
厌氧氨氧化脱氮除碳功能菌群结构及代谢途径
2022-03-29 12:40常尧枫郭萌蕾谢军祥谢嘉玮陈重军
中国环境科学 2022年3期
常尧枫,郭萌蕾,谢军祥,谢嘉玮,陈重军,2,3*
厌氧氨氧化脱氮除碳功能菌群结构及代谢途径
常尧枫1,郭萌蕾1,谢军祥1,谢嘉玮1,陈重军1,2,3*
(1.苏州科技大学环境科学与工程学院,江苏 苏州 215009;2.江苏省环境科学与工程重点实验室,江苏 苏州 215009;3.江苏水处理技术与材料协同创新中心,江苏 苏州 215009)
为明确不同有机物浓度(50~150mg/L)和竹炭同时存在下厌氧氨氧化颗粒污泥系统的脱氮除碳功能菌群结构及代谢途径差异,采用宏基因组测序技术对其微生物分布规律和碳氮代谢基因表达进行了研究.结果表明,当COD浓度为50,150mg/L,添加竹炭显著提升了厌氧氨氧化菌(AnAOB)的相对丰度,的相对丰度分别由2.0%和2.9%增加至1.8%和4.5%.与只添加有机物的对照组相比,添加竹炭的处理组显著改变了微生物群落结构,在炭存在下异化硝酸盐还原为铵(DNRA)细菌丰度下降,反硝化菌属和碳代谢相关菌属丰度增加,说明竹炭的投加有助于维持厌氧氨氧化、反硝化和DNRA 3个氮代谢途径群落结构的稳定.微生物共现网络分析表明,不同脱氮菌群的协同作用提高了总氮(TN)的去除率,竹炭可以通过的富集提高和的抗逆性.通过KEGG数据库注释表明,有机物存在条件下竹炭提升了厌氧氨氧化颗粒污泥系统的碳氮代谢效能,尤其促进了糖酵解途径(EMP)与三羧酸循环(TCA cycle)的衔接.
宏基因组;厌氧氨氧化颗粒污泥;脱氮除碳;菌群结构;代谢途径
厌氧氨氧化颗粒污泥结构致密,生物量高,具有良好的污泥处理能力和沉降性能,对复杂的环境条件具有更强的适应性[1].废水中普遍存在大量有机物[2],过高的有机物浓度会对厌氧氨氧化颗粒污泥系统产生不利影响,导致厌氧氨氧化颗粒污泥解体成小颗粒[1,3].当进水COD<100mg/L时,由于反硝化和厌氧氨氧化共存,脱氮效果得到改善[4].而当COD为284mg/L时,厌氧氨氧化活性会被完全抑制[5-7].添加一定量的载体可以促进厌氧氨氧化菌的富集和生长繁殖,在高浓度的有机物(COD 200mg/L)下,与空白组相比,投加7.84mg/L Fe3+可使氨氮和总氮去除率分别提高9.9%和4.9%[6].在UASB反应器中投加竹炭可以加速厌氧氨氧化启动并且显著富集厌氧氨氧化菌[8-9],并且可以在有机物存在的条件下强化厌氧氨氧化颗粒污泥的脱氮效能[10].
目前,在有机物和载体存在下厌氧氨氧化系统脱氮除碳微生物群落的功能组成与代谢差异鲜有报道,宏基因组技术可以在16S rDNA测序的基础上研究微生物的基因与功能.本文利用宏基因组测序技术研究了有机物和竹炭共存条件下厌氧氨氧化颗粒污泥系统微生物群落结构的组成与功能,以功能基因的表达构建了厌氧氨氧化颗粒污泥系统的脱氮除碳代谢路径,以期为有机物存在条件下强化厌氧氨氧化反硝化耦合脱氮除碳提供理论支撑.
1 材料与方法
1.1 研究污泥的来源
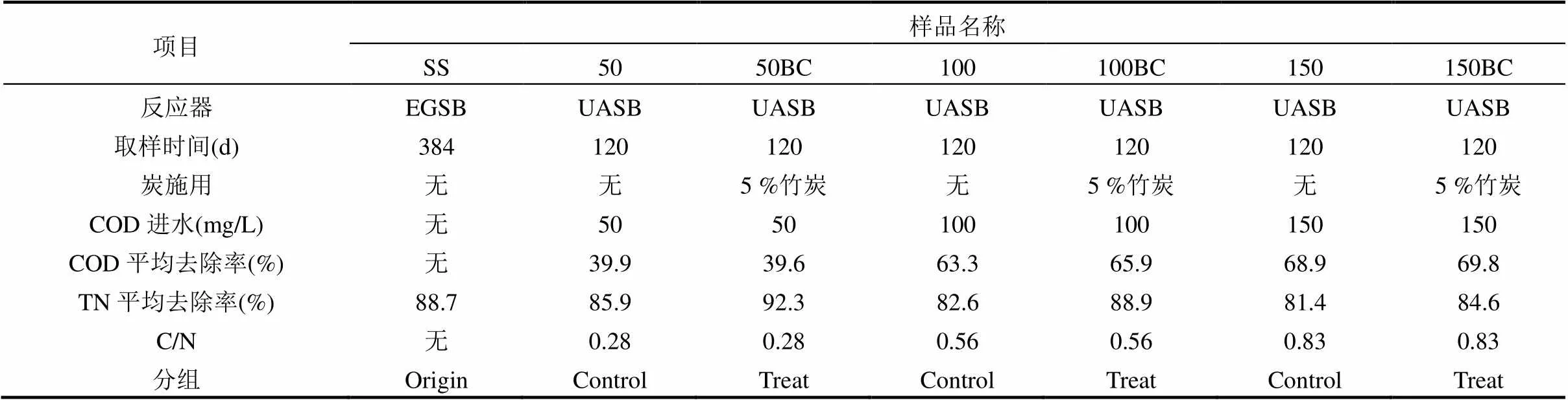
表1 样品情况说明
污泥采集于6个由PVC材质制作的UASB(上流式厌氧污泥床)反应器,进水NH4+-N和NO2--N分别控制在80,100mg/L,水力停留时间(HRT)为2.8h,反应器水浴温度为(32±1)℃.添加葡萄糖作为碳源,设置3个碳源浓度分别为50,100,150mg/L作为对照组,在此基础上再设置3个添加5%体积竹炭(BC)的反应器作为处理组.在反应器运行至120d时采集样品,具体见表1.种泥为厌氧氨氧化颗粒污泥,采集于运行了384d的膨胀颗粒污泥床(EGSB)反应器,运行过程中未添加碳源和竹炭,其氮去除负荷为5.17kg N/(m3·d).7个颗粒污泥样品经过多点采集混合后,均在-70℃保存.具体采样方式为:以8cm为间隔将整个UASB反应器分为5个区域,将采集的5份颗粒污泥样品破碎混合为待测样品.
1.2 DNA提取和宏基因组测序
样品的DNA提取和宏基因组测序均由上海美吉生物医药科技有限公司完成,具体见参考文献[10-11].利用E.Z.N.A.®DNA试剂盒进行样品DNA抽提,利用TBS-380检测DNA浓度,利用NanoDrop200检测DNA纯度,利用1%琼脂糖凝胶电泳检测DNA完整性.桥式PCR扩增后使用Illumina NovaSeq/Hiseq Xten测序平台进行宏基因组测序.利用软件Fastp对原始数据进行质控,利用软件BWA将reads比对到宿主的DNA序列,并去除比对相似性高的污染reads.使用基于succinct de Bruijin graphs原理的拼接软件MEGAHIT对优化序列进行拼接组装.在拼接结果中筛选³300bp的contigs作为最终的组装结果.使用MetaGene对组装出的contig进行ORF预测,然后使用CD-HIT软件对所有样品预测出来的基因序列进行聚类,构建非冗余基因集.最后使用SOAPaligner软件,分别将每个样品的高质量reads与非冗余基因集进行比对(95% identity),统计基因在对应样品中的丰度信息.
1.3 物种与功能注释
利用BLASTP将非冗余基因集序列与NR数据库进行比对,获得物种在Domain(域)、Kingdom(界)、Phylum(门)、Class(纲)、Order(目)、Family(科)、Genus(属)、Species(种)各个分类学水平上的物种注释信息.使用BLASTP将非冗余基因集序列与KEGG的基因数据库(GENES)进行比对[10-11].根本比对结果使用KOBAS 2.0进行功能注释.使用KO、Pathway、EC、Module对应的基因丰度总和计算对应功能类别的丰度[10-11].
2 结果与讨论
2.1 微生物群落结构分析
2.1.1 门水平微生物群落组成 如图1所示,所有样本的微生物群落组成在门水平上主要包括Proteobacteria、Planctomycetes、Chloroflexi、Actinobacteria、Bacteroidetes、Acidobacteria、Ignavibacteria、unclassified_d_Bacteria、Armatimonadetes和Firmicutes.其中厌氧氨氧化菌(AnAOB)隶属于Planctomycetes[12],碳源浓度50, 150mg/L时,与对照组相比,炭处理下Planctomycetes的相对丰度分别提升了2.8%和4.2%,表明两个处理下的AnAOB相对丰度可能有所提升.反硝化菌属一般隶属于Proteobacteria、Firmicutes和Bacteroidetes[13],而异化硝酸盐还原为铵(DNRA)细菌一般存在于Proteobacteria和Bacteroidetes[14].7个样品中Proteobacteria、Firmicutes和Bacteroidetes的相对丰度之和最高的为100BC (71.2%),而其Planctomycetes的相对丰度只占5.1%,表明该处理下异养菌可能是优势菌.从图中聚类树结果可以看出对照组可以聚为一类, 种泥、50BC和150BC可以聚为一类,100BC单独聚为一类,说明50BC和150BC、100和50这两组样品门水平的群落丰度组成具有高度的相似性.
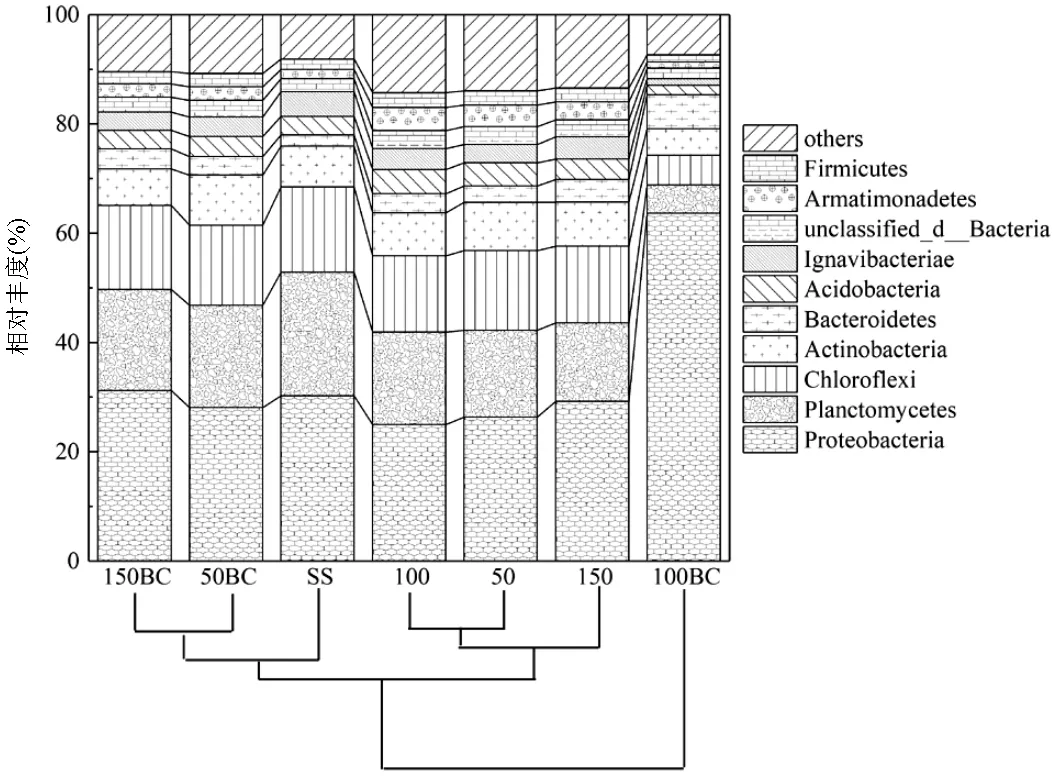
图1 门水平细菌群落结构及分布
2.1.2 属水平微生物群落组成 样品SS、50、50BC、100、100BC、150和150BC分别检出2454、2604、2612、2594、2579、2561和2524种属,有2351种属同时存在于7个样品中.由图2可见,Control组位于第一象限,Origin组位于第四象限,Treat组位于第一、二、四象限,说明在属水平上仅添加葡萄糖的3个样品拥有极高的群落相似度,尽管它们添加葡萄糖的浓度不同,而竹炭的投加对群落组成造成了较大影响.图3中,、及作为AnAOB的优势菌在7个样品均被检出,其中在7个样品中相对丰度都比较小,均未超过0.7%.的相对丰度在添加碳源后急剧下降,50、100、150的相对丰度分别由种泥的8.4%下降至2.0%、2.6%、1.8%,表明对有机物非常敏感,而50BC和150BC的相对丰度下降至2.9%和4.5%,明显高于未投加竹炭的对照组.在碳源存在时下降的幅度相对较小,且50BC和150BC的相对丰度高于其对照组.而在Zhang等[15]的研究中,随着进水C/N的增加,的相对丰度下降而的相对丰度趋于零,这与本研究的结果相反,推测K型菌和B型菌对于有机物的敏感程度取决于两者在接种污泥的初始生态位.本研究中C/N为0.28和0.83时竹炭的投加在一定程度缓解了碳源对和的抑制,C/N为0.56时的结果与之相反.
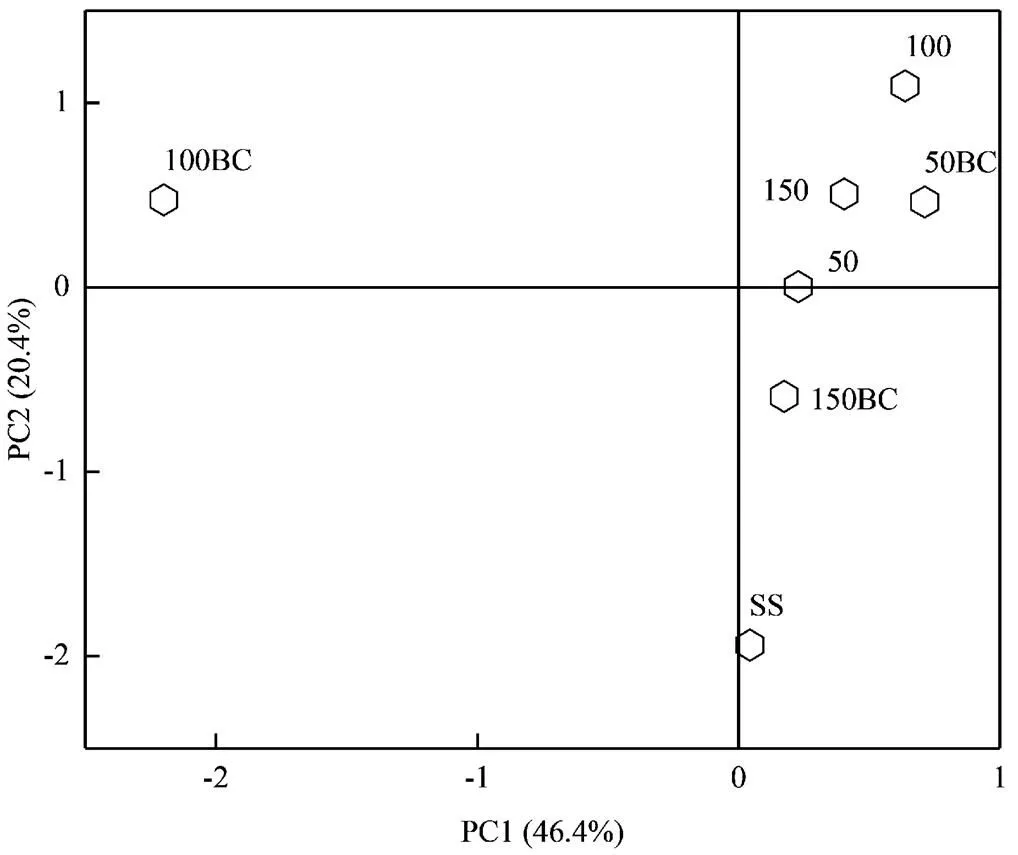
图2 属主成分分析
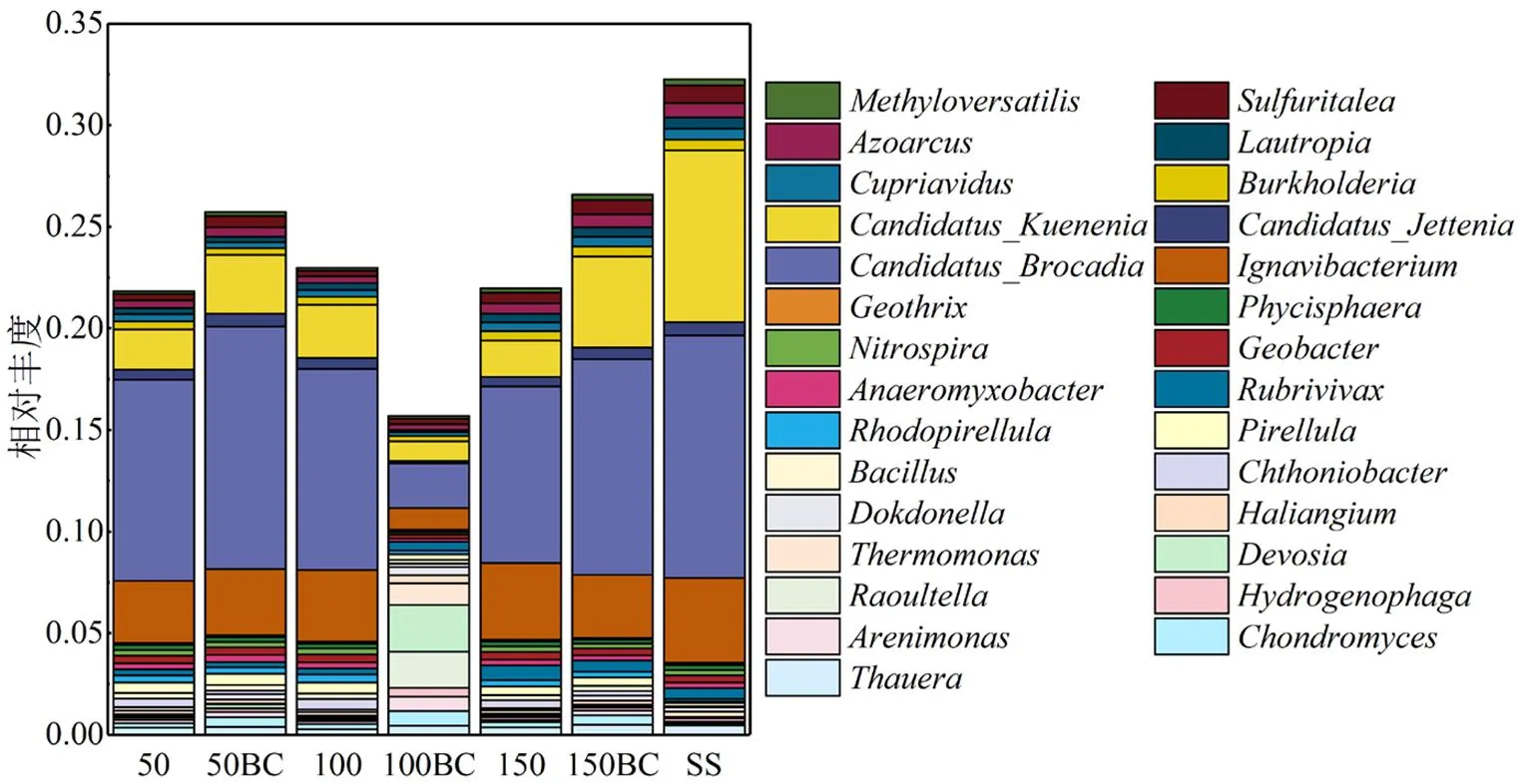
图3 属水平细菌群落结构及分布
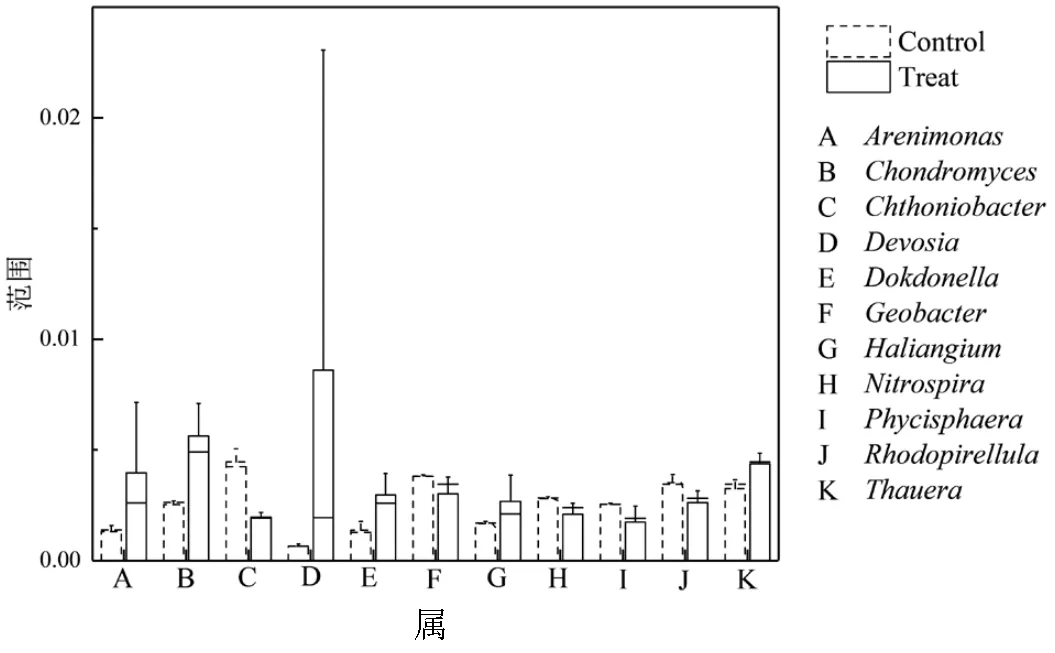
图4 相对丰度显著差异(P<0.05)的主要菌属
图4为29个菌属的差异检验箱线图,一共筛选出11种差异显著的菌属,竹炭处理下、、、和的相对丰度均显著下降,其中[16]和[17]是DNRA菌,[18-19]是一种优势亚硝酸盐氧化菌(NOB),一些属于的微生物还具有全程氨氧化(Comammox)能力.[20]是一种在海洋沉积物中广泛分布的化学异养菌,[21]在某些研究中表明参与了海洋沉积物中厌氧氨氧化耦合硫酸盐还原或三价铁还原过程.在投加竹炭的处理组中、、、、、的相对丰度均高于对照组,其中[22]、[23]、[24]和[25]是反硝化菌,[26]是一种DNRA菌属.[27]是一种粒杆粘细菌属,参与糖酵解和有机酸合成过程.在这11种差异菌属中,DNRA细菌在炭处理下趋向于减少,反硝化菌属和碳代谢相关菌属趋向于增加.而Keren等[28]的研究发现,DNRA菌可能在反应器不稳定的状态下无效复制与_竞争氮源,导致厌氧氨氧化菌效能下降.在有机物存在的条件下,竹炭的加入可能有助于维持厌氧氨氧化、反硝化和DNRA 3个氮代谢途径群落结构的稳定.
2.1.3 微生物共现网络分析 利用R语言的igraph包和Hmisc包对7个样品相对丰度前300的属进行相关性系数的计算,、矩阵阈值分别设置为0.6和0.001,在只考虑正相关的情况下生成对象结构,最后经Gephi进行可视化生成微生物共现性网络图,见图5.整个微生物群落的共发生网络包含191个节点和373条边,网络模块化指数为0.927(高于0.44)[29],证明网络图已经达到了较好的模块化程度.、和连接在一起,[30]是一种可以同时利用外源有机物和内源有机物的反硝化菌属,而[11]是一种硝化菌属,3个关键物种形成的模块表明厌氧氨氧化和硝化反硝化共存于一个平衡的生态位,暗示同一生态位下的物种通过协同作用对强化厌氧氨氧化颗粒污泥系统脱氮除碳起到一定贡献.、和相连接,据报道是一种中度嗜盐菌,具有反硝化作用[31],可以产聚羟基丁酸酯(PHA)[32],而PHA可以保护微生物细胞受极端环境胁迫[33]同时被储存在胞内作为缓释碳源[34].较一般的异养反硝化菌而言,不易被环境扰动从而碳代谢更加稳定,这可能是和的相对丰度在添加碳源的情况下下降幅度较小的原因.
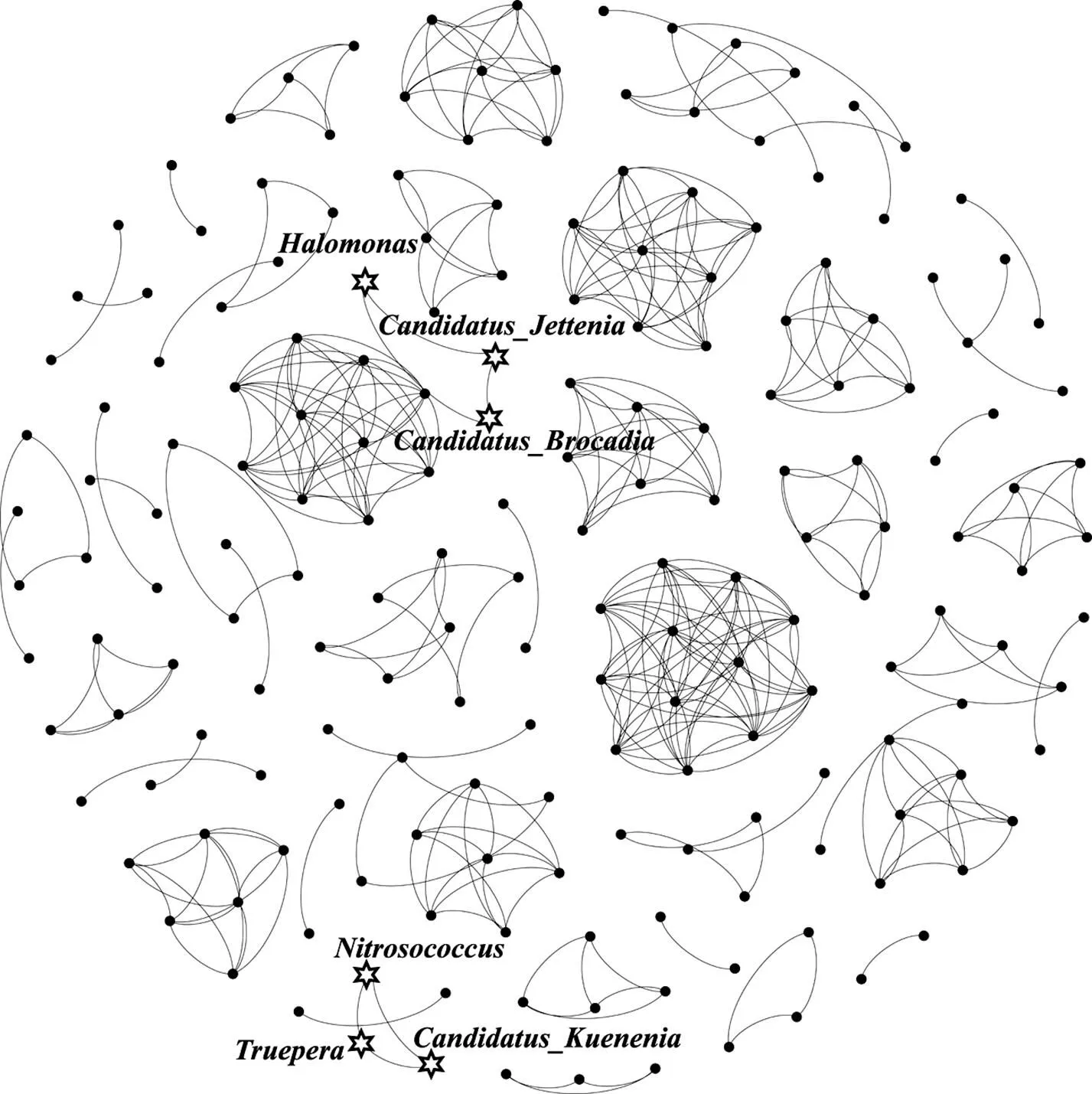
图5 微生物网络分析
2.2 脱氮除碳代谢途径
2.2.1 微生物群落基因组功能 利用KEGG数据库进行代谢通路分析,共注释获得代谢通路405个,共涉及7334个基因.在第1层级通路中,新陈代谢共注释到157条通路,环境信息处理共注释到40条通路,细胞过程共注释到31条通路,其中新陈代谢占整个代谢通路的38.77%.图6为3个通路中筛选出的较为关键的29条第3层级通路,包括氮代谢通路以及11条从属于第2层级的碳水化合物代谢通路.当C/N为0.28和0.83时,热点通路均呈现50BC大于50,150BC大于150,反映了竹炭的投加强化了厌氧氨氧化颗粒污泥系统的整体代谢水平.氮代谢通路,所有炭处理样品的相对丰度均高于未处理组,说明有机物存在的条件下竹炭的投加确实改变了原有的氮代谢途径,这与属水平微生物群落组成中的结论一致.而在碳代谢通路中,氨基糖和核苷酸糖代谢、三羧酸循环、果糖和甘露糖代谢、半乳糖代谢、糖酵解/糖异生和磷酸戊糖途径在样品中呈现50小于50BC,100大于100BC,150小于150BC.虽然当C/N比为0.56时上述关键的糖代谢通路丰度并没有在炭处理下提高,但是鞭毛组装、生物被膜形成-大肠杆菌、生物被膜形成-铜绿假单胞菌、生物被膜形成-霍乱弧菌、ABC转运蛋白、细菌分泌系统和双组分系统等代谢通路得到显著富集,这些通路[35]是细菌在压力环境下传导信号分子并激发相应调控机制的重要通路,这反映竹炭处理下的厌氧氨氧化颗粒污泥系统抗逆性得到了显著增强.
2.2.2 主要碳氮代谢功能基因丰度 编码联氨脱氢酶(hydrazine dehydrogenase, HDH,EC: 1.7.2.8)的基因和编码联氨合成酶(hydrazine synthase, HZS,EC:1.7.2.7)3个亚基的基因只存在于厌氧氨氧化体中[36],如图7所示,当C/N为0.28和0.83时,竹炭促进了和基因的表达,然而C/N为0.56时的结果与之相反.C/N为0.56时样品100BC的基因的表达量是所有样品中最低的,而、和的基因表达量是所有样品中最高的.基因编码的周质细胞色素C亚硝酸盐还原酶(nitrite reductase (cytochrome c-552) ,ccNIR,EC:1.7.2.2)负责催化异化亚硝酸盐还原成氨反应[37],和分别编码一氧化氮还原酶(nitric oxide reductase, EC:1.7.2.5)的两个亚基[38],负责编码一氧化二氮还原酶(nitrous-oxide reductase,EC:1.7.2.4)[38].因此可以得出结论样品100BC中厌氧氨氧化和DNRA途径相关的基因表达量特别低而反硝化过程相关基因表达量却特别高,因竹炭处理下DNRA细菌趋向减少而反硝化菌属趋向增多,因此推测当DNRA途径被过度抑制同时反硝化途径过度表达时,厌氧氨氧化过程的生态位将受到严重压缩.
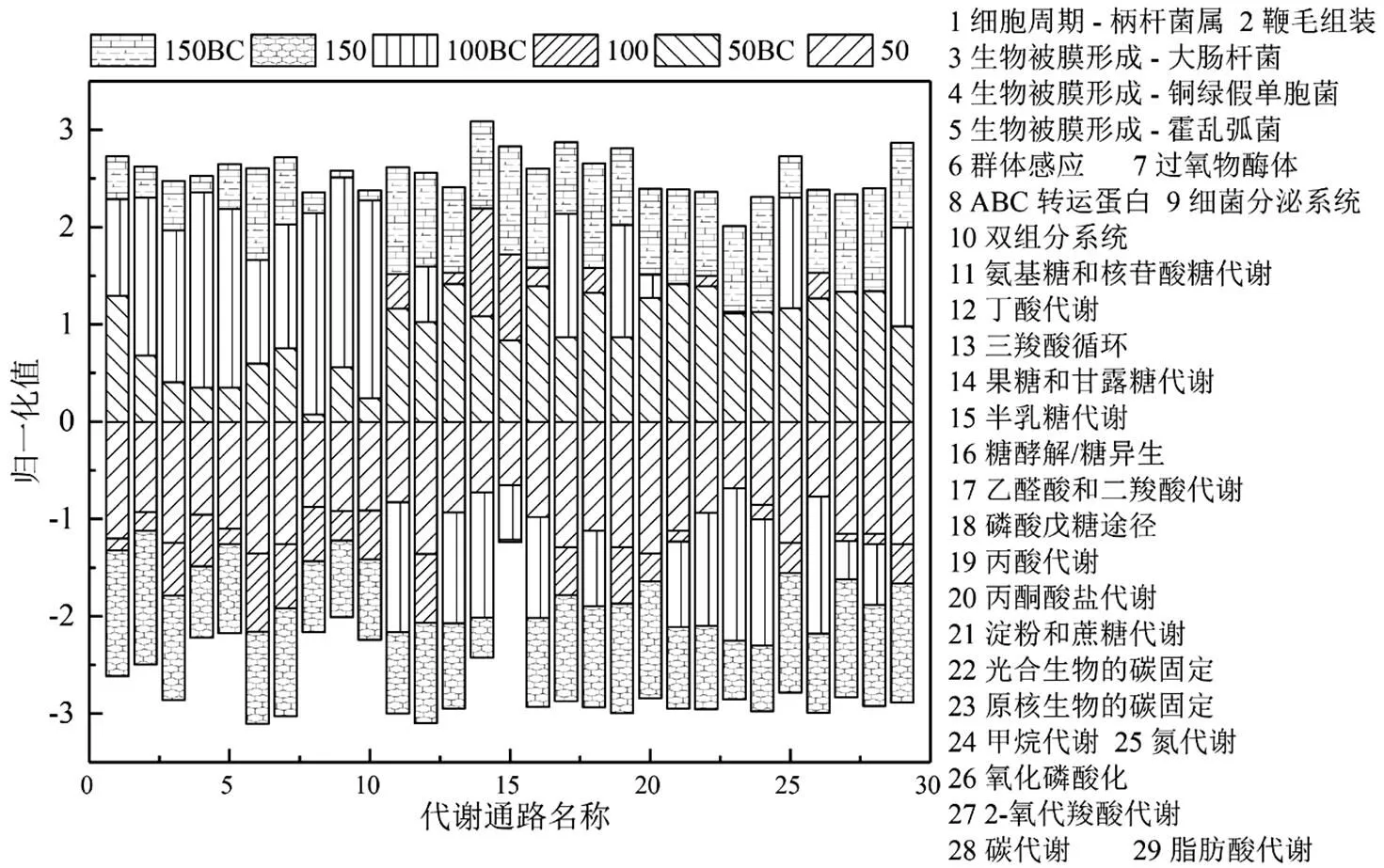
图6 注释到KEGG第3层级的基因数目热图
糖酵解途径(EMP)和三羧酸循环(TCA cycle)是大多数生物所共有的糖分解代谢途径,而且二者通路在炭处理下差异明显.从葡萄糖到丙酮酸共有10步连续的酶促反应,其中3步最主要的限速步骤分别为:葡萄糖在葡萄糖激酶(glucokinase, EC:2.7.1.2)的催化下生成葡萄糖-6-磷酸、果糖-6-磷酸在果糖磷酸激酶(phosphohexokinase, EC:2.7.1.11)催化下生成果糖-1,6-二磷酸以及磷酸烯醇式丙酮酸在丙酮酸激酶(pyruvate kinase, EC:2.7.1.40)的催化下生成丙酮酸,3个反应均为不可逆反应.当C/N比为0.28和0.83时,竹炭的投加显著促进了葡萄糖激酶基因、果糖磷酸激酶基因、丙酮酸激酶基因的表达,而C/N为0.56时和在炭处理下下调.其中丙酮酸脱氢酶系(丙酮酸脱氢酶E1,二氢硫辛酰转乙酰基酶E2,二氢硫辛酰胺脱氢酶E3,EC: 1.2.4.1,EC:2.3.1.12,EC:1.8.1.4)催化的丙酮酸氧化脱羧形成乙酰辅酶A过程是连接EMP和TCA的中心环节(不可逆)[39].丙酮酸脱氢酶系是一个位于线粒体内膜上的多酶复合体,涉及、和3个功能基因,不同有机物浓度下3个功能基因的表达量均表现为处理组大于对照组,说明竹炭有效促进了EMP途径与TCA循环的衔接.柠檬酸合酶(Citrate synthase,EC:2.3.3.1)催化乙酰辅酶A和草酰乙酸缩合合成柠檬酸和辅酶A,该过程控制TCA的入口,是TCA最重要的一个限速步骤[40].
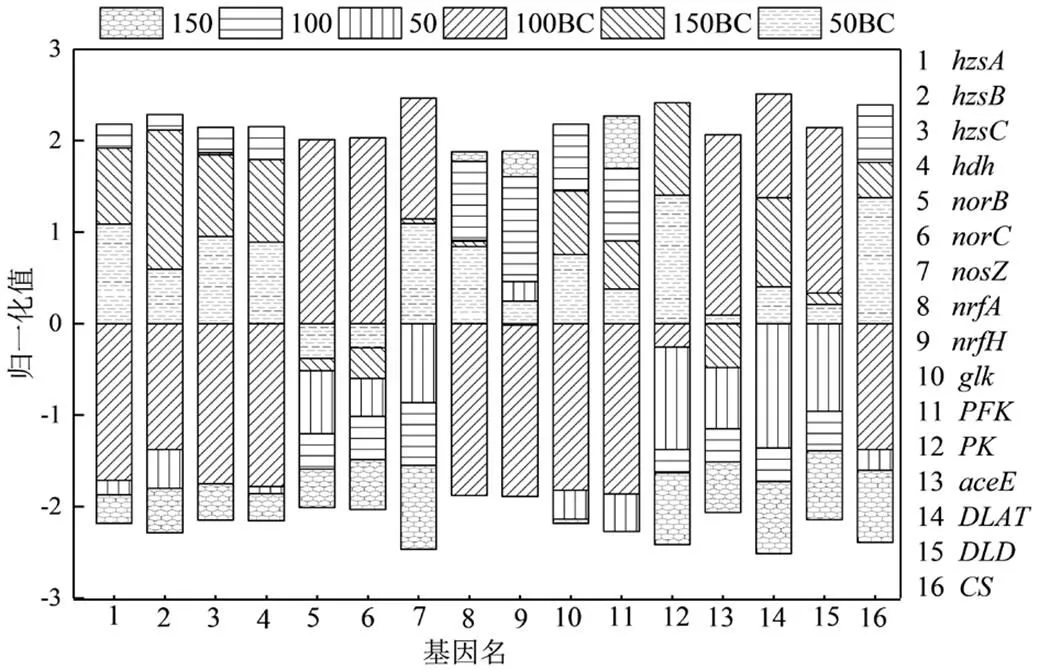
图7 功能基因表达热图
柠檬酸合酶编码基因在各样品中的表达情况为:50BC大于50, 150BC大于150,100BC小于100,这与EMP中葡萄糖激酶基因和果糖磷酸激酶基因的表达趋势一致,说明EMP与TCA的代谢效能呈正相关.总而言之,有机物存在条件下竹炭促进了厌氧氨氧化颗粒污泥系统的碳代谢.
3 结论
3.1 竹炭的投加在一定程度缓解了有机物对的抑制,C/N为0.28和0.83时,对照组的相对丰度分别由种泥的8.4%下降至2.0%和1.8%,而处理组的相对丰度下降至2.9%和4.5%.
3.2 在有机物存在条件下投加竹炭可能有助于维持厌氧氨氧化、反硝化和DNRA三个氮代谢途径群落结构的平衡,从而强化TN去除效果.其中C/N比为0.28时,竹炭处理组的TN平均去除率高达92.3%.
3.3 竹炭和有机物共存时可以通过特定微生物物种的富集提高和的抗逆性,并促进了厌氧氨氧化颗粒污泥系统的碳代谢,尤其是EMP与TCA的衔接.
[1] 郭萌蕾,姜 滢,谢嘉玮,等.厌氧氨氧化颗粒污泥性质及影响因素研究进展 [J]. 化学通报, 2020,83(1):30-34.
Guo M L, Jiang Y, Xie J W, et al. Research progress in properties and influencing factors of anaerobic ammonium oxide granular sludge [J]. China Environmental Science, 2020,83(1):30-34.
[2] 陈重军,朱为静,黄孝肖,等.有机碳源下废水厌氧氨氧化同步脱氮除碳 [J]. 生物工程学报, 2014,30(12):1835-1844.
Chen C J, Zhu W J, Huang X X, et al. Simultaneous removal of carbon and nitrogen from organic-rich wastewater with Anammox [J]. Chin. J. Biotech., 2014,30(12):1835-1844.
[3] 操沈彬,王淑莹,吴程程,等.有机物对厌氧氨氧化系统的冲击影响 [J]. 中国环境科学, 2013,33(12):2164-2169.
Cao S B, Wang S Y, Wu C C, et al. Shock effect of organic matters on anaerobic ammonia oxidation system [J]. China Environmental Science, 2013,33(12):2164-2169.
[4] 陈重军,汪瑶琪,姜 滢,等.有机物对连续流Anammox脱氮及微生物群落影响 [J]. 中国环境科学, 2019,39(12):5049-5055.
Chen C J, Wang Y Q, Jiang Y, et al. Effects of organic matter on nitrogen removal and microbial community in anammox reactor [J]. China Environmental Science, 2019,39(12):5049-5055.
[5] 谢军祥,姜 滢,常尧枫,等.城镇生活污水厌氧氨氧化处理的研究进展 [J]. 化工进展, 2020,39(10):4175-4184.
Xie J X, Jiang Y, Chang Y F, et al. Research progress of anaerobic ammonia oxidation treatment of urban sewage [J]. Chemical Industry and Engineering Progress, 2020,39(10):4175-4184.
[6] 王海月,彭 玲,毛念佳,等.三价铁对有机物存在下厌氧氨氧化脱氮的影响 [J]. 中国环境科学, 2021,41(4):1672-1680.
Wang H Y, Peng L, Mao N J, et al. Effect of Fe3+on nitrogen removal of Anammox in the presence of organic matter [J]. China Environmental Science, 2021,41(4):1672-1680.
[7] 杨京月,郑照明,李 军,等.厌氧氨氧化耦合反硝化底物竞争抑制特性 [J]. 中国环境科学, 2018,38(8):2947-2953.
Yang J Y, Zheng Z M, Li J, et al. The inhibitive characteristics by substrate competition of the anammox coupling denitrification process [J]. hina Environmental Science, 2018,38(8):2947-2953.
[8] Chen C J, Huang X X, Lei C X, et al. Improving anammox start-up with bamboo charcoal [J]. Chemosphere, 2012,89(10):1224-1229.
[9] Adams Mabruk, Xie J X, Xie J W, et al. The effect of carrier addition on Anammox start-up and microbial community: a review [J]. Reviews in Environmental Science and Bio/Technology, 2020,19(2): 355-368.
[10] Guo M L, Jiang Y, Xie J X, et al. Bamboo charcoal addition enhanced the nitrogen removal of anammox granular sludge with COD: Performance, physicochemical characteristics and microbial community [J]. Journal of Environmental Sciences, 2022,115:55-64.
[11] He X Q,Yin H J, Fang C, et al. Metagenomic and q-PCR analysis reveals the effect of powder bamboo biochar on nitrous oxide and ammonia emissions during aerobic composting [J]. Bioresource Technology, 2021,323:124567.
[12] Podder Aditi, Reinhart Debra, Goel Ramesh. Nitrogen management in landfill leachate using single-stage anammox process-illustrating key nitrogen pathways under an ecogenomics framework [J]. Bioresource Technology, 2020,312:123578.
[13] Dang Bao-Trong, Bui Xuan-Thanh, Itayama Tomoaki, et al. Microbial community response to ciprofloxacin toxicity in sponge membrane bioreactor [J]. Science of The Total Environment, 2021,773: 145041.
[14] Broman Elias, Zilius Mindaugas, Samuiloviene Aurelija, et al. Active DNRA and denitrification in oxic hypereutrophic waters [J]. Water Research, 2021,194:116954.
[15] Zhang X Y, Liu Y, Li Z R, et al. Impact of COD/N on anammox granular sludge with different biological carriers [J]. Science of The Total Environment, 2020,728:138557.
[16] Sun F, Deng Q H, Li X W, et al. Organic carbon quantity and quality jointly triggered the switch between dissimilatory nitrate reduction to ammonium (DNRA) and denitrification in biofilters [J]. Chemosphere, 2021,280:130917.
[17] Yu T T, Li M, Niu M Y, et al. Difference of nitrogen-cycling microbes between shallow bay and deep-sea sediments in the South China Sea [J]. Applied Microbiology and Biotechnology, 2018,102(1):447-459.
[18] Mehrani Mohamad-Javad, Sobotka Dominika, Kowal Przemyslaw, et al. The occurrence and role of Nitrospira in nitrogen removal systems [J]. Bioresource Technology, 2020,303:122936.
[19] Zhao X D, Jiang J, Zhou Z, et al. Responses of microbial structures, functions and metabolic pathways for nitrogen removal to different hydraulic retention times in anaerobic side-stream reactor coupled membrane bioreactors [J]. Bioresource Technology, 2021,329:124903.
[20] Chen Z, Chang Z Q, Zhang L, et al. Effects of carbon source addition on microbial community and water quality in recirculating aquaculture systems for Litopenaeus vannamei [J]. Fisheries Science, 2020,86(3): 507-517.
[21] Stultiens K, Kessel M, Frank J, et al. Diversity, enrichment, and genomic potential of anaerobic methane- and ammonium-oxidizing microorganisms from a brewery wastewater treatment plant [J]. Applied Microbiology and Biotechnology, 2020,104(16):7201-7212.
[22] Wang J, Liu X L, Jiang X B, et al. Facilitated bio-mineralization of N, N-dimethylformamide in anoxic denitrification system: Long-term performance and biological mechanism [J]. Water Research, 2020,186: 116306.
[23] Ren Y P, Lv Y, Wang Y, et al. Effect of heterotrophic anodic denitrification on anolyte pH control and bioelectricity generation enhancement of bufferless microbial fuel cells [J]. Chemosphere, 2020,257:127251.
[24] Li L, Dong Y H, Qian G S, et al. Performance and microbial community analysis of bio-electrocoagulation on simultaneous nitrification and denitrification in submerged membrane bioreactor at limited dissolved oxygen [J]. Bioresource Technology, 2018,258:168- 176.
[25] Pishgar Roya, Dominic John Albino, Sheng Z Y, et al. Denitrification performance and microbial versatility in response to different selection pressures [J]. Bioresource Technology, 2019,281:72-83.
[26] Yin X J, Rahaman Md Hasibur, Liu W B, et al. Comparison of nitrogen and VFA removal pathways in autotrophic and organotrophic anammox reactors [J]. Environmental Research, 2021,197:111065.
[27] Mustapha Nurul Asyifah, Sakai Kenji, Shirai Yoshihito, et al. Impact of different antibiotics on methane production using waste-activated sludge: mechanisms and microbial community dynamics [J]. Applied Microbiology and Biotechnology, 2016,100(21):9355-9364.
[28] Keren Ray, Lawrence Jennifer E, Zhuang Weiqin, et al. Increased replication of dissimilatory nitrate-reducing bacteria leads to decreased anammox bioreactor performance [J]. Microbiome, 2020, 8(1):7.
[29] 刘 洋,陈永娟,王晓燕,等.水库与河流沉积物中好氧甲烷氧化菌群落差异性研究 [J]. 中国环境科学, 2018,38(5):1844-1854.
Liu Y, Chen Y J, Wang X Y, et al. Microbial communities differences between aerobic methanotrophs in Miyun Reservoir and North Canal [J]. China Environmental Science, 2018,38(5):1844-1854.
[30] Zhang X J, Zhang H L, Chen Z, et al. Achieving biogas production and efficient pollutants removal from nitrogenous fertilizer wastewater using combined anaerobic digestion and autotrophic nitrogen removal process [J]. Bioresource Technology, 2021,339:125659.
[31] Pan Z L, Zhou J, Lin Z Y, et al. Effects of COD/TN ratio on nitrogen removal efficiency, microbial community for high saline wastewater treatment based on heterotrophic nitrification-aerobic denitrification process [J]. Bioresource Technology, 2020,301:122726.
[32] Pernicova Iva, Kucera Dan, Nebesarova Jana, et al. Production of polyhydroxyalkanoates on waste frying oil employing selected Halomonas strains [J]. Bioresource Technology, 2019,292:122028.
[33] Obulisamy Parthiba Karthikeyan, Mehariya Sanjeet. Polyhydroxyalkanoates from extremophiles: A review [J]. Bioresource Technology, 2021,325:124653.
[34] Zhang S S, Sun X B, Wang X M, et al. Bioaugmentation with Diaphorobacter polyhydroxybutyrativorans to enhance nitrate removal in a poly (3-hydroxybutyrate-co-3-hydroxyvalerate)-supported denitrification reactor [J]. Bioresource Technology, 2018,263:499-507.
[35] Chen Q, Li Q, Guo A L, et al. Transcriptome analysis of suspended aggregates formed by Listeria monocytogenes co-cultured with Ralstonia insidiosa [J]. Food Control, 2021,130:108237.
[36] Bhattacharjee A S, Sha W, Lawson C E, et al. Whole-Community Metagenomics in Two Different Anammox Configurations: Process Performance and Community Structure [J]. Environmental Science & Technology, 2017,51(8):4317-4327.
[37] An T T, Wang F, Ren L L, et al. Ratio of nitrate to ammonium mainly drives soil bacterial dynamics involved in nitrate reduction processes [J]. Applied Soil Ecology, 2022,169:104164.
[38] Zhuang J L, Zhou Y Y, Liu Y D, et al. Flocs are the main source of nitrous oxide in a high-rate anammox granular sludge reactor: insights from metagenomics and fed-batch experiments [J]. Water Research, 2020,186:116321.
[39] 张胜男,桑 楠. PM2.5暴露对小鼠肝脏能量代谢的影响 [J]. 环境科学学报, 2017,37(8):3207-3212.
Zhang S N, Sang N. Exposure to PM2.5promotes the conversion of energy metabolism in livers of mice [J]. Acta Scientiae Circumstantiae, 2017,37(8):3207-3212.
[40] 余苗苗,江 敏,吴 昊,等.亚硝酸盐和氨对日本沼虾肝胰腺代谢的影响 [J]. 生态毒理学报, 2019,14(3):99-111.
Yu M M, Jiang M, Wu H, et al. Effects of nitrite and ammonia on the hepatopancreas metabolism of[J]. Asian Journal of Ecotoxicology, 2019,14(3):99-111.
The structure and metabolic pathway of functional bacteria for nitrogen and carbon removal in Anammox.
CHANG Yao-feng1, GUO Meng-lei1, XIE Jun-xiang1, XIE Jia-wei1, CHEN Chong-jun1,2,3*
(1.School of Environmental Science and Engineering, Suzhou University of Science and Technology, Suzhou 215009, China;2.Jiangsu Key Laboratory of Environmental Science and Engineering, Suzhou 215009, China;3.Jiangsu Collaborative Innovation Center of Technology and Material of Water Treatment, Suzhou 215009, China)., 2022,42(3):1138~1145
In order to clarify the differences of the nitrogen and carbon removal functional bacteria community structure and metabolic pathway in the anammox granular sludge system with different concentrations of organic matter (50~150mg/L) and bamboo biochar, the microbial distribution and gene expression of carbon and nitrogen metabolism were studied by metagenomic sequencing in this paper. The results showed that the bamboo charcoal addition was significantly increased the relative abundance of anammox when the COD concentration was 50and 150mg/L, the relative abundance ofincreased from 2.0% and 1.8% to 2.9% and 4.5%. In addition, compared with the control group only with organic matter (without bamboo biochar), the bamboo biochar addition was significantly changed the microbial community structure. The abundance of DNRA (dissimilatory nitrate reduction to ammonium process) bacteria was decreased, while the abundance of denitrifying bacteria and carbon metabolism-related bacteria were increased, which indicated that bamboo biochar addition was helpful to maintain the stability of nitrogen metabolism pathway community structure of anammox, denitrification and DNRA. The network analysis of microorganisms was showed that the removal efficiency of TN was increased by the synergetic effect of different nitrogen removal bacteria groups. However, the resistance ofandcould be improved byenrichment with bamboo biochar addition. The KEGG annotation indicated that bamboo charcoal addition would be enhanced the carbon and nitrogen metabolism efficiency of anammox granular sludge system in the presence of organic matter, especially promoted the connection between EMP (glycolytic pathway) and TCA (tricarboxylic acid cycle).
metagenomics;anammox granular sludge;nitrogen and carbon removal;bacteria community structure;metabolism pathway
X703
A
1000-6923(2022)03-1138-08
常尧枫(1996-),男,江苏宜兴人,苏州科技大学硕士研究生,研究方向为环境污染控制理论与技术.发表论文1篇.
2021-08-17
江苏省自然科学基金资助项目(BK20201450);中国博士后科学基金资助项目(2020M671400);苏州市民生科技项目(SS202016, SS2019022);江苏水处理技术与材料协同创新中心预研项目(XTCXSZ2019-3)
*责任作者, 副教授, chongjunchen@163.com
猜你喜欢
军事文摘(2022年16期)2022-08-24
环境工程技术学报(2022年3期)2022-06-05
科学(2020年6期)2020-02-06
第二课堂(课外活动版)(2018年5期)2018-08-20
科学与财富(2016年7期)2016-03-25
专用汽车(2015年2期)2015-03-01
绿色科技(2014年5期)2014-08-08
表面工程与再制造(2014年2期)2014-02-27
绿色科技(2011年3期)2011-04-19
