
微气候和适度放牧对高寒草甸植物群落特征的影响
2022-03-27 08:49斯确多吉尕藏加边巴普赤谢正新伪玛张次仁曲宗
草业科学 2022年12期
南 吉 ,斯确多吉,尕藏加,4,边巴普赤 ,谢正新,4,伪玛张,4,次仁曲宗, 4
(1.中国科学院大学, 北京 100049;2.中国科学院青藏高原研究所青藏高原地球系统与资源环境全国重点实验室, 北京 100101;3.西藏那曲高寒草地生态系统国家野外科学观测研究站,西藏 那曲 852000;4.西藏大学理学院生物系, 西藏 拉萨 850000)
放牧是草地生态系统主要的土地利用方式之一,但放牧对草地植物种群、群落及其结构和功能的影响因草地生态系统类型(荒漠、灌丛、草原、草甸等)及其所处的自然环境条件(高海拔山区、平原、温带、寒带等)和放牧利用方式(不同放牧强度)呈现差异[1-3]。据报道,在南美洲巴塔哥尼亚潘帕斯草原上适度放牧有效提高了生产力,但对物种组成的影响不显著[4];欧洲西南部和我国内蒙贝尔加地区的斯泰普草原上适度放牧有利于植物多样性的提高[5-6],但青藏高原东缘甘肃天祝草原区域适度放牧显著降低了植物多样性[7]。适度放牧对青藏高原腹地各类草地生态系统影响的研究相对匮乏,尤其利用野外控制试验探讨其变化过程和机理的研究更十分鲜见[8-10]。
青藏高原腹地是高寒草地生态系统典型发育区,是我国草地主要分布区域之一,是我国重要的生态安全屏障,也是西藏草地畜牧业发展的重要基础[11]。近年来,随着青藏高原各省区社会经济状况不断发展,草地利用方式也在发生着深刻变化[12-13]。在青藏高原大部分牧区因家畜数量加,草地放牧压力加大,草地生态系统呈现退化趋势的同时,部分区域则因经济收入来源变化(例如冬虫夏草经营、外出务工等)等原因,出现弃牧现象[14-15]。与此同时,国家为了缓解草地生态系统退化压力,恢复草地生态系统结构和功能,实施了一系列禁牧、轮牧政策和措施[16]。但禁牧与放牧(例如适度放牧)对高寒草地生态系统的影响有何差异等科学问题有待深入研究,相关争议也在不断凸显[17-18]。另外,有大量研究表明,放牧对草地生态系统的影响受区域温湿度条件等环境因子的影响[19],但温湿度条件如何调控放牧对草地生态系统的影响?高寒草地植物种群和群落的结构和动态受草食动物采食(主要是放牧利用)还是气候环境主导(equilibrium and nonequilibrium dynamic)等科学问题至今没有明确定论。为探讨以上科学问题,本研究以青藏高原腹地纳木错区域作为研究对象,以高寒草甸植物群落特征的动态变化作为研究内容,利用长期野外控制试验研究探讨:适度放牧如何影响高寒草甸植物群落特征?适度放牧对高寒草甸植物群落的影响是否受土壤温湿度条件调控?
1 材料与方法
1.1 研究区域概况
本研究区域位于拉萨市当雄县纳木错附近,属于半干旱典型高寒草甸草地发育区,地理坐标30°41.208′~30°44.579′ N;91°01.117′~91°04.590′ E,平均海拔为4 500 m,植被类型是高寒草甸草地,高山嵩草(Kobresia pygmaea)为优势种,矮生嵩草(K.humilis)、钉柱委陵菜(Potentilla saundersiana)、二裂委陵菜(Potentilla bifurca)、青藏苔草(Carex moorcroftii)等为亚优势种,团垫黄芪(Astragalus arnoldii)、小景天(Sedum fischeri)、胀果棘豆(Oxytropis stracheyana)等为稀有种[20]。纳木错年平均气温约为-0.71 ℃,最高气温一般出现在6 月1 日至8 月30 日,这期间的平均温度约为8.43 ℃。最低温度一般出现在12 月1 日至2 月30 日,这期间的平均温度约为-9.59 ℃。年平均湿度约为52%,年降水量约为407 mm,主要降水集中在6 月到9 月[21](图1)。
图1 青藏高原纳木错野外观测区域定位图Figure 1 Map of the observational area of the Namtso field of the Qinghai-Tibetan plateau
1.2 研究设计
2008 年,在纳木错附近典型高寒草甸草地选择约2 hm2草地进行了围封,于2009 年起通过严格控制放牧家畜数量与放牧时间开展牦牛放牧试验。本研究涉及2 个处理,放牧(Grazing)和不放牧(对照,Control),每个处理设8 个重复,每个处理小区是直径为8 m 的圆形区,共计16 个处理小区。自2009年起,每年夏季分3 次(6 月、7 月、8 月中旬)在牦牛放牧试验小区中央用铁钉固定1 条长度约4 m 的绳子拴住1 头成年牦牛,任其在栓绳范围内连续3 d,每天约7 h 进行自由采食,同时不定期(按天气条件决定)喂水。试验初期,对放牧试验小区内不同放牧时长条件下牦牛的采食强度进行了测定。当牦牛采食后剩余的生物量、植被高度等指标均与适度放牧条件下的天然健康草甸植被特征基本一致时确定为适度放牧强度[22]。此外,在每个试验小区内,随机选择5 个直径为1.5 m 的小样圆,固定观测植物群落特征、微气候环境监测、土壤样品采集等。植物群落特征和微气候(地表10 cm 空气温度和地下10 cm 土壤温湿度)监测小样圆内除放牧处理之外未被人为干扰。放牧处理期间,放牧处理样方内的地表空气温度监测探头要移开,以免家畜踩踏破坏监测设备。
1.3 数据采集
1.3.1 微 气候数据
利 用Decagon ECT、5 TM 和EC-TM 传 感 器 和EM50 记录器(Decagon Devices,Pullman,Washington,USA)测量处理样圆内距离地面10 cm 处的植被表层空气温度(air temperature)和地下10 cm 处的土壤温度(soil temperature)和土壤湿度(soil moisture),所有传感器每15 min 记录一次测量结果,并利用每年5 月25 日 至8 月25 日(2010、2011、2012、2014、2015、2016、2018、2019、2020 年)的数据来统计每个小区每年的生长季平均微气候特征,同时排除由于传感器故障引起的异常值。如果一个小区缺失连续8 d 以上的微气候数据,将排除在统计计算之外,以防止结果出现误差。
另外,借鉴关于年份划分标准的相关研究[23],每年的生长季节(5 月25 日-8 月25 日)的土壤湿度作为年份类型划分的依据,利用2010-2020 年5 月25 日至8 月25 日的土壤湿度平均值,其相对变异值可以采用如下公式来计算。

若相对变异值RVYear∈(-∞,-0.3],则该年份植物生长而言定义为干旱年;若相对变异值RVYear∈(-0.3,0.3),则该年份植物生长而言定义为正常年;若相对变异值RVYear∈[0.3,+∞),则该年份植物生长而言定义为湿润年。
根据上述土壤湿度相对变异计算公式以及年份类型划分标准,可以得到2010-2020 年试验期间每年对应的年份类型(表1)。

表1 基于5 月-8 月土壤湿度的各年份气候类型Table 1 Annual climate conditions based on soil moisture variations from May to August
1.3.2 植物群落特征与多样性
在每年8 月中旬,当大多数植物达到最大生长时,利用样方(0.75 m × 0.75 m)测量植物群落特征。为了确保每年在同一地点进行测量,将样方放置于小样圆中心,其四角用金属桩进行固定,样方框用细线均匀分成400 个栅格单元进行测量。观察400 个栅格中各类植物物种出现与否,并分别用“1”和“0”记录各自的频率。由于劳动力短缺,2014 年本试验只记录了100 个栅格单元内的物种频 率 数 据(0.75 m × 0.75 m 样 方 框 平 均 划 分 为100 个栅格单元)。这种方法比点截距法提供更高的观测精度[24]。由于观测人员短缺,2009、2013 和2017 年未获取到相关数据。本试验利用每年观测获取的植物分种频度数据计算各样方内的多样性指数、物种丰富度、优势度和均一度。多样性指数(diversity index)采用辛普森多样性指数(Simpson diversity index),为测度各样方多样程度,即:群落中连续两次抽样所得到的个体数属于同一种的概率。丰富度(richness)为样方内出现的所有物种数总和。优势度(dominance)反映了群落中各物种种群数量的相对变化状况,即:各物种间种群数量差异较大时优势度较高,相反则优势度较低。均匀度(evenness)与优势度相反,即:群落中各物种种群数量差异较小时均一度较高,相反则群落均一度较低。上述植物群落多样性指标均使用PAST 软件来获得[25]。植物不同功能类群盖度为相对盖度,即:每种不同功能类群的个体数占群落中总个体数的比例。本研究中可将植物功能类群分化为:杂类草、莎草科、灌木类、禾本科。
1.3.3 数 据分析
本研究利用SPSS 21.0 软件中的一般线性模型多变量方差分析方法来分析各年份、各处理及其交互作用对土壤温湿度、植物多样性、物种丰富度、优势度、均一度、不同功能类群相对盖度的影响;利用SPSS 21.0 软件中的线性回归进行分析方法来分析不同放牧处理条件下微气候变化特征与植物多样性、物种丰富度、优势度、均一度、不同功能类群相对盖度等变量之间的相关关系。绘图工具使用Graphpad prism 8.0.1。
2 结果与分析
2.1 适度放牧对植被表层空气温度和土壤温、湿度的影响
与对照比较,适度放牧显著降低了土壤湿度(P<0.001),降幅为7.86%,但空气温度和土壤温度差异不显著(P> 0.05) (图2)。适度放牧对土壤湿度、土壤温度和空气温度的影响在年份之间呈现显著差异(P< 0.001) (图3);2014、2016、2018、2019 年,适度放牧显著降低了土壤湿度(P< 0.05),其余年份(2010、2011、2012、2015、2020 年)没有显著差异(P>0.05);2010 和2012 年,土壤湿度较其他年份低,分别为11.20%和12.45%,2018 年和2020 年较其他年份高,分别为26.48%和28.89%。2010 和2012 年空气温度较其他年份高,分别为8.47 和8.44 ℃,2011年最低(7.01 ℃)。2012 年土壤温度最高(11.87 ℃),2019 年最低(9.73 ℃)。

图2 适度放牧对空气温度、土壤温度、土壤湿度的影响Figure 2 Effects of moderate grazing on the air and soil temperatures and soil moisture

图3 适度放牧对不同年份空气温度、土壤温度、土壤湿度的影响Figure 3 Effects of moderate grazing on the air and soil temperatures and soil moistures across 10 years
2.2 适度放牧对植物群落多样性的影响
与对照比较,适度放牧显著降低了辛普森多样性指数(降幅为8.8%) (P< 0.001),显著提高了优势度(幅度为15.13%) (P< 0.001),但对物种丰富度、均匀度没有显著影响(P> 0.05) (图4)。适度放牧对辛普森指数、物种丰富度、均匀度和优势度的影响在年份之间呈现显著差异(P< 0.001) (图5);试验前3 年,适度放牧显著降低了辛普森多样性指数,显著增加了优势度,后6 年则没有显著差异;无论放牧与否,试验前4 年辛普森多样性指数低于后5 年(平均值相差0.15),而试验前4 年优势度则高于后5 年(平均值相差0.14);另外,无论放牧与否,2010 和2012 年的物种均匀度较其他年份低,分别为0.23、0.24,2015 年最高(0.43);而2010 和2012 年物种丰富度较其他年份高,分别为15.50、16.25,2015 年较低(11.13) (图5)。

图4 适度放牧对植物群落优势度(DM)、辛普森多样性指数(DV)、均匀度(EV)、丰富度 (RC)的影响Figure 4 Effects of moderate grazing on plant communities evaluated using dominance (DM), Simpson diversity index(DV), evenness (EV), and richness (RC)
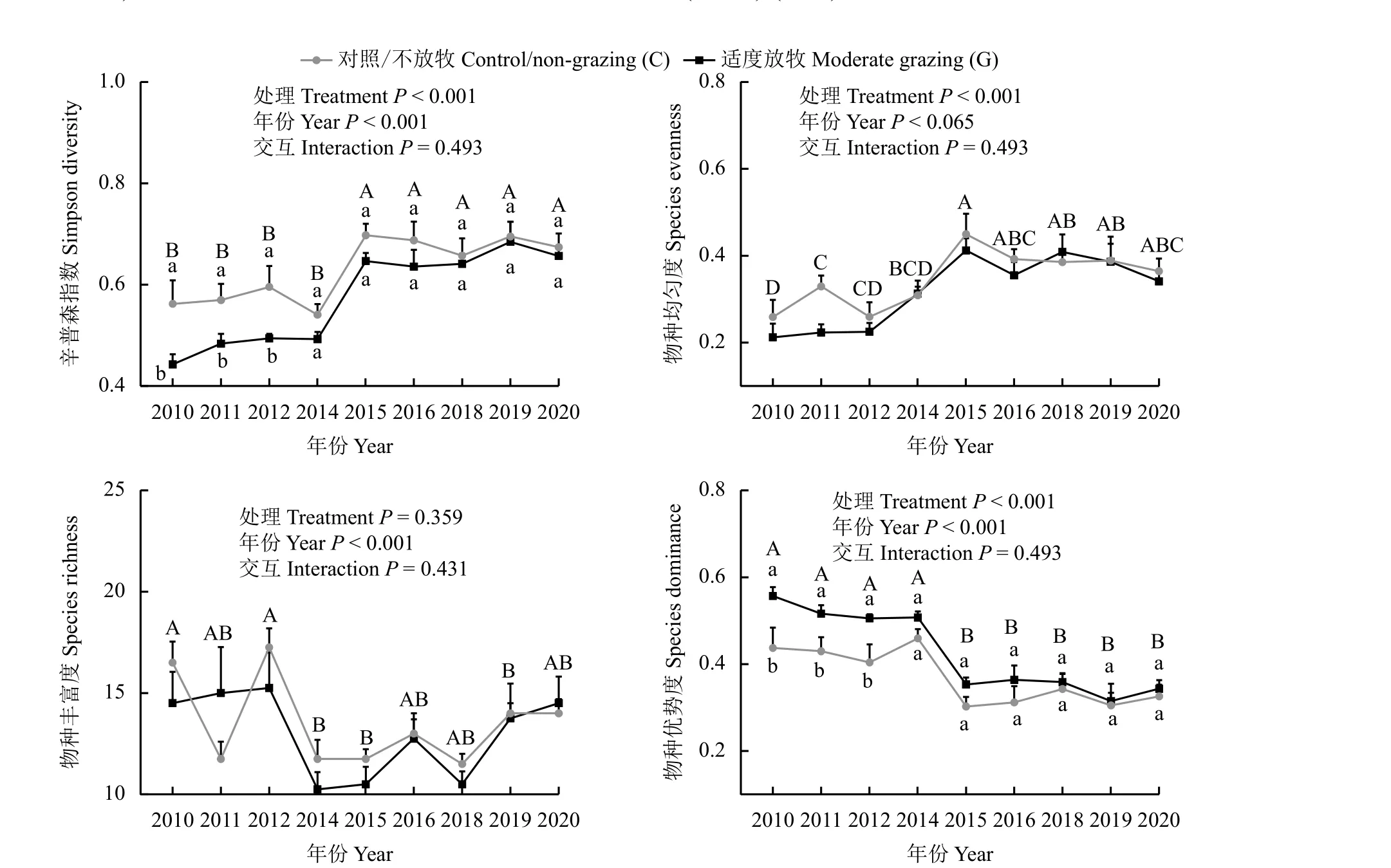
图5 适度放牧对不同年份植物群落多样性的影响Figure 5 Effects of moderate grazing on plant community diversity across 10 years
2.3 适度放牧对植物不同功能类群盖度的影响
与对照比较,适度放牧降低了禾本科类盖度(降幅为69%),提高了莎草科草类和灌木类的盖度(提高的幅度分别为7.8%、35%),但对杂类草盖度没有显著影响(P> 0.05) (图6)。杂类草、莎草科、禾本科的盖度在年份之间呈现显著差异(P< 0.05) (图7);2012 年,适度放牧增加了莎草科植物盖度;无论放牧与否,试验前4 年,莎草科植物盖度高于后5 年(平均值相差13.36%),试验前4 年杂类草植物盖度则低于后5 年(平均值相差14.62%);另外,与对照比较,适度放牧降低了2010、2011、2012、2015、2016年的禾本科植物盖度,其中适度放牧对2010 和2012年禾本科植物盖度的降幅高于其他年份(平均值相差3.48%) (图7)。
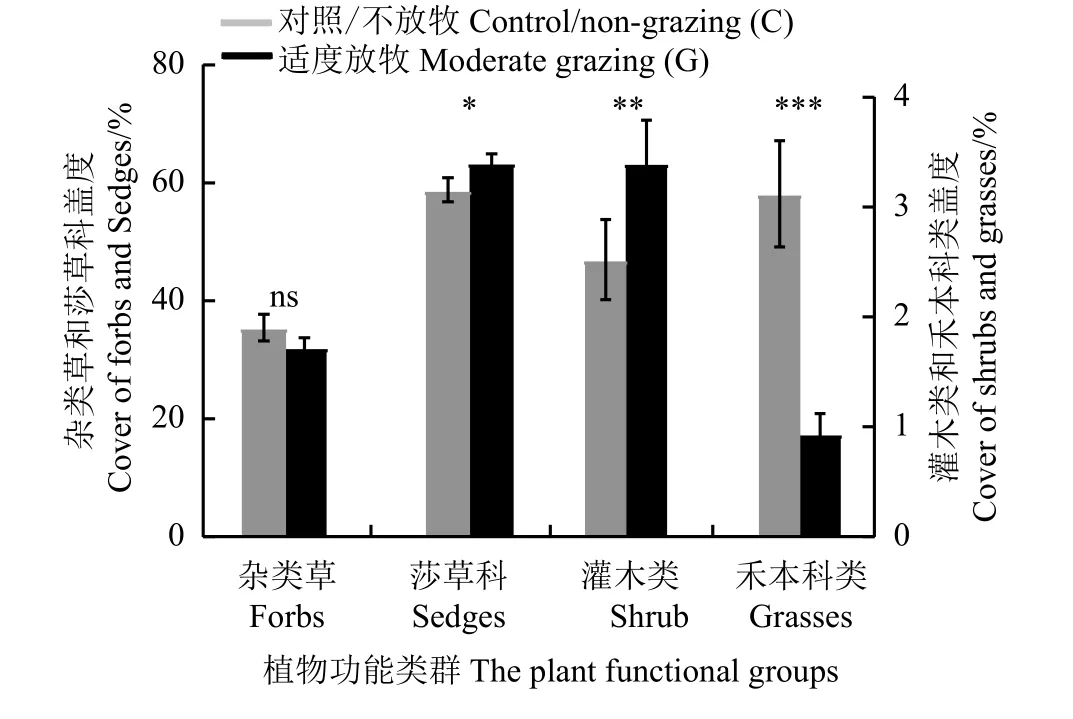
图6 适度放牧对植物不同功能类群相对盖度的影响Figure 6 Effects of moderate grazing on the relative coverages of different plant functional groups
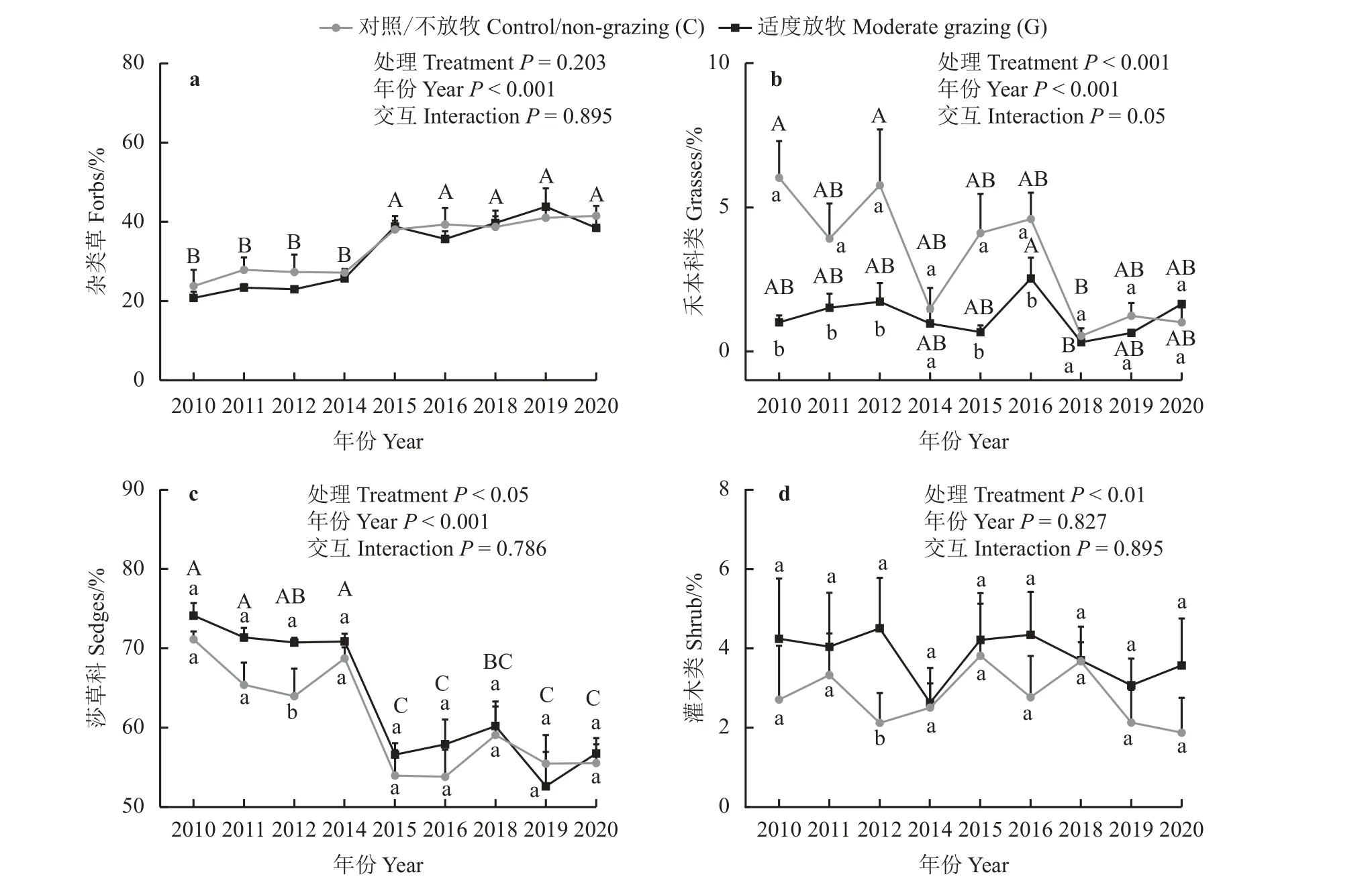
图7 适度放牧对不同年份植物不同功能类群盖度的影响Figure 7 Effects of moderate grazing on relative coverages of different plant functional groups across 10 years
2.4 不同气候类型放牧对植物群落特征的影响
2.4.1 不同气候类型适度放牧对植物群落多样性的影响
与对照相比,正常年份和干旱年份,适度放牧显著降低了辛普森多样性指数(P< 0.001),而在湿润年份没有显著影响(P> 0.05);辛普森多样性指数在湿润年份(0.66)显著高于正常年份(0.61)和干旱年份(0.52),正常年份和干旱年份之间没有显著差异;物种丰富度在干旱年份(15.87)高于正常年份(12.06)和湿润年份(12.63);均匀度在干旱年份(0.24)低于正常年份(0.35)和湿润年份(0.37),而正常年份和湿润年份之间没有显著差异;优势度在干旱年份(0.48)高于正常年份(0.39)和湿润年份(0.34),与对照相比,干旱年份,放牧增加了优势度,而在正常年份和湿润年份放牧对优势度没有显著影响(图8)。
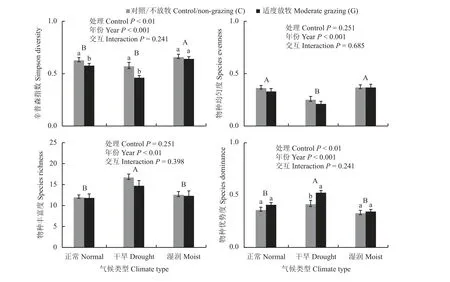
图8 不同气候类型下适度放牧对植物群落多样性的影响Figure 8 Effects of moderate grazing on plant community diversity under different climate conditions
2.4.2 不同气候类型下适度放牧对植物不同功能类群的盖度的影响
杂类草盖度在干旱年份(23.76%)低于正常年份(32.57%)和湿润年份(43.20%);禾本科盖度在干旱年份(5.89%)高于正常年份(3.44%)和湿润年份(0.69%),与对照相比,正常和干旱年份,适度放牧显著降低了禾本科盖度,但湿润年份无显著差异;莎草科盖度在干旱年份(69.08%)高于正常年份(62.28%)和湿润年份(54.46%);与对照相比,正常年份和干旱年份,适度放牧增加了莎草盖度,湿润年份适度放牧对莎草科盖度则没有显著影响;与对照相比,干旱年份,放牧增加了灌木类盖度,正常年份和湿润年份,适度放牧对灌木类盖度没有显著影响(图9)。

图9 不同气候类型下适度放牧对植物不同功能类群盖度的影响Figure 9 Effects of moderate grazing on relative cover of different plant functional groups under different climate conditions
2.5 土壤温湿度与植物多样性和不同功能类群盖度的关系
无论放牧与否,土壤湿度与辛普森多样性指数和物种均匀度呈显著正相关(P< 0.001),与物种优势度呈显著负相关(P< 0.001);土壤湿度与物种丰富度在适度放牧条件下相关性不显著,但在不放牧条件下呈显著负相关(P< 0.05) (图10)。

图10 适度放牧的土壤湿度与植物群落多样性的关系Figure 10 Relationship between soil moisture and plant community diversity under moderate grazing
无论放牧与否,土壤湿度和杂类草盖度均呈现正相关(P< 0.001),与莎草科盖度则呈负相关(P<0.001);在适度放牧条件下,土壤湿度与禾本科盖度相关性不显著;在不放牧条件下土壤湿度与禾本科盖度则呈显著负相关(P< 0.05);土壤湿度与灌木类盖度在不放牧条件下呈显著负相关(P< 0.05),但在适度放牧条件下相关性不显著(图11)。
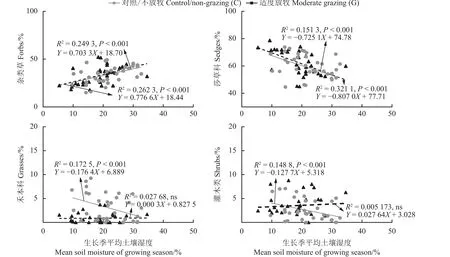
图11 适度放牧的土壤湿度与植物不同功能类群相对盖度的关系Figure 11 Relationship between soil moisture and relative coverage of different functional groups of plants under moderate grazing
3 讨论
3.1 适度放牧促进莎草类植物生长,长期禁牧则提高禾草类植物种群盖度
放牧家畜可通过采食、践踏和排泄等行为直接或间接影响植物及其群落结构特征[26],但影响程度随放牧强度和植被类型[27]以及自然环境特征[28]而不同。Georg 等[29]认为,高寒草甸植被类型是在长期适度放牧条件下形成的次生植被类型,因此当放牧干扰被禁止时高寒草甸草地可能会演替成为自然顶级群落。本试验利用11 年的围栏禁牧与适度放牧控制进行研究,结果表明,适度放牧显著降低了禾本科草类盖度,但提高了莎草类盖度(图6)。以高山嵩草为主的莎草类植物是高寒草甸草地的绝对优势植物,其在群落中的占比可高达80%以上;适度放牧提高高山莎草类盖度,表明适度放牧有利于形成或维持高寒草甸草地。事实上,以高山嵩草为主的大部分莎草类植物生长点较低、地下可通过庞大的根茎系统形成致密的草皮,十分耐牧[30],因此适度放牧可能通过采食地上生物量,诱发根茎生长,形成不定根,促进其无性繁殖能力,从而提高了高山嵩草在群落中的覆盖度。相反,高寒草甸常见的禾本科草类为针茅类、早熟禾类植物,大多丛生,相对高大,而且家畜十分喜食[3],因此在放牧条件下可被大量采食;但是,由于其在群落中的占比本身较低,不到2%,因此种群补偿能力低下,导致种群密度、盖度下降。值得一提的是,大量文献结果表明,放牧有利于抑制灌木生长[31-33],但本研究结果表明,放牧提高了灌木盖度(图6)。该研究区域的主要灌木为金露梅,尽管其叶片肉质,家畜可食,而且并非喜食物种。由于放牧试验期间(每年6 月、7 月、8 月中旬)草地植物茂盛,可食、甚至喜食植物较多,且放牧试验时间较短(每年3 次,每次在放牧试验样圆仅连续采食3 d),因此家畜可能并不需要采食灌木充饥,因此其开花、结果可能不受放牧影响。另外,本研究结果表明,放牧导致地表土壤湿度显著降低(P< 0.01) (图2),而灌木类植物因为根系较深,可以有效吸收较深的土壤水分[34],因此在放牧条件下,不受采食压力时,生长和繁育能力可能未受影响,受这些因素综合影响,可能导致其种群得到扩张。
3.2 土壤湿度调控放牧对高寒草甸植物群落变化特征
本研究结果表明,适度放牧对高寒植物群落特征的影响在不同年份差异明显(图5、图7),可能是不同年份之间的水热条件差异所致。尽管温度被认为是引起高寒高海拔地区植物种群和群落变化的重要环境因素[35],但本研究结果表明,放牧与不放牧处理小区之间的平均土壤和地表空气温度均差异不显著,放牧显著降低了土壤湿度(图2);而在干旱年份,无论放牧与否,杂类草盖度显著低于正常和湿润年份,不放牧时禾草类盖度则显著高于正常和湿润年份(图9)。这与植物功能类群的形态和生理特征相关[36]。杂类草植物均为双子叶宽叶直根系植物,单位质量的叶面积较大,可以通过气孔来调节蒸腾作用,但是极端的干旱可能会破坏植物的光合作用能力,导致植物叶片损伤而水分降低[37]。禾本草类为单子叶细叶、须根系植物,叶面蒸腾丧失水分相对较少,在干旱年份具备一定的优势。另外,本研究发现,除了湿润年份,放牧在干旱年份和正常年份显著降低了多样性(辛普森指数),增加莎草科植物盖度(图8、图9)。放牧导致植物地上生物量被采食,降低其生长能力、种子繁殖力下降[38]。除莎草科植物之外,其他植物,尤其是双子叶植物,主要通过种子繁殖扩大其种群密度[39]。当放牧且降水量补给不足时(例如干旱年份),这些植物的生长和繁殖能力更加不足,可能导致部分植物种群密度下降,尤其是稀有植物和种群密度较低的植物物种丧失,导致多样性降低。
4 结论与展望
综上,可得出以下结论:1)适度放牧有利于维持和促进高山嵩草为优势的高寒草甸草地植被类型。2)高寒草甸长期禁牧(不放牧)可提高禾草类植物种群,可能导致草甸转变为以禾草类占优势的高寒草原化草甸类草地。3)土壤湿度可调控放牧对高寒草甸植物群落变化特征。
展望:由于目前在本研究中报道的数据分析结果仅为适度放牧对植物群落特征的多年平均水平影响,而并未深度探讨高寒草甸植物群落对适度放牧效应的敏感性以及稳定性,未来有待进一步深入分析相关内容。另外,放牧对植物群落的影响在某种程度上受局域气候条件的影响,本研究在纳木错开展的试验还包括候变化因素(不同季节增温、早春增雪)设计,未来有待深入分析气候变化因素与禁牧和放牧因素的交互影响,更深层次揭示气候变化和放牧/禁牧对植物群落特征的交互影响。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
黑龙江工程学院学报(2020年5期)2020-10-21
天津农林科技(2020年3期)2020-08-13
水土保持研究(2019年6期)2019-10-19
电子制作(2019年15期)2019-08-27
活力(2019年21期)2019-04-01
河北林业科技(2016年5期)2016-11-08
高原山地气象研究(2016年4期)2016-02-28
高原山地气象研究(2016年4期)2016-02-28
