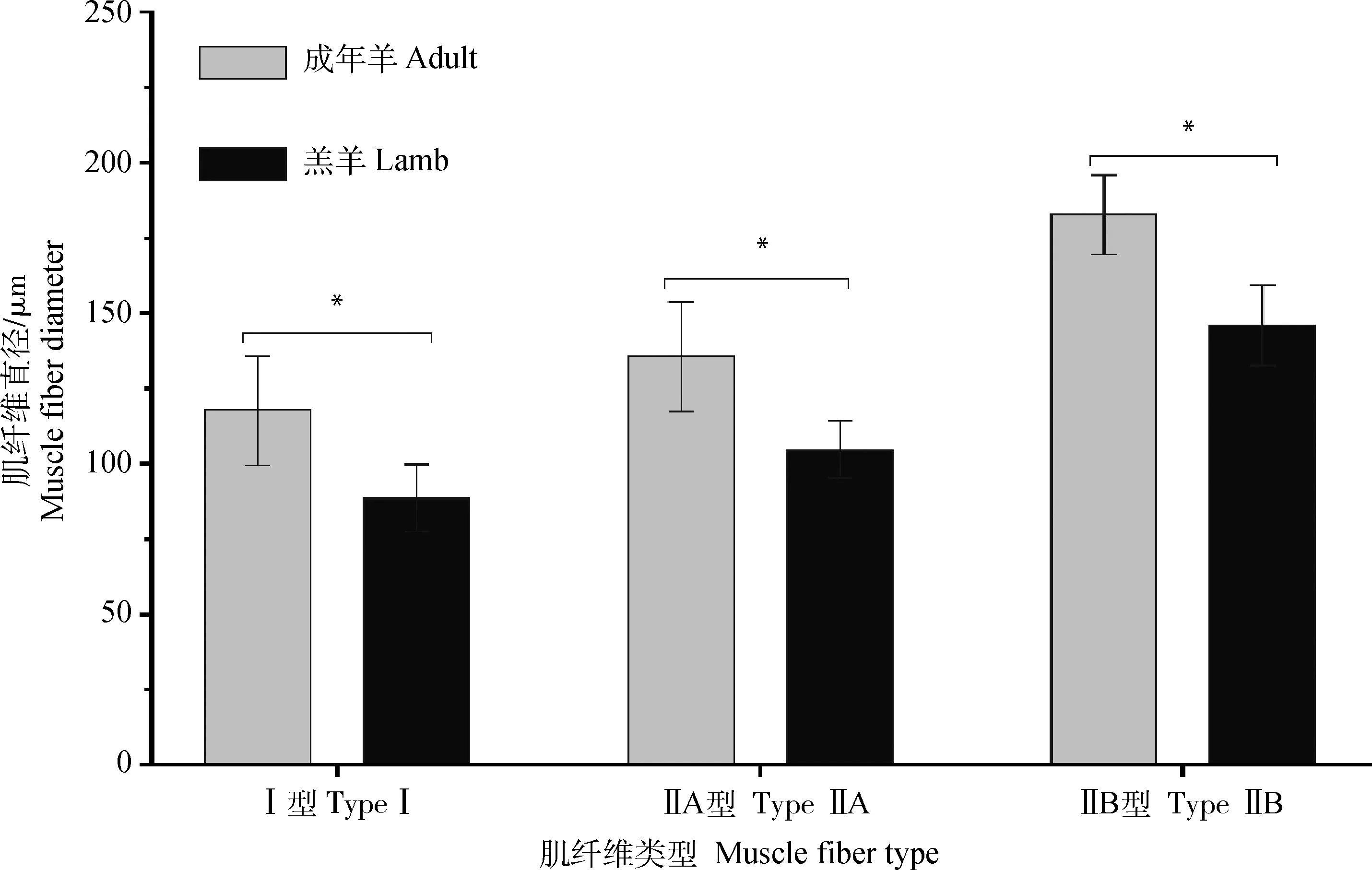
*表示差异显著(P<0.05),下同
肌纤维横截面积分析结果如图2所示。成年羊背最长肌肌纤维横截面积Ⅰ型、ⅡA型和ⅡB型均显著高于初生羔羊(P<0.05)。由此可知,不同发育时期对黑藏羊肌纤维横截面积Ⅰ型、ⅡA型和ⅡB型具有明显的影响。
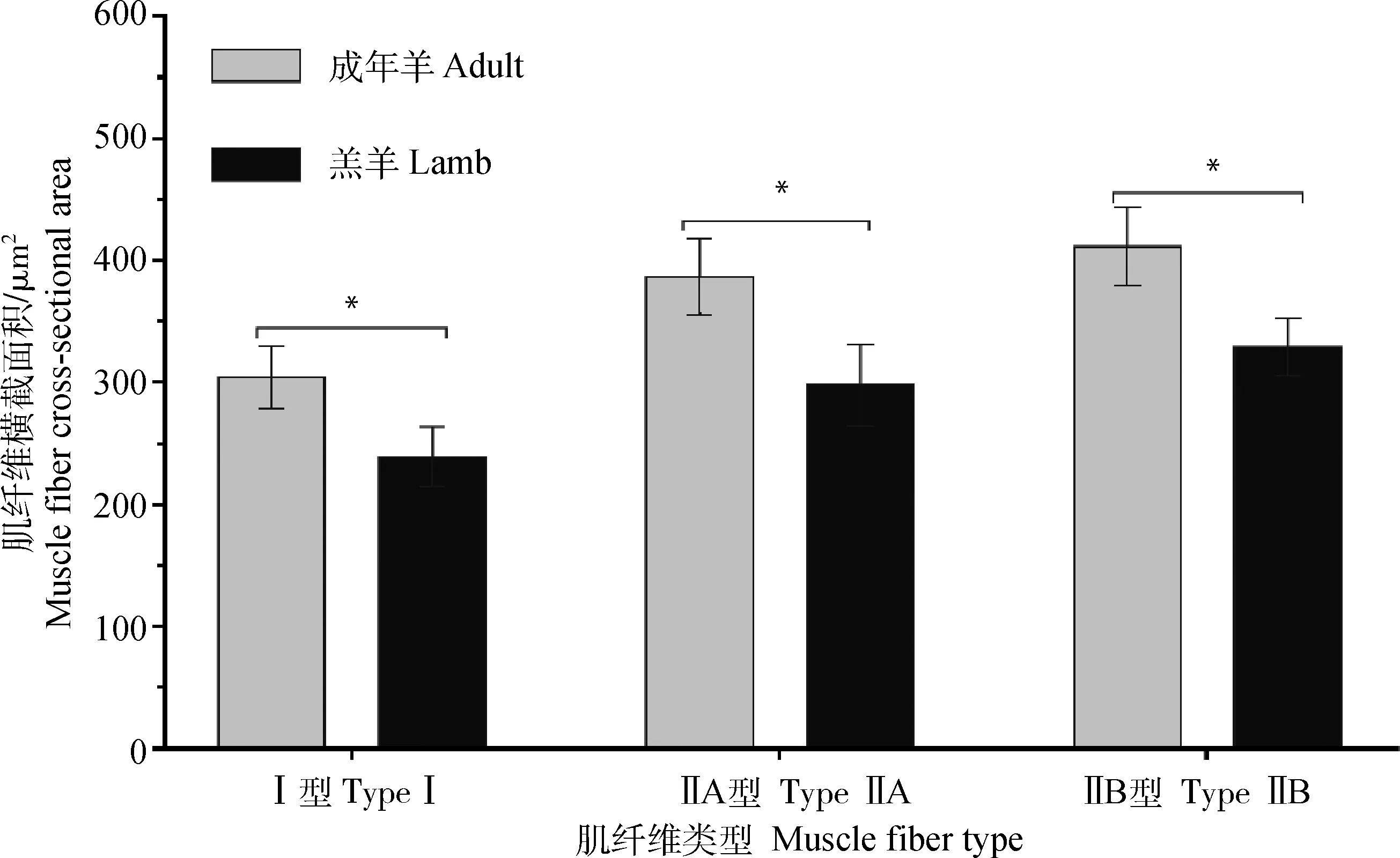
图2 不同生长期黑藏羊背最长肌肌纤维横截面积的比较
肌纤维密度分析结果如图3所示。初生羔羊背最长肌肌纤维密度Ⅰ型、ⅡA型和ⅡB型均高于成年羊,其中2组间肌纤维密度Ⅰ型、ⅡA型和ⅡB型差异显著(P<0.05)。由此揭示,不同生长时期黑藏羊肌纤维密度Ⅰ型、ⅡA型和ⅡB型存在一定的差异。

图3 不同生长期黑藏羊背最长肌肌纤维密度的比较
由表4可知,初生羔羊背最长肌Ⅰ型肌纤维数量比例显著高于成年羊(P<0.05)。2组间黑藏羊背最长肌肌纤维ⅡA型和ⅡB型差异均不显著(P>0.05)。据此可见,随着黑藏羊年龄的增长,肌肉中Ⅰ型肌纤维数量比例会明显减少。

表4 不同生长期黑藏羊背最长肌肌纤维数量比例的比较
由表5可知,成年羊背最长肌Ⅰ型和ⅡA型肌纤维面积比例显著低于初生羔羊(P<0.05)。2组间黑藏羊背最长肌肌纤维ⅡB型差异不显著(P>0.05),据此可知,随着黑藏羊年龄的变化,肌肉中Ⅰ型和ⅡA型肌纤维面积比例会明显降低。

表5 不同生长期黑藏羊背最长肌肌纤维面积比例的比较
2.4 不同生长期黑藏羊背最长肌MyHCs基因表达规律
黑藏羊MyHCs基因的表达水平如图4所示,羔羊背最长肌中MyHCⅠ和MyHCⅡA基因的表达量显著低于成年羊(P<0.05),而MyHCⅡB基因的表达量则呈现相反的趋势(P<0.05)。以上结果说明,不同生长期黑藏羊肌肉中MyHCs基因表达水平差异明显。

图4 黑藏羊MyHCs基因的表达量(n=5)
2.5 MyHCs基因表达量与肉品质的相关性分析
通过相关性分析发现(表6),水分、灰分、总抗氧化能力、过氧化氢酶、超氧化物歧化酶与MyHCⅠ、MyHCⅡA正相关;与MyHCⅡB负相关,而丙二醛则表现相反趋势。粗脂肪、谷胱甘肽过氧化物酶与MyHCⅠ、MyHCⅡA之间正相关,其中与MyHCⅠ显著正相关(P<0.05),但与MyHCIIB负相关。粗蛋白质与MyHCⅠ、MyHCⅡA之间正相关,其中与MyHCⅡA呈显著正相关(P<0.05),但与MyHCⅡB负相关。以上结果表明,黑藏羊背最长肌肌纤维特性与肉品质是密切相关的。

表6 MyHCs表达量与肉品质的相关性分析
3 讨 论
3.1 不同生长期黑藏羊背最长肌常规营养成分的比较
水分、粗蛋白质、粗脂肪、矿物质和维生素等均是羊肉常规营养成分,其组成和含量是营养价值的直接体现[14]。其中水分含量与肌肉的多汁性关系密切,若水分含量越高,口感就越好,机体更易消化吸收。灰分含量越多,意味着肌肉中所含矿物质元素和部分微量元素的含量也就越高[15]。本试验中,羔羊和成年羊背最长肌中水分和灰分含量差异不显著。说明随着黑藏羊年龄的增长,肌肉的多汁性以及矿物质含量没有发生明显的变化。粗蛋白和粗脂肪的含量直接决定着肌肉的营养价值[16]。研究发现,3岁羊肌内脂肪含量极显著高于4月龄羔羊[17]。其他试验也表明,随着湖羊年龄的增长,肌肉中除水分降低外,粗脂肪、粗蛋白质、粗灰分、钙和磷的含量逐渐增加,且成年羊与初生羔羊和断奶羔羊间差异显著[18]。孟凡珠等[19]也证实,在天然放牧条件下,随着年龄的增长,苏尼特羊臂三头肌、股二头肌和臀肌粗脂肪含量均显著提高,增加了新鲜肉的风味。以上研究结论表明,年龄对肌肉粗脂肪以及粗蛋白含量具有明显的影响。而本试验却发现不同生长时期黑藏羊肌肉中粗蛋白与粗脂肪含量差异不显著,这与上述研究结论不一致,究其原因可能是品种、饲养环境、放牧时间以及屠宰年龄等不同造成的。
3.2 不同生长期黑藏羊背最长肌抗氧化能力的比较
一般而言,机体内自由基的产生和清除处于动态平衡中。而当自由基生成量增加或自由基清除能力下降时,则发生氧化应激反应[20]。自由基能够使组织器官受到氧化损伤,降低机体免疫力,从而导致疾病发生[21]。因此,测定肌肉中抗氧化能力,能够反映黑藏羊健康状况,为其不同生长发育阶段提供理论参考。一般认为总抗氧化能力、谷胱甘肽过氧化物酶、超氧化物歧化酶、过氧化氢酶以及丙二醛等是衡量机体抗氧化能力强弱的重要标志[22]。其中总抗氧化能力是衡量机体抗氧化系统的综合性指标[23],其含量可以直接衡量黑藏羊肌肉中抗氧化防卫能力。谷胱甘肽过氧化物酶能够清除活性氧自由基,作为体内抗氧化防御系统的主要组成部分[24]。超氧化物歧化酶作为保护细胞膜结构完整、功能完整一种重要的酶[25]。过氧化氢酶可以催化细胞内过氧化氢分解,防止机体发生损伤[26]。而丙二醛能够降低机体抗氧化酶的活性,反映机体脂质氧化的程度[27]。丙二醛值越低说明肌肉脂质氧化程度越低,抗氧化能力就越强[28]。同时肌肉丙二醛含量也可评价生肉新鲜程度的重要指标。从本试验来看,羔羊和成年羊背最长肌抗氧化能力差异不显著。由此可知,随着黑藏羊年龄的增长,其抗氧化效果并未发生明显的改变,这种现象在韩国本地肉牛[29]也有类似的报道。
3.3 不同生长期黑藏羊背最长肌组织形态学特性的比较
肌纤维作为骨骼肌组织的基本组成单位,其类型组成的差异是影响肌肉品质的关键因素之一[30]。由此可见,骨骼肌的生长发育与动物肌纤维有着密切的联系。其生理生化特性的差异以及代谢的特征从而直接影响动物屠宰后肌肉的质量。当酵解型肌纤维含量高时会使肉的保水性下降,不利于改善肉质[31]。王莉[32]试验发现,牦牛肉Ⅰ型肌纤维比例与剪切力呈负相关关系,即Ⅰ型肌纤维比例越高,剪切力越低,嫩度也就越好。在新疆褐牛与安格斯牛研究表明[33],3、7、12、24月龄新疆褐牛Ⅰ型肌纤维密度随着年龄的增大而减少,安格斯牛Ⅰ型肌纤维密度的变化趋势与新疆褐牛相同,但ⅡB型肌纤维密度随着年龄变化呈现无规律变化。此外发现,1~6岁伊犁马随着年龄的增大,三角肌及背阔肌部位肌纤维横截面积、平均直径增大,单位面积内肌纤维的个数随之减少,失水率变小,剪切力增加[34]。其他试验也进一步证实,犊牦牛背最长肌Ⅰ型和ⅡA型肌纤维直径显著低于成年牦牛,ⅡB型肌纤维也呈现低于成年牦牛趋势,但差异不显著,同时犊牦牛背最长肌Ⅰ型、ⅡA型ⅡB型肌纤维横切面积均显著低于成年牦牛,但肌纤维密度则呈现相反趋势[35]。以上研究均说明酵解型肌纤维含量与肌肉品质呈负相关,肌纤维类型受年龄的影响较大。本试验得出黑藏羊在成年前可以降低酵解型纤维的比例,揭示年龄会影响黑藏羊肌纤维类型的转化,从而影响肌肉的生长发育。另外本试验亦发现,粗脂肪和谷胱甘肽过氧化物酶与MyHCⅠ基因表达丰度呈显著正相关,而粗蛋白质与MyHCⅡA基因表达丰度亦呈显著正相关。由此可见,肌纤维组织形态学特性与其肉品质具有紧密的的联系。动物出生后肌纤维数量不再发生改变,但在年龄影响下肌纤维类型却可发生明显的转化[36]。在苏尼特羊上试验发现,MyHCⅠ、MyHCⅡA基因表达量分别与其Ⅰ型和ⅡA型肌纤维的数量比例呈正相关关系,MyHCⅡB表达量与ⅡB型肌纤维的数量比例呈正相关关系[37],本试验结果也验证了这一观点。
4 结 论
综上所述,在本试验条件下,随着年龄的增长,黑藏羊背最长肌肌纤维类型由氧化型纤维(MyHCⅠ和MyHCⅡA)向酵解型纤维(MyHCⅡB)转化,在一定程度上影响了机体的生长发育。
猜你喜欢
中国畜牧杂志(2021年10期)2021-12-03
养殖与饲料(2021年3期)2021-11-30
今日农业(2021年2期)2021-03-19
国际放射医学核医学杂志(2021年10期)2021-02-28
灌篮(2020年36期)2020-05-16
作文·初中版(2019年8期)2019-09-09
散文诗(2017年22期)2017-06-09
中国社区医师(2016年8期)2016-12-20
大众健康(2015年12期)2015-09-10
小说月刊(2014年6期)2014-04-18
