
生物炭对污泥热解液与牛粪共厌氧发酵的影响
2022-03-29 12:45陈德珍
中国环境科学 2022年3期
岳 霞,陈德珍,安 青,钦 佩
生物炭对污泥热解液与牛粪共厌氧发酵的影响
岳 霞1,2,陈德珍1*,安 青1,钦 佩3
(1.同济大学热能与环境工程研究所,上海 200092;2.内蒙古工业大学机械工程学院,内蒙古 呼和浩特 010051;3.南京大学盐生植物实验室,江苏 南京 210023)
利用4种以芦苇和互花米草为原料的生物炭促进含大量毒性有机物的污泥热解液(SSPL)和牛粪的共厌氧发酵,以实现SSPL的降解并促进其产甲烷,同时分析了炭源中无机质的影响.结果表明:与对照组相比,添加生物炭后可将产甲烷高峰日提前4~8d,累计甲烷产量提升78.8%~157.2%;4种生物炭的促进效果与其炭源中无机质的含量呈负相关.生物炭通过吸附SSPL降低其生物有效性、为微生物提供载体和微量元素、促进种间电子转移来提升甲烷产量.炭源中无机质含量越高,生物炭对SSPL的吸附效果越差,厌氧发酵结束后SSPL的残留量越高;炭源中的碱土金属盐比碱金属盐对厌氧发酵过程更有利.
生物炭;污泥热解液;厌氧发酵;无机质
厌氧发酵是最直接、简单的集中封闭化处理有机废弃物的技术[1-2],在厌氧发酵系统中添加生物炭可增加系统碱度、为微生物提供稳定载体、强化微生物种间电子转移以提高有机物的处理效率和增加甲烷产量[3],不同类型的生物炭对厌氧发酵过程的促进效果差异显著[4].
热解技术是实现污泥减量化的有效方法[5].热解液是污泥热解过程的副产物,含有大量焦油等有机物,但含水率高(90%以上[6])、成分复杂而难以利用;此外污泥热解液具有很高的毒性[7],不宜直接排放;利用厌氧发酵的方式处理污泥热解液,既可解决其环境污染问题,又可将其中的有机物转变为可利用的清洁能源.研究发现[8],相比于其他厌氧发酵底物,牛粪的氮素含量高、发酵过程稳定,自身携带的丰富的菌种有利于形成多种微生物群,将污泥热解液与牛粪以小质量比混合厌氧发酵可实现热解液的降解,但热解液的高毒性会抑制牛粪正常的厌氧发酵进程;在系统中添加包括污泥炭在内的生物炭可缓解这种毒性抑制作用[9],然而生物炭来源多样,不同炭源导致生物炭的性质存在较大差异.
由于生物质原料中无机质的广泛存在,生物炭为碳骨架和无机质相互交错的结构[10],碳骨架与无机质的相互作用,共同决定着生物炭的理化性质和利用途径.木质生物炭是无机质含量低的、高品质的生物炭,但其价格昂贵,应用于厌氧发酵过程不经济;以废弃生物质为原料的生物炭用作厌氧发酵系统的促进剂则是较佳选择.本文以4种不同无机质含量的芦苇和互花米草为原料制备生物炭,用于促进污泥热解液和牛粪共厌氧发酵产甲烷.在对比各实验组产甲烷效率、累计甲烷产量和总挥发性脂肪酸变化情况的基础上,探究炭源中的无机质对生物炭性质和毒性有机物厌氧降解的影响机理,为污泥热解液厌氧处置过程中生物炭促进剂的炭源选择提供支持.
1 材料与方法
1.1 实验材料
污泥样品和污泥热解液的制备方法与文献[8]相同.当热解温度为350~550℃时,污泥热解液的产率随热解温度的升高而增加,但增幅不大[11],为便于比较,采用350℃的污泥热解液(350SSPL)与牛粪进行共厌氧发酵.新鲜牛粪取自上海市郊某奶牛场,接种液取自上海市青浦区农业废弃物处理中心.
为探究炭源中无机质的影响,选取淡水芦苇(FW)、海水芦苇(SW)、无机盐含量非常高的互花米草(H)和提取矿质液后的互花米草(DH)残渣为原料制备生物炭,它们同属多年水生的禾本科植物,但无机质含量差异显著.海水芦苇取自上海市崇明岛沿海地带,淡水芦苇取自浙江省西塘镇某湿地,互花米草取自江苏省盐城市,均为冬季刈割后的废弃生物质秸秆;提取矿质液后的互花米草残渣来自南京大学盐生植物实验室[12],简称脱矿米草.4种生物质原料的特性见表1,受生长环境的影响,互花米草和海水芦苇的无机质以碱金属盐为主,而淡水芦苇的无机质则以碱土金属盐为主;与互花米草相比,脱矿米草的无机质中碱金属元素的含量显著降低,但存在大量的Al和Fe元素.4种生物质原料无机质的含量从高到低为:DH > H > SW > FW.
使用固定床热解炉通过分批多次热解的方式获取生物炭,热解过程以N2为载气,流量80mL/min,热解升温速率20℃/min,热解终温600℃,终温停留时间1h.采用这一制炭条件是因为600℃获得的生物炭具有丰富的孔隙结构,既能够吸附焦油,又具有一定的亲水性,能在水体中充分分散.热解结束待固定床冷却至室温后收集4种生物炭,分别记为FWB(淡水芦苇炭)、SWB(海水芦苇炭)、HB(互花米草炭)和DHB(脱矿米草炭).生物炭经研磨过筛后保留1~2mm的颗粒备用.厌氧发酵各原料的性质如表2所示.4种生物炭无机质的含量从高到低为:DHB > HB > SWB > FWB,此顺序与其炭源中无机质含量的顺序一致.4种生物炭中挥发性固体(VS)的含量顺序与其无机质的含量顺序相反.
注:O含量计算方法:C+H+N+O+灰分=100%.
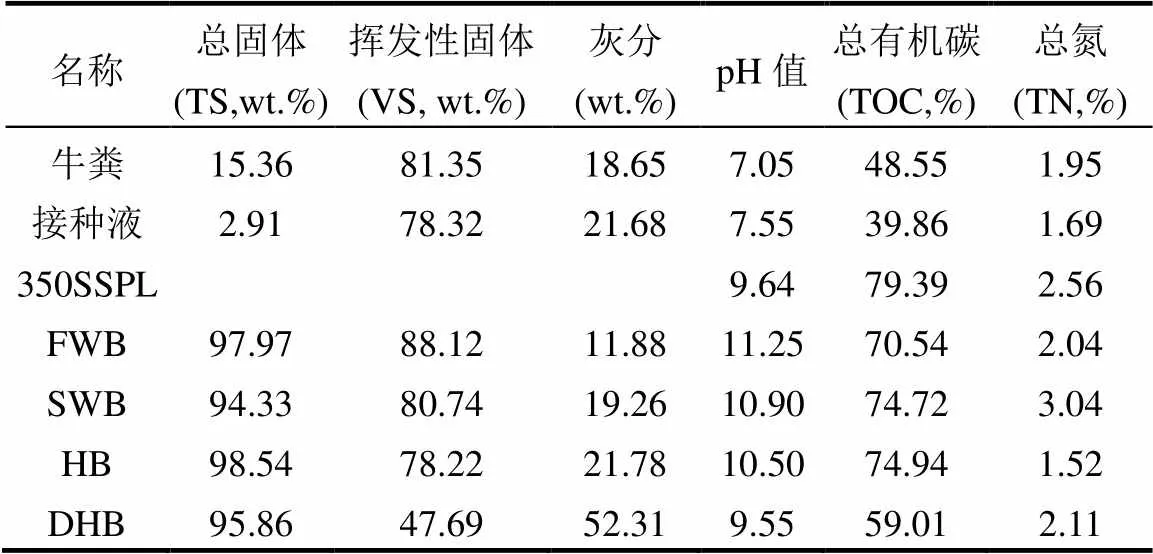
表2 厌氧发酵原料的主要特性
注:350SSPL:污泥350℃热解获得的热解液.
1.2 厌氧发酵实验
厌氧发酵实验共分6组,每组设3个平行反应,各实验组的原料配比如表3所示,由于系统中污泥热解液的浓度较低,为1.25wt.%,因此生物炭的添加浓度选择2.5wt.%,二者质量比为1:2.CD组为牛粪单独厌氧发酵,SSPL+CD组为牛粪和350SSPL共厌氧发酵,这2组未添加生物炭,为对照组.各实验组的C/N值是通过测量原料各组分中的C和N的含量后,根据Mei等[13]的方法计算得出.由于污泥热解液和生物炭均呈碱性,因此各实验组的初始pH值均略偏碱性,尽管常见的厌氧发酵的初始pH值为中性,但也有部分弱碱性发酵的案例[14],并已证明弱碱性环境有助于产生更多的甲烷,因此本实验未调节各实验组的初始pH值.
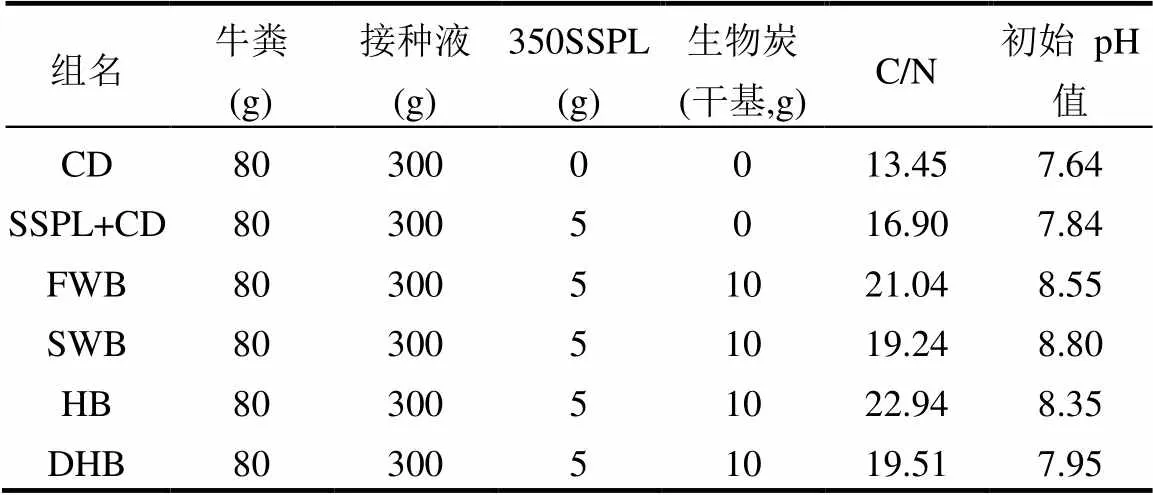
表3 厌氧发酵实验设计
厌氧发酵实验在0.5L的玻璃发酵瓶内进行,顶部密封盖设有出气孔和取样口,出气孔连接气袋.原料按表3的配比称量后装入发酵瓶,充分搅拌混合均匀后,通入氮气2min以驱赶瓶内空气.厌氧发酵瓶密封后放入恒温水浴摇床,摇床温度55℃,转速70r/min.水浴摇床连接有自动补水装置,之后每2日定时检测气体和液体指标,直至产气结束.
1.3 生物炭对污泥热解液的吸附实验
为探明4种生物炭对污泥热解液的吸附能力,以判断各实验组在厌氧发酵初期沼液中污泥热解液毒性抑制作用的强弱,进行了5组吸附实验,每组实验设3个平行反应.第1组向厌氧发酵瓶中添加5g的 350SSPL和340g去离子水,去离子水的添加量是根据表3中的原料配比按照式(1)计算后得到.其他4组在第1组的基础上,再分别添加10g的FWB、SWB、HB和DHB.将厌氧发酵瓶密封后置于水浴摇床中,摇床的设置条件与厌氧发酵实验相同,6d后将厌氧发酵瓶从水浴摇床中取出,混合液经5000r/min离心后,取上清液过滤,测定滤液中总有机碳(TOC)的含量.
去离子水量=牛粪×(1-TS牛粪)+接种液×(1-TS接种液) (1)
1.4 分析方法
工业分析按照《固体生物质燃料工业分析方法》进行[15].元素分析采用Elementer Vario EL Ⅲ 型元素分析仪(BRUKER,德国)测定.样品中无机元素的含量采用Axios-Max型X射线荧光光谱仪(PANalytical,荷兰)测定.生物炭pH值参照《煤质颗粒活性炭试验方法pH值的测定》[16]得到.厌氧发酵过程产生的沼气通过气袋收集后采用BT100-1L型蠕动泵(LongerPump,英国)和LML-1型湿式气体流量计(北京金志业仪器设备有限责任公司,中国)测量体积,采用7820型气相色谱仪(TVFA,安捷伦,美国)测定组分.样品中总挥发性脂肪酸(包括乙酸、丙酸、正丁酸和正戊酸)的含量采用6890N型气相色谱仪(安捷伦,美国)测定.液体中总有机碳(TOC)的含量采用TOC-V CPN 型分析仪(岛津,日本)测得.样品的表面形貌采用Quanta250FEG型场发射扫描电子显微镜(Thermo Fisher Scientific, 美国)进行观察,同时对样品进行能谱分析(EDS),EDS采用Aztec X-MaxN 80型能量色散X射线光谱仪(Oxford Instruments,英国)进行.生物炭的比表面积和孔结构采用Pore Master-33型比表面积及孔径检测仪(Quantachrome,美国)测得,比表面积采用Brunauer–Emmett–Teller (BET)模型计算,孔体积和平均孔径采用Barrett– Joyner–Halenda(BJH)法计算.厌氧发酵结束后的全部残余物用足量的二氯甲烷反复萃取,然后采用QP2010型气相色谱-质谱(GC-MS)联用仪(岛津,日本)分析萃取液中的有机组分,GC-MS分析过程中, 350SSPL的稀释倍数为100倍,发酵残余物的稀释倍数为10倍.
2 结果与讨论
2.1 产甲烷特性
如图1所示,CD组产甲烷良好,具有标准的厌氧发酵规律,即:日甲烷产量逐渐增加,在第6d达到产甲烷高峰后逐渐下降.SSPL+CD组虽有一定量的甲烷产生,但产甲烷高峰日出现在第12d,严重滞后于CD组,并且产甲烷高峰日对应的最高甲烷产量仅约为CD组的1/3,这是因为污泥热解液中的芳香族化合物和糠醛等毒性有机物抑制了厌氧菌细胞间的能量转换,从而降低了厌氧菌的活性[17].对比SSPL+ CD组,添加生物炭的4个实验组的厌氧发酵进程明显加快,其中DHB组将产甲烷高峰日提前到第8d,SWB组和HB组将产甲烷高峰日提前到第6d,与CD组同步,而FWB组将产甲烷高峰日提前到第4d,超前于CD组,这说明生物炭可以快速缓解污泥热解液对微生物的毒性抑制作用,提高厌氧发酵效率.
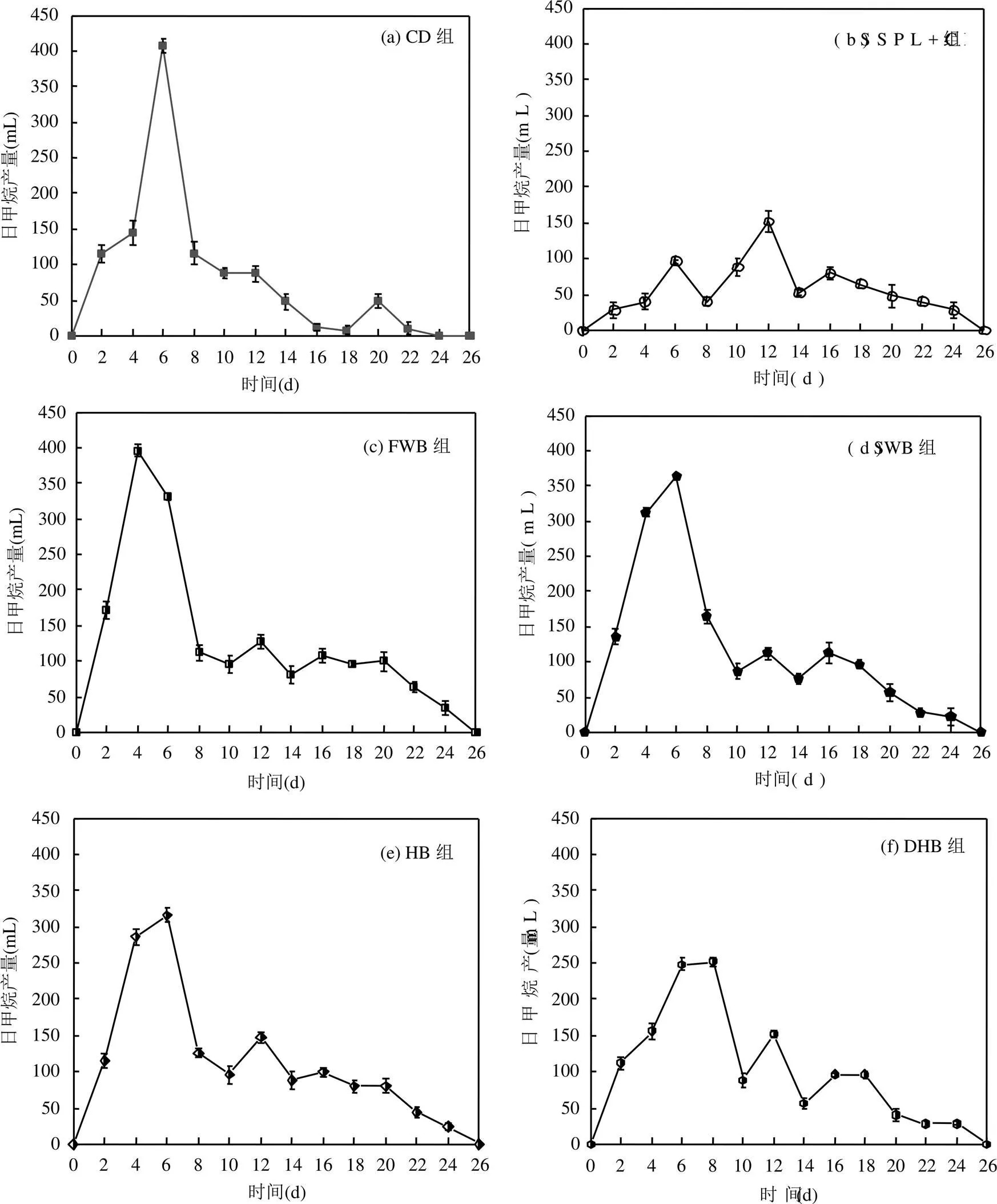
图1 各厌氧发酵实验组的日甲烷产量
如图2所示,与SSPL+CD组相比,FWB组、SWB组、HB组和DHB组的累计甲烷产量分别提高了157.2%、134.1%、125.0%和78.8%;与CD组相比,FWB组、SWB组、HB组和DHB组的累计甲烷产量分别提高了58.2%、44.0%、38.4%和24.49%;说明添加生物炭后,污泥热解液中的有机物得到有效降解并转化为甲烷.添加生物炭的4个实验组的累计甲烷产量的顺序与生物炭中无机质含量的顺序相反,说明无机质含量越低的生物炭促进污泥热解液厌氧发酵产甲烷的效果越好.
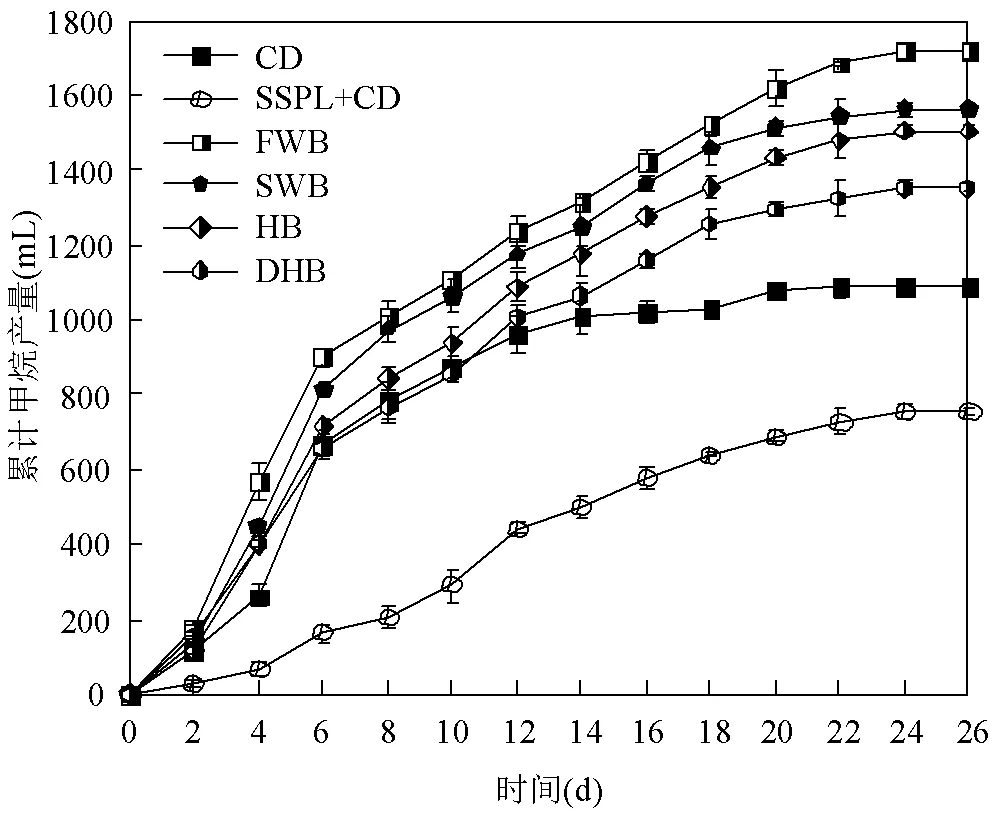
图2 各厌氧发酵实验组的累计甲烷产量
2.2 挥发性脂肪酸含量和组分的变化
如图3所示,根据Cohen等[18]对于厌氧发酵类型的定义判断,CD组前6d为混合型发酵,第8d开始转为丙酸型发酵;CD组第0d(厌氧发酵开始当天)的TVFA含量最高,厌氧发酵开始后波动降低,表明有机物水解后产生的挥发性脂肪酸可被顺利转化为甲烷.SSPL+CD组前10d为混合型发酵,第14d开始转为丙酸型发酵;SSPL+CD组TVFA的含量升高幅度较小,且升高后下降缓慢,结合其日甲烷产量可知,系统中的有机物未被快速水解并转化为甲烷,说明SSPL+CD组微生物的活性较低.添加生物炭的4个实验组从第0d开始即为丙酸型发酵,且各实验组的TVFA含量在第2 ~ 6d迅速升高后又快速降低,结合日甲烷产量可知,其有机物水解迅速且VFA能被微生物快速地转化为甲烷.
对比添加生物炭的4个实验组TVFA的变化情况可知,SWB组和HB组第2 ~ 4d TVFA的含量远高于FWB组,结合FWB组、SWB组和HB组的日甲烷产量可知,SWB组和HB组产生的VFA未被及时转化,存在酸积累现象;DHB组TVFA含量的升高幅度和下降幅度都较低,结合其日甲烷产量可知,系统中有机物的水解速度和VFA转化为甲烷的速度均低于其他3组.
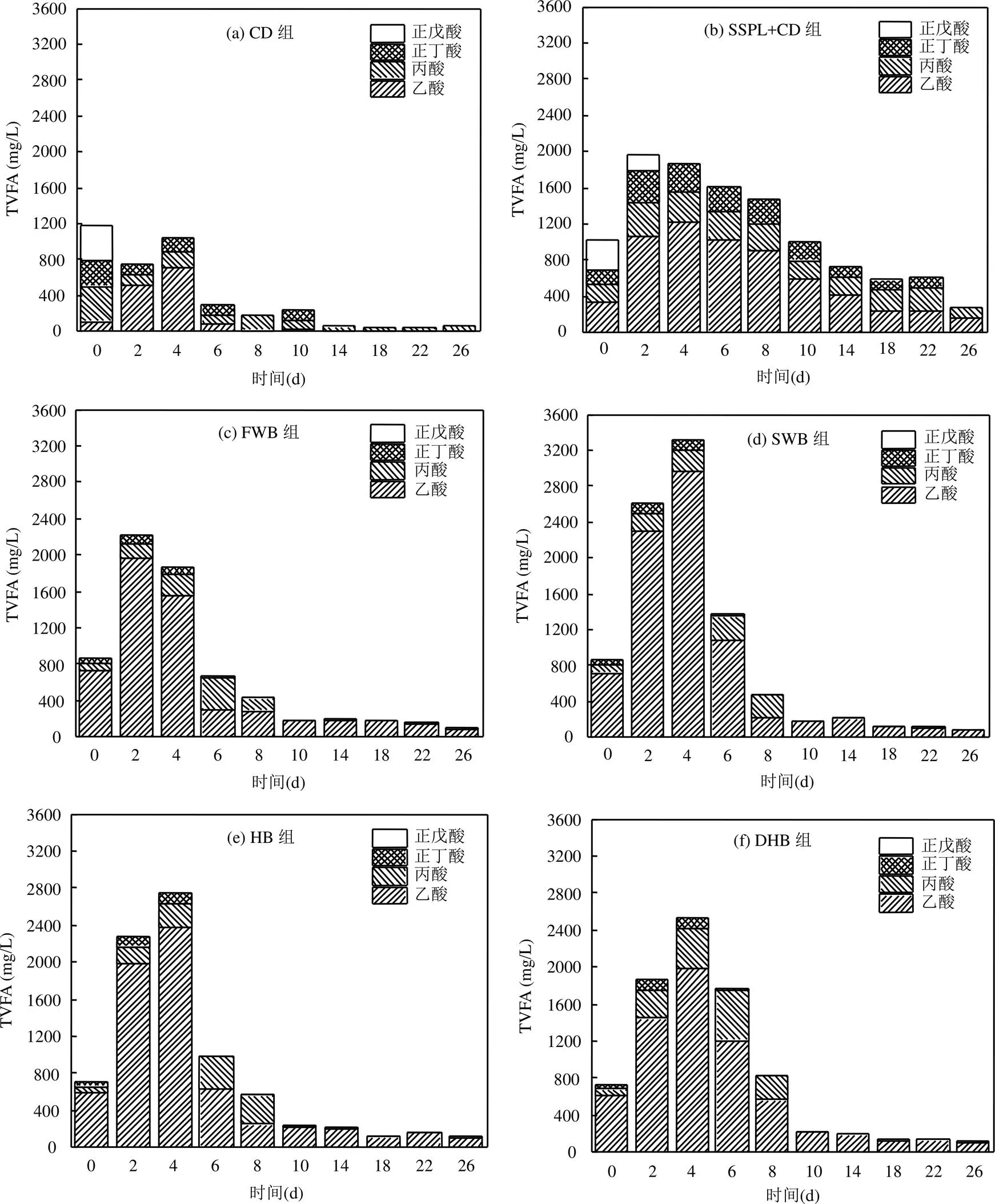
图3 各厌氧发酵实验组总挥发性脂肪酸(TVFA)随时间的变化情况
2.3 炭源中的无机质在生物炭中的转移以及对微生物的影响
如表4所示,除Si外,FWB中的无机质以碱土金属元素为主,SWB和HB中的无机质中以碱金属元素为主,DHB中的无机质以Ca、Fe和Al元素的含量最多,4种生物炭中无机质的元素组成与表1中的炭源一致.
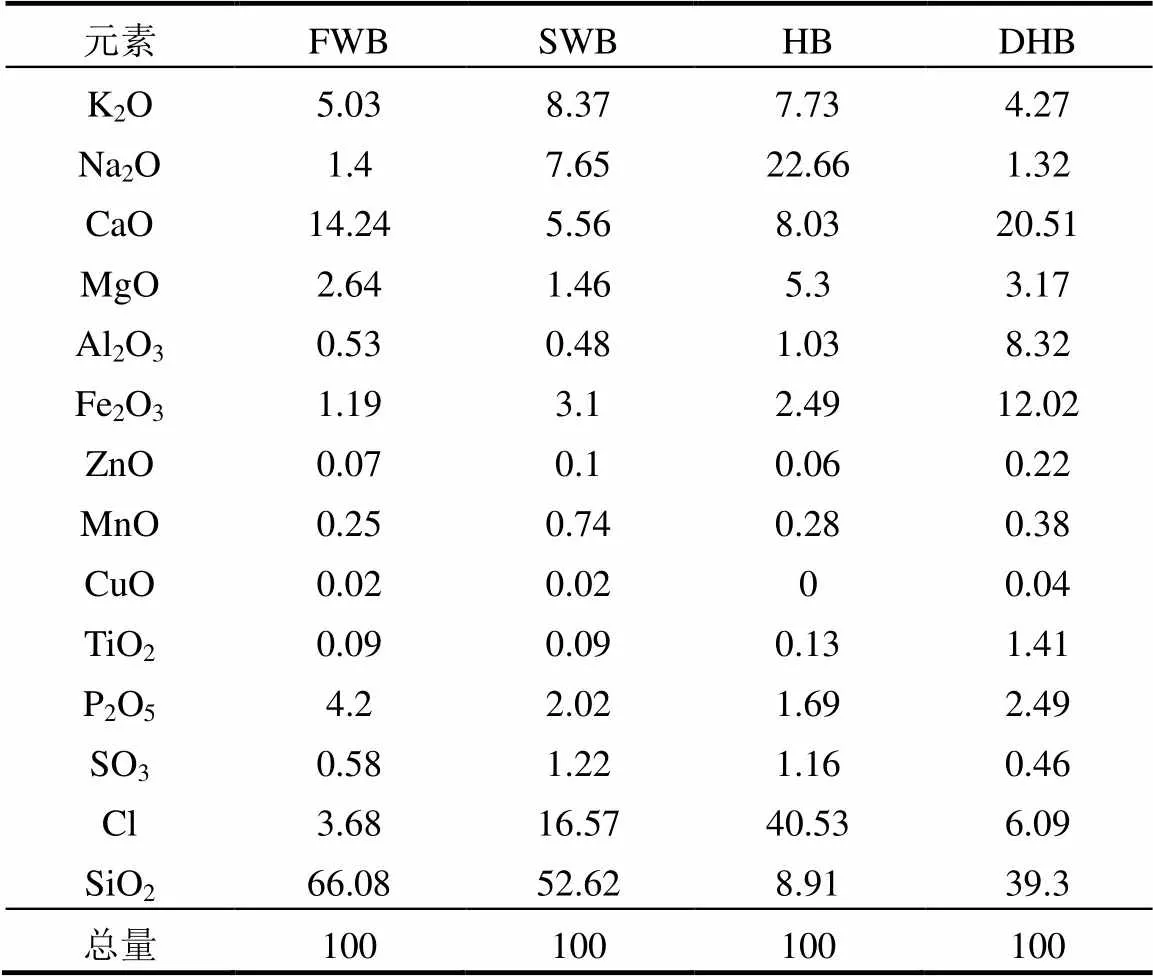
表4 生物炭中无机质的元素组成(%)
生物炭中的无机质可为微生物的生长和代谢过程补充所需的微量元素.研究表明,一定剂量Na能够提高酶活性,调节细胞渗透压和维持膜平衡[19];Ca是产甲烷菌生长过程所需的关键元素,对微生物聚集体的形成具有重要作用[20];Mg能够改善细胞通透性,产生特殊水解酶,提高有机物的水解效率[21];Fe、Cu、Zn是多种产甲烷菌和酶的重要组成成分[22]. Molaey等[23]的研究发现,多种微量元素对微生物的促进作用优于单一种微量元素,微量元素之间具有协同作用,这种协同作用可以提高厌氧系统的稳定性[24],同时可促进微生物种群的增殖和甲烷的形成[25];Nordell等[26]的研究表明,当畜禽粪便作为共发酵基质且含量超过50%时,再额外加入微量元素仍然能够提高系统的甲烷产量;因此生物炭中所含的丰富的无机质是提高厌氧发酵效率、加快甲烷生物合成的重要原因之一.
对比4种生物炭中无机质的元素组成发现, SWB和HB中的碱金属元素,尤其是Na元素的含量远高于FWB和DHB.赵建伟等[27]认为,碱金属盐对微生物的影响主要为Na,过高的Na+质量浓度会破坏细胞内外渗透压甚至造成质壁分离;Kugelman等[28]和Li等[29]的研究表明,厌氧发酵系统中Na+含量过高对微生物的毒害作用表现为Na+和其他离子争夺与蛋白质的结合电位,导致蛋白质活性下降甚至丧失,另外微生物细胞还具有吸Na+和排K+的功能[27];王暾等[30]在单级厌氧发酵装置中研究了NaCl对厨余垃圾厌氧发酵的影响,结果表明甲烷产率随着NaCl质量浓度的增加而降低;王权等[31]的研究也表明, NaCl对厌氧发酵有抑制作用,随NaCl含量的升高系统VFA的酸积累时间延长;因此SWB和HB中Na元素含量较高是导致SWB组和HB组出现酸积累现象(图3)的原因之一,并且从HB组的累计甲烷产量低于SWB组(图2)可以判断,生物炭中Na元素含量越高,对厌氧发酵系统的抑制作用越严重.
2.4 炭源中的无机质对生物炭的比表面积和吸附性能的影响
如表5所示,SWB和HB的比表面积是FWB的2倍以上,这与海水芦苇和互花米草中高含量的碱金属盐在热解过程中对生物炭的造孔作用[32]和对沉积于生物炭孔内的焦油的催化裂解作用有关[33];由于DHB中的无机质(灰分)含量占比超过一半(52.31wt.%),因此DHB的比表面积最小.
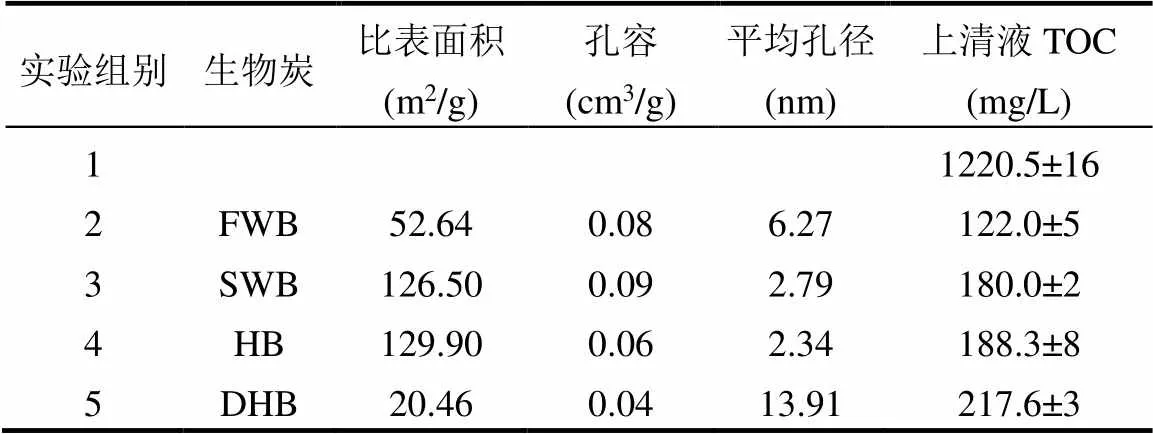
表5 生物炭的比表面积、孔结构和其对污泥热解液的吸附性能
为了比较4种生物炭对污泥热解液的吸附效果,以生物炭吸附污泥热解液后上清液中TOC的含量作为判断依据,TOC含量越低,说明吸附效果越好.对比表5中生物炭的比表面积和上清液TOC的含量可知,生物炭对污泥热解液的吸附能力受其比表面积、孔容和平均孔径的共同影响;虽然SWB和HB具有大的比表面积,但较小的孔容和平均孔径使得二者对污泥热解液中大分子有机物的吸附不利[34]; DHB的平均孔径最大,但其比表面积和孔容均最小,因此对污泥热解液的吸附能力最弱;拥有中等比表面积和较大孔容孔径的FWB对污泥热解液的吸附能力最强.炭源中过高的无机质含量会极大的降低生物炭的品质(如DHB),通过比较炭源中无机质的含量和种类(表1)、生物炭的吸附性能和促进产甲烷性能可以判断:1)炭源的无机质含量越低,生物炭的碳含量越高,孔容孔径较大时对污泥热解液的吸附能力越强;2)炭源中碱金属盐含量越高,生物炭的比表面积越大,但孔容和平均孔径越小,对污泥热解液的吸附能力越弱;3)当炭源中无机质的含量相近时,无机质的组成为碱土金属元素比组成为碱金属更有利于厌氧发酵系统产甲烷(如FWB和SWB),这是因为碱土金属元素可提升厌氧发酵系统的碱度,缓解系统中VFAs大量生成导致的过度酸化,帮助CO2向碳酸氢盐或碳酸盐的转化,从而有效提升系统的甲烷产量[35].
生物炭对污泥热解液的吸附作用使得厌氧发酵初期沼液中毒性有机物的生物有效浓度大幅降低,从而使微生物能够保持活性并快速进入增殖阶段[36].对比图2和表5可知,生物炭对污泥热解液的吸附作用越强,对应实验组的累计甲烷产量越高;Mao等[37]的研究发现,生物炭表面裸露的羰基易与CO2结合,从而间接提高了沼气中甲烷的含量.
研究表明,厌氧菌能够在有生长基质存在时对其他非生长基质进行降解,这种作用称之为共代谢[38].由共代谢作用可以判断,当生长基质(即牛粪)存在时,生物炭吸附的大分子毒性有机物会在厌氧菌的作用下发生氧化、还原、脱烃、脱卤和环裂解等一系列反应,进而逐渐被水解为无毒的小分子有机物并从生物炭上脱附,最终被降解并转化为甲烷;在这个过程进行的同时,厌氧菌逐渐恢复活性并大量增殖,生物炭粗糙的表面和多孔的结构为厌氧菌的生长和繁殖提供了稳定的载体.由图4可观察到在互花米草炭表面存在许多NaCl颗粒;经过厌氧发酵后,大量微生物无序的富集在生物炭的表面和孔隙中,且大部分微生物呈现团聚态,这和Luo等[39]对微生物在生物炭上附着状态的研究一致.厌氧发酵系统中微生物种间快速稳定的电子转移是实现底物降解和甲烷生成的重要途径.生物炭的微生物载体作用和高导电性可缩短共生菌和产甲烷菌之间的距离[40],并提供效率更高的微生物直接或间接的电子传递路径,使电子供体直接或通过生物炭间接的将电子高效的转移到电子受体上,从而有效加速VFA的氧化并促进甲烷的生成[41];但目前对于生物炭中的无机质对其上微生物的富集以及种间电子转移的影响程度还不明确,后续需继续结合微生物菌群特性作进一步的研究.

图4 生物炭厌氧发酵前后的扫描电镜图
2.5 污泥热解液的降解情况
由图5和表6可见,厌氧发酵结束后,SSPL+CD组的残余物中检测到原350SSPL中部分甲苯、乙苯、1-甲基乙基-苯和苯酚残留,它们均属于含苯环的有机物,这说明350SSPL可通过与牛粪共厌氧发酵被降解,但对含苯环类有机物的降解不彻底.
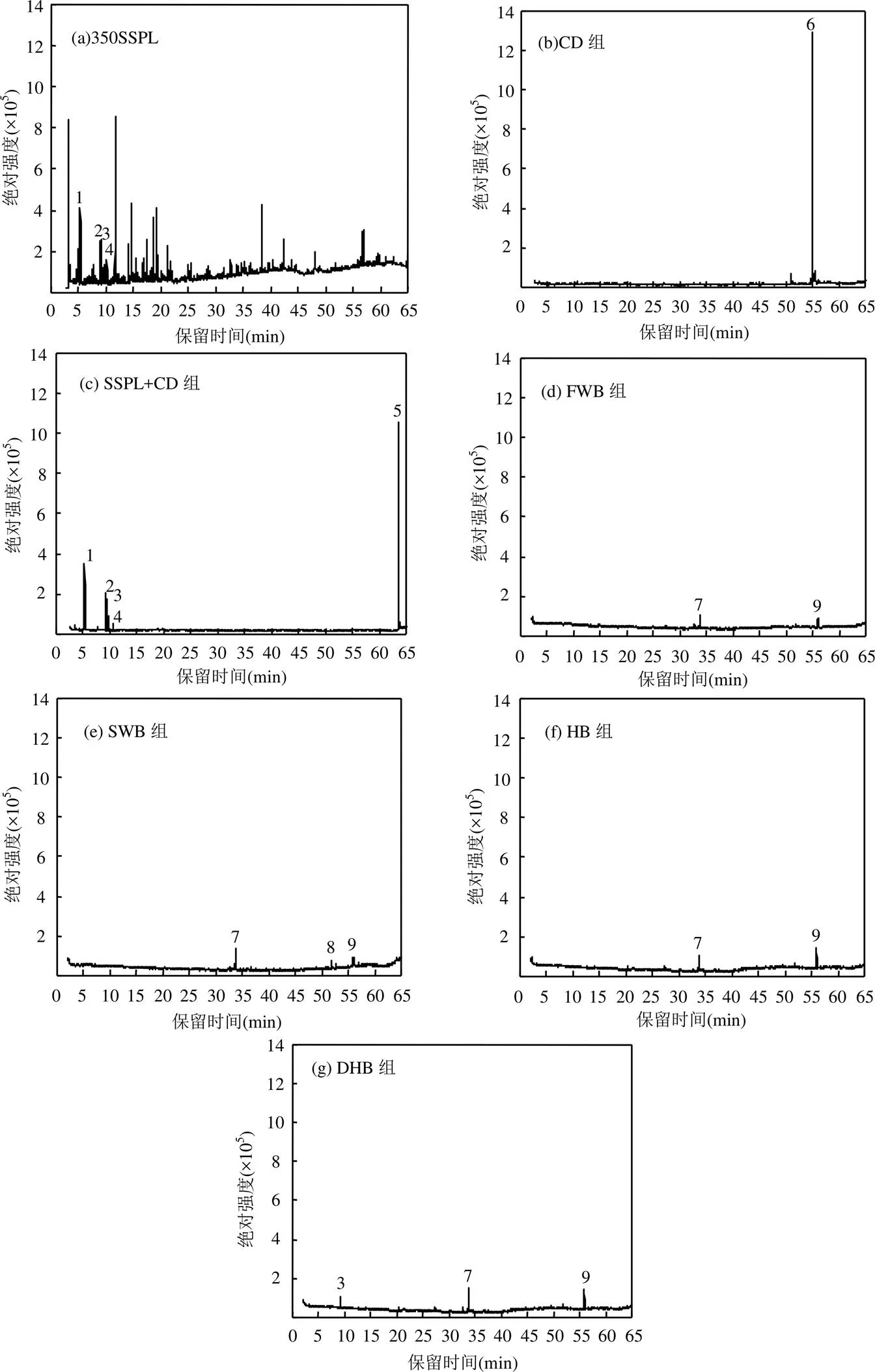
图5 350SSPL和厌氧发酵结束后各实验组残余物的GC-MS图谱
1. 甲苯;2. 乙苯;3. 1-甲基乙基-苯;4. 苯酚;5. 顺-11-二十碳酰胺;6. (Z)-13-二十二碳酰胺;7. 2,4-双(1,1-二甲基乙基)-苯酚;8. 乙酸二十烷基酯;9. 2-羟基-1-(羟甲基)乙酯十六酸
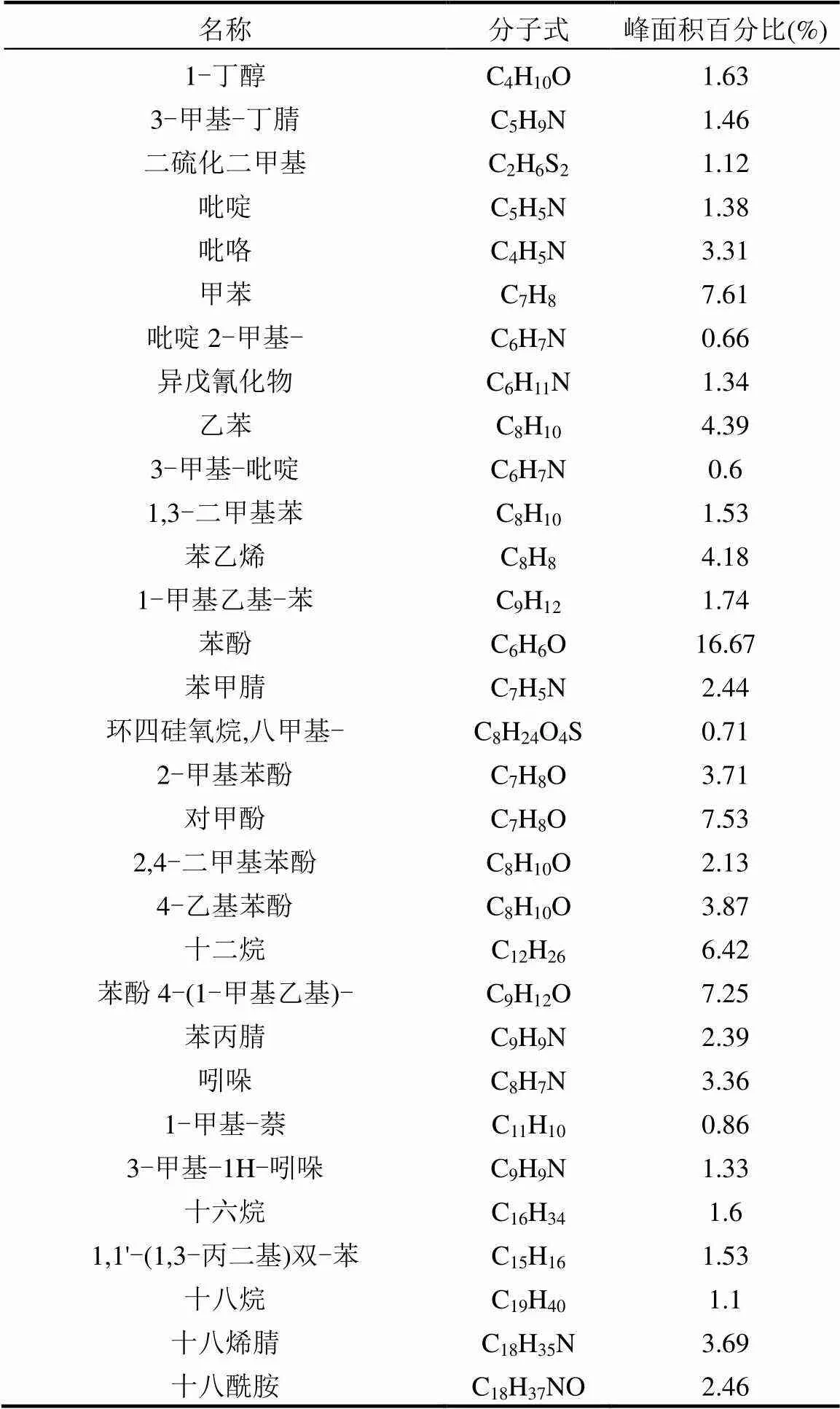
表6 350SSPL中主要的有机组分和各组分含量
添加生物炭后,除DHB组的残余物中依然能检测到少量1-甲基乙基-苯残留外,其他实验组的残余物中未检测到原350SSPL中的有机物,说明添加生物炭可有效促进厌氧发酵系统对含苯环类有机物的降解.厌氧发酵结束后CD组的残余物中存在大量的(Z)-13-二十二碳酰胺,而SSPL+CD组的残余物中也检测到了顺-11-二十碳酰胺,这2种有机物属同类物质,此类物质可能是由牛粪厌氧发酵产生,而添加生物炭的4个组的残余物中均未检测到该类有机物,说明生物炭的添加同样可促进此类物质的降解.添加生物炭的4个实验组中检测到少量的苯酚、酯和乙酯十六酸残留,对比4个组的有机物残留量发现,FWB组最低,DHB组最高,说明炭源中的无机质含量越低,其生物炭对厌氧发酵系统中毒性有机物的去除效果越好.
3 结论
3.1 芦苇和互花米草中的无机质会转移到其对应的生物炭中并影响生物炭对污泥热解液和牛粪共厌氧发酵系统的促进效果.
3.2 水生生物质炭源中的无机质含量越高、生物炭对污泥热解液的吸附能力越弱,促进系统产甲烷的性能越差,厌氧发酵结束后毒性有机物的残留量越高.
3.3 海水芦苇和互花米草中高含量的碱金属元素使得其生物炭的比表面积较大,但孔容和平均孔径较小,因而对污泥热解液的吸附能力较差.
3.4 海水芦苇炭和互花米草炭中高含量的碱金属元素对厌氧发酵过程存在抑制作用.
3.5 水生生物质炭源中无机质含量较低且组成以碱土金属元素为主时对厌氧发酵过程更有利.
[1] Zhang C, Su H, Jan Bet al. Reviewing the anaerobic digestion of food waste for biogas production [J]. Renewable & Sustainable Energy Reviews, 2014,38:383-392.
[2] Pham T P, Kaushik R, Parshetti G Ket al. Food waste-to-energy conversion technologies: current status and future directions [J]. Waste Management, 2015,38:399-408.
[3] 冯 晶,荆 勇,赵立欣.生物炭强化有机废弃物厌氧发酵技术研究 [J]. 农业工程学报, 2019,35(12):256-264.
Feng J, Jing Y, Zhao L X,et al. Research progress on biochar enhanced anaerobic fermentation technology of organic wastes [J]. Transactions of the Chinese Society of Agricultural Engineering, 2019,35(12):256-264.
[4] Pan J, Ma J, Liu Xet alEffects of different types of biochar on the anaerobic digestion of chicken manure [J]. Bioresour. Technol., 2019, 275:258-265.
[5] Song X D, Xue X Y, Chen D Zet al. Application of biochar from sewage sludge to plant cultivation: Influence of pyrolysis temperature and biochar-to-soil ratio on yield and heavy metal accumulation [J]. Chemosphere, 2014,109:213-220.
[6] 李 娜,王建俊,孟记朋.城市污泥热解液相产物分析及焦油加氢精制 [J]. 可再生能源, 2019,37(1):19-25.
Li N, Wang J J, Meng J P, et al. Analysis of pyrolytic liquid phase products of municipal sludge and hydrorefining of tar [J]. Renewable Energy Resources, 2019,37(1):19-25.
[7] Cordella M, Torri C, Adamiano Aet al. Bio-oils from biomass slow pyrolysis: a chemical and toxicological screening [J]. Journal of Hazardous Materials, 2012,231:26-35.
[8] 杨 敏,陈德珍,戴晓虎.污泥热解液与牛粪混合厌氧消化特性研究 [J]. 中国环境科学, 2018,38(2):634-642.
Yang M, Chen D Z, Dai X H. Anaerobic co-digestion of the sewage sludge pyrolysis liquids and cow dung under thermophilic condition [J]. China Environmental Science, 2018,38(2):634-642.
[9] Yue X, U Arena, Chen D,et al. Anaerobic digestion disposal of sewage sludge pyrolysis liquid in cow dung matrix and the enhancing effect of sewage sludge char [J]. Journal of Cleaner Production, 2019, 235:801-811.
[10] Mei Z, He X, Chen Det al. Comparison of chars from municipal solid waste and wheat straw for understanding the role of inorganics in char-based catalysts during volatile reforming process [J]. Energy, 2021,229:120619.
[11] 徐根利,梅振飞,陈德珍.污泥热解-自源炭重整获取高品质油气产物 [J]. 环境工程学报, 2019,13(5):1209-1219.
Xu G L, Mei Z F, Chen D Z. Produce high quality gas & oil products from sewage sludge pyrolysis & char reforming process [J]. Chinese Journal of Environmental Engineering, 2019,13(5):1209-1219.
[12] 刘金珂.互花米草等盐生植物的黄酮含量、抗氧化分析及互花米草黄酮成分鉴定 [D]. 南京:南京大学, 2015.
Liu Jinke. Analysis on the content, antioxidation ability of flavonoids inand some other halophytes, and identification of flavonoids in[D]. Nanjing: Nanjing University, 2015.
[13] Mei Z, Liu X, Huang Xet al. Anaerobic mesophilic codigestion of rice straw and chicken manure: effects of organic loading rate on process stability and performance [J]. Applied Biochemistry and Biotechnology, 2016,179(5):846-862.
[14] Yang L, Huang Y, Zhao Met al. Enhancing biogas generation performance from food wastes by high-solids thermophilic anaerobic digestion: Effect of pH adjustment [J]. International Biodeterioration & Biodegradation, 2015,105:153-159.
[15] GB/T28731-2012 固体生物质燃料工业分析方法 [S].
GB/T28731-2012 Industrial analysis method of solid biomass fuel [S].
[16] GB/T7702.16-1997 煤质颗粒活性炭试验方法 pH值的测定 [S]. 中国:国家技术监督局; 1997.
GB/T7702.16-1997 Test method for granular activated carbon from coal -- Determination of pH value [S].
[17] Fagbohungbe M O, Herbert B M, Hurst L,et al. The challenges of anaerobic digestion and the role of biochar in optimizing anaerobic digestion [J]. Waste Management, 2017,61:236-249.
[18] Cohen B A, Vangemert J M, Zoetemeyer R J. Main characteristics and stoichiometric aspects of acidogenesis of soluble carbohydrate containing wastwater [J]. Process Biochemistry, 1984,19(6):228-232.
[19] 陶冶平,赵明星,阮文权.氯化钠对餐厨垃圾厌氧发酵产沼气影响 [J]. 食品与生物技术学报, 2013,32(6):596-602.
Tao Y P, Zhao M X, Ruan W Q, et al. Effect of sodium chloride on biogas generation of kitchen waste by anaerobic fermentation [J]. Journal of Food Science and Biotechnology, 2013,32(6):596-602.
[20] Murray P A, Zinder S H. Nutritional requirements of methanosarcina sp. Strain TM-1 [J]. Applied and Environmental Microbiology, 1985, 50(1):49-55.
[21] Yadvika, Santosh T R. Sreekrishnan, et al. Enhancement of biogas production from solid substrates using different techniques: a review [J]. Bioresource Technology, 2004,95(1):1-10.
[22] Marcel H. Zandvoort, Eric D. van Hullebusch, Jarno Gietelinget al. Granular sludge in full-scale anaerobic bioreactors: Trace element content and deficiencies [J]. Enzyme and Microbial Technology, 2006, 39(2):337-346.
[23] Molaey R, Bayrakdar A, Sürmeli R Ö, et al. Influence of trace element supplementation on anaerobic digestion of chicken manure: Linking process stability to methanogenic population dynamics [J]. Journal of Cleaner Production, 2018,181:794-800.
[24] Herbert Pobeheim, Bernhard Munk, Harald Lindorfer,et al. Impact of nickel and cobalt on biogas production and process stability during semi-continuous anaerobic fermentation of a model substrate for maize silage [J]. Water Research, 2011,45(2):781-787.
[25] Herbert Pobeheim, Bernhard Munk, Henry Mülleret al. Characterization of an anaerobic population digesting a model substrate for maize in the presence of trace metals [J]. Chemosphere, 2010,80(8):829-836.
[26] Erik Nordell, Britt Nilsson, Sören Nilsson Påledalet al. Co-digestion of manure and industrial waste-The effects of trace element addition [J]. Waste Management, 2016,47:21-27.
[27] 赵建伟.盐度和油酯对餐厨垃圾和剩余污泥厌氧发酵产短链脂肪酸的影响与机理 [D]. 长沙:湖南大学, 2018.
Zhao Jianwei. Effects and mechanisms of salinity, fat, oil and grease (FOG) on short chain fatty acids production from food waste and waste activated sludge anaerobic fermentation [D]. Changsha: Hunan University, 2018.
[28] Kugelman I J, Chin K K. Anaerobic Biological Treatment Processes [M]. United States: American Chemical Socisty, 1971.
[29] Li Y. Stimulation effect of trace metals on anaerobic digestion of high sodium content substrate [J]. Water treatment, 1995,22(10):145-154.
[30] 王 暾.油脂和盐分对餐厨垃圾单级厌氧消化影响的试验研究 [D]. 重庆:重庆大学, 2008.
Wang Tun. Study on effects of grease and salt in single-stage anaerobic digestion of food waste [D]. Chongqing: Chongqing University, 2008.
[31] 王 权,宫常修,蒋建国.NaCl对餐厨垃圾厌氧发酵产VFA浓度及组分的影响 [J]. 中国环境科学, 2014,34(12):3127-3132.
Wang Q, Gong C X, Jiang J G, et al. Effect of NaCl content on VFA concentration and composition during anaerobic fermentation of kitchen waste [J]. China Environmental Science, 2014,34(12):3127- 3132.
[32] Gao Y, Sun R, Li A, et al.self-activation strategy toward highly porous biochar for supercapacitors: Direct carbonization of marine algae [J]. Journal of Electroanalytical Chemistry, 2021,882: 114986.
[33] Yue X, Chen D Z, Luo Jet al. Upgrading of reed pyrolysis oil by using its biochar-based catalytic esterification and the influence of reed sources [J]. Appl. Energy, 2020,268:114970.
[34] Li C, Zhu X, He H, et al. Adsorption of two antibiotics on biochar prepared in air-containing atmosphere: Influence of biochar porosity and molecular size of antibiotics [J]. Journal of Molecular Liquids, 2019,274:353-361.
[35] Wang D, Ai J, Shen F, et al. Improving anaerobic digestion of easy-acidification substrates by promoting buffering capacity using biochar derived from vermicompost [J]. Bioresource Technology, 2017,227:286-296.
[36] Hale S E, Satoshi Endo, Hans Peter H Arp, et al. Sorption of the monoterpenes α-pinene and limonene to carbonaceous geosorbents including biochar [J]. Chemosphere, 2015,119:881-888.
[37] Mao J D, Johnson R L, Lehmann J, et al. Abundant and stable char residues in soils: implications for soil fertility and carbon sequestration [J]. Environmental Science & Technology, 2012,46(17):9571-9576.
[38] Leadbetter E R, Foster J W. Oxidation products formed from gaseous alkanes by the bacterium Pseudomonas methanica [J]. Archives of Biochemistry and Biophysics, 1959,82(2):491-492.
[39] Luo C, Lu F, Shao L, et al. Application of eco-compatible biochar in anaerobic digestion to relieve acid stress and promote the selective colonization of functional microbes [J]. Water Research, 2015,68: 710-718.
[40] Stams Alfons J M. Metabolic interactions between anaerobic bacteria in methanogenic environments [J]. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology, 1994, 66(3):271-294.
[41] Yang Y, Zhang Y, Li Z, et al. Adding granular activated carbon into anaerobic sludge digestion to promote methane production and sludge decomposition [J]. Journal of Cleaner Production, 2017,149:1101- 1108.
Biochars enhancing anaerobic co-digestion of sewage sludge pyrolysis liquid and cow dung: influences of inorganics in biochar raw materials.
YUE Xia1,2, CHEN De-zhen1*, AN Qing1, QIN Pei3
(1.Thermal & Environmental Engineering Institute, Tongji University, Shanghai 200092, China;2.School of Mechanical Engineering, Inner Mongolia University of Technology, Hohhot 010051, China;3.Halophyte Research Laboratory, Nanjing University, Nanjing 210023, China)., 2022,42(3):1267~1277
4 biochars obtained fromandwere added to the anaerobic co-digestion system of SSPL and cow dung matrix. Batch experiment revealed that biochar addition brought forward the peak day of daily methane yield by 4~8 days and increased the cumulative methane yield by 78.8%~157.2% compared with the controlled group without biochar addition. The promoting effects of the biochars were inversely correlated with the inorganic contents in their raw materials. Biochar addition weakened the microbial bioavailability of SSPL in the liquid by adsorbing its toxic compounds, providing supports and trace elements for microbials, and promoting the interspecific electrical transmission of microbials. The higher the inorganic contents in biochar raw materials, the worse the adsorption performances on SSPL of biochars, which resulted in the higher the organic contents in residues after anaerobic digestion. Biochar raw materials containing alkaline earth metals had better promoting effects on anaerobic digestion than alkali metals when their inorganic contents were similar.
biochar;sewage sludge pyrolysis liquid;anaerobic digestion; inorganics
X705
A
1000-6923(2022)03-1267-11
岳 霞(1989-),女,内蒙古呼和浩特人,同济大学博士研究生,主要从事固体废弃物资源化研究.发表论文7篇.
2021-08-09
国家重点研发计划课题(2017YFC0506004)
*责任作者, 教授, chendezhen@tongji.edu.cn
猜你喜欢
煤炭转化(2022年4期)2022-07-14
煤气与热力(2021年12期)2022-01-19
中国沼气(2021年4期)2021-12-15
贵州农业科学(2021年7期)2021-09-13
煤炭转化(2021年4期)2021-07-14
石材(2020年9期)2021-01-07
江苏理工学院学报(2020年2期)2020-10-23
建材发展导向(2019年10期)2019-08-24
中学生数理化(高中版.高考理化)(2019年6期)2019-06-22
农业与技术(2017年3期)2017-03-22
