
卡森斯氏和短尾异小杆单菌线虫的制备方法
2022-03-26 02:15李茜童张文德杨亚贤钱秀娟
草业科学 2022年3期
李茜童,张文德,杨亚贤,钱秀娟
(甘肃农业大学植物保护学院 / 甘肃省农作物病虫害生物防治工程实验室, 甘肃 兰州 730070)
线虫是一类种类庞大的物种,在自然界中仅次于昆虫。线虫与昆虫之间存在普遍而广泛的的联系,其中少数线虫因对昆虫具有较强的侵染致死力被称为昆虫病原线虫 (entomopathogenic nematodes,EPNs),通常与其携带的共生细菌一同引起寄主昆虫死亡[1],是一类带有共生菌的专性寄生线虫[2]。
共生细菌存在于侵染期线虫肠道内,该线虫进入寄主昆虫血腔并在其中繁殖时,寄主昆虫体内营养物质被分解,共生菌被释放,大量的共生菌繁殖产生毒素使寄主昆虫败血死亡。共生菌的大量繁殖在这一过程中起到两个作用,一是产生抑菌物质(如抗生素)以抑制其他微生物的生长,为线虫提供适宜的生长环境;二是降解其他物质为线虫提供营养[3-4]。线虫与细菌共生关系的研究主要集中在携带共生菌的斯氏线虫属(Steinernema)与异小杆线虫属(Heterorhabditis)等昆虫病原线虫上[2,5],其具有寄主范围广泛,致死速度快,寄生杀虫效果可观,对脊椎动物、植物、人类无害,且对环境安全、无有毒残留物,植物不易产生抗药性等特点[6]。此外,线虫在土壤中的生存能力和扩散能力强,对害虫(尤其是对土栖性和钻蛀性害虫有替代化学药剂的潜能)具有主动搜寻能力[7-10],其侵染期幼虫[11]能存活1 到多个月,可进行产业化培养[1,12]生产和销售,并广泛应用于害虫的生物防治领域[13]。
近年来,由于草原退化导致的草地病、虫、鼠害愈发猖獗,进而引起生态恶化、牧草产量和品质降低,以及生物多样性减少等问题[14]。化学农药治理见效快却有很大毒性,且残留量高,虽然根据我国目前所提倡“预防为主,综合治理”的方针,已将昆虫病原线虫应用于防治多种农林、花卉以及卫生害虫,并且取得了令人鼓舞的成果[1],但在草地害虫防治方面仍有巨大进步空间。单菌病原线虫在产业和科学研究中具有重要作用。研究表明,细菌和线虫的共生关系是具有一定程度专化性的,即一种线虫只能同一种细菌共生。因此,单菌线虫的培养获得尤为重要。研究制备只携带专化性共生菌的昆虫病原线虫的方法不仅能够明确昆虫病原线虫与其共生菌的关系,也有利于侵染期幼虫的产业化生产制备,并提高昆虫病原线虫对草地害虫的防治效果[15]。
本研究以从甘肃省分离诱集得到的抗低湿胁迫能力较强的卡森斯氏线虫(Steinernema kraussei)和非耐旱性短尾异小杆线(Heterorhabditis brevicaudis)两种昆虫病原线虫为研究材料,分离其携带的共生菌,并采用直接从大蜡螟(Galleria mellonella)体内分离种子怀卵成虫至纯化好的专化性共生菌上孵育单菌线虫的简易方法,以期快速准确获得高防效的侵染期幼虫,为昆虫病原异小杆属和斯氏属线虫的产业化生产奠定基础,为高效防除害虫提供理论依据。
1 材料与方法
1.1 供试昆虫
大蜡螟3 龄幼虫由甘肃农业大学植物保护/甘肃省农作物病虫害生物防治工程实验室保存。
1.2 供试线虫
抗低湿胁迫能力较强的卡森斯氏线虫0657L 和非耐旱性短尾异小杆线虫0641TY 均由甘肃农业大学植物保护学院昆虫生态实验室/甘肃省农作物病虫害生物防治工程实验室保存。
1.3 培养基的配制
NBTB 培养基:琼脂15 g,LB 琼脂预混干粉40 g,溴百里酚蓝0.01 g,无菌水1 000 mL,摇匀。配置好培养液,于121 ℃,1 240 Pa 灭菌20 min,倒平板培养基保存备用。
LB 培养基:营养肉汤8 g,琼脂15 g,酵母粉5 g,六水合氯化镁(MgCl2·H2O) 12 g,蒸馏水900 mL,摇匀。配置好培养液,于121 ℃,1 240 Pa 灭菌20 min。玉米糖浆7 mL,玉米胚芽油4 mL,蒸馏水89 mL,混合均匀,加入已灭菌的培养液中,置于平板培养基保存备用。
1.4 共生菌的分离
在两个直径为9 cm 的培养皿内铺垫两层无菌滤纸,将经过75%酒精进行体表消毒的3 龄大蜡螟(大小、生理活性基本一致)各5 头置于其中,用移液枪分别吸取1.5 mL 密度为3.84 头·μL-1的卡森斯氏线虫和密度为2.82 头·μL-1的短尾异小杆线虫悬浮液滴至大蜡螟头部附近体壁上,在20~25 ℃培养箱中放置24 h。
在超净工作台上,用75%酒精消毒的小剪刀将经过线虫侵染的大蜡螟第1 对胸足剪破,用拇指和食指从虫体尾部向前按压,使大蜡螟的体液从第1 对胸足伤口处流出,滴加在NBTB 平板培养基上。接菌环在酒精灯下灭菌,蘸取大蜡螟体液,划Z 字型分离共生菌。用封口膜密封培养基,置于20 ~25 ℃培养箱中培养。侵染24、28、32、36、40 h 各分离一次,每次分离3 皿,培养后比较菌落生长情况。
1.5 共生菌的纯化
卡森斯氏线虫和短尾异小杆线虫共生菌分别培养至出现易于挑取的单菌落,无菌条件下每8 h 挑取绿色单菌落一次,转移至新的NBTB 平板培养基上,继续划线进行共生菌的二次纯化,以此重复,直到培养纯化出均匀的单菌落。挑取纯化的初生型单菌落转移至LB 平板培养基,在20 ~ 25 ℃培养箱中培养72 h 后保存备用。
1.6 单菌线虫的制备
在两个直径为9 cm 的培养皿内铺垫两层无菌滤纸,将经过75%酒精消毒的3 龄大蜡螟各5 头置于其中,用移液枪分别吸取1.5 mL 密度为3.84 头·μL-1的卡森斯氏线虫和密度为2.82 头·μL-1的短尾异小杆线虫悬浮液侵染大蜡螟。侵染7 d 后,在显微镜下用酒精灯灭过菌的昆虫针解剖侵染至死的大蜡螟,观察并分别挑取大蜡螟体内的卡森斯氏线虫和短尾异小杆线虫怀卵成虫各1 头,接到长好共生菌的LB 平板培养基上,用封口膜密封培养基,并将培养基置于培养箱中继续培养,每24 h 观察一次幼虫发育情况,连续观察2 周。
1.7 数据统计
观察侵染不同时间的大蜡螟体内初次分离的共生菌在NBTB 上培养出单菌落的情况。
记录怀卵成虫在培养有共生菌的LB 培养基上的存活情况,获得侵染期幼虫(即3 龄幼虫)所需最短时间和全部幼虫形成侵染期幼虫的时间。
2 结果与分析
2.1 共生菌的分离及纯化
观察比较两个品系线虫悬浮液接种大蜡螟24、28、32、36 以及40 h 后,从其体内分离出来的共生菌的生长情况。
2.1.1 卡森斯氏线虫专化性共生菌
卡森斯氏线虫侵染大蜡螟24 h 后分离获得的共生菌更易在NBTB 培养基上培养出较为均匀生长的单菌落(图1),便于被二次纯化。
在NBTB 培养基上,经过纯化的卡森斯氏线虫共生菌呈墨绿色,菌株可以吸收培养基中的溴百里酚蓝,周围培养基也呈现出蓝色,菌株大,边缘整齐规则,表面凸起有光泽(图2);在LB 培养基上,菌株呈现棕黄色。经在黑暗条件下观察有生物荧光,为发光杆菌属。

图2 卡森斯氏线虫共生菌在NBTB 培养基上的菌株形态Figure 2 Morph of the symbiotic bacteria of Steinernema kraussei on NBTB
2.1.2 短尾异小杆线虫专化性共生菌
从短尾异小杆线虫侵染大蜡螟28 h 分离获得的共生菌在NBTB 培养基上生长最佳(图3),有较均匀单菌落,便于二次纯化。

图3 短尾异小杆线虫侵染大蜡螟28 h 分离的共生菌Figure 3 Symbiotic bacteria isolated from Galleria mellonella infected by Heterorhabditis brevicaudis for 28 hours
在NBTB 培养基上,经过纯化的短尾异小杆线虫共生菌也呈墨绿色,菌株吸收培养基中的溴百里酚蓝,但周围的培养基呈明显不规则黄色,菌株较小,边缘稍不规则,表面较为扁平,中心有小突起(图4);在LB 培养基上,菌株也呈现棕黄色,经在黑暗条件下观察无生物荧光,为嗜线虫杆菌属。共生细菌种的鉴定,拟另文发表。

图4 短尾异小杆线虫共生菌在NBTB 培养基上的菌株形态Figure 4 Morph of the symbiotic bacteria of Heterorhabditis brevicaudis on NBTB
2.2 种子怀卵成虫的培养及在LB 培养基上的培养
用线虫悬浮液侵染大蜡螟24 h 后,大蜡螟患败血症死亡,开始膨胀,褪色。侵染1 周后,在解剖镜下解剖大蜡螟,两个品系线虫悬浮液侵染的大蜡螟体内均存在多数幼虫及少数怀卵成虫。分别将单头怀卵成虫挑取接种至 LB 培养基上恒温培养观察。
2.2.1 卡森斯氏线虫
斯氏属的种子怀卵成虫繁殖较慢,在LB 培养基存活时间约为3 d,接入前两天产出第二代,虽在解剖镜下可明显看到怀卵成虫体内的卵粒(图5),也可看到卵在母体内孵化成幼虫(图6),但并未观察到体外有卵粒排出,第二代均以幼虫形态钻出怀卵成虫体外。怀卵成虫最短仅存活2 d,最长存活5 d 后死亡。

图5 卡森斯氏线虫怀卵成虫体内的卵粒Figure 5 Eggs in adult Steinernema kraussei

图6 卡森斯氏线虫怀卵成虫体内孵化的幼虫Figure 6 Hatching juveniles in a gravid Steinernema kraussei female adult
2.2.2 短尾异小杆线虫
异小杆属的种子怀卵成虫自接入LB 培养基上的存活时间约为3 d,接入后2~3 d 即产出下一代,繁殖速度相对较快,但下一代同样是以幼虫形态钻出母体(图7)。怀卵成虫最短存活3 d,最长5 d 即死亡。

图7 短尾异小杆线虫幼虫钻出已死亡的母体Figure 7 Juveniles of Heterorhabditis brevicaudis emerging from the body of the dead female adult
2.3 侵染期幼虫的形成
2.3.1 卡森斯氏线虫
卡森斯氏线虫侵染期幼虫在LB 培养基中形成较快,最快需要2 d 时间即可观察到侵染期幼虫的形成。在观察的2 周内,可观察到其2 龄幼虫均可全部转化成侵染期幼虫,而且龄期整齐,全部转化为侵染期幼虫所需时间约为10 d (图8)。

图8 卡森斯氏线虫侵染期幼虫在LB 培养基上的形成过程Figure 8 Steinernema kraussei infective juveniles cultured on LB medium
2.3.2 异小杆线虫
异小杆线虫侵染期幼虫在LB 培养基中较慢形成,最长需要4 d 才可观察到侵染期幼虫的形成。在观察的2 周内,可观察到卡森斯氏线虫2 龄幼虫均可全部转化成龄期整齐的侵染期幼虫,全部转化为侵染期幼虫所需时间为15 d (图9)。
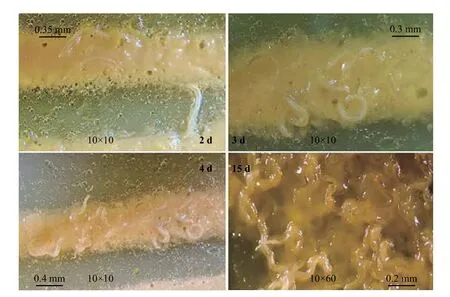
图9 异小杆线虫侵染期幼虫在LB 培养基上的形成过程Figure 9 Heterorhabditis brevicaudis infective juveniles cultured on LB medium
3 讨论与结论
本研究选用了卡森斯氏线虫和短尾异小杆线两个品系的线虫侵染大蜡螟,分别于接种24、28、32、36 以及40 h 大蜡螟体内分离其共生菌,并纯化成具有专化性的单菌,观察菌株生长情况,以及单菌下侵染期幼虫诱导形成的情况。结果表明,卡森斯氏线虫共生菌分离的最佳时间为接种24 h,而短尾异小杆线虫共生菌分离的最佳时间为接种28 h。结合分析培养过程中菌株的形态特征和分离、纯化、培养所需时间的差异,有力地证明了线虫与细菌的一一对应关系,即不同品系的线虫其肠道内共生菌的种类也是不同的。加之单菌培养制备线虫通气性好、产量高、生产效率高[15],因而可以推断这种专化性能够较好的保证昆虫病原斯氏属和异小杆属线虫的生活力、生殖力和致病力。
此外,斯氏线虫属和异小杆线虫属共生细菌均具有两型现象,通称共生细菌菌型的初生型与次生菌。在线虫大量培养中,培养基必须接入初生型共生菌。初生型能把培养基中的一系列物质转化为适宜于线虫生长繁殖的营养基质,而次生型则不能[16],因此,线虫只吸收并保留初生型共生菌[17]。
虽然在LB 培养基中共生细菌不能大量生长[1],但接入异小杆属和斯氏属两个不同品系的怀卵成虫均能存活,将怀卵成虫接入LB 培养基后,均能在较短时间内产出下一代幼虫。同时,可以清楚地观察到怀卵成虫体内的卵,有些也可以观察到卵在母体内孵化成幼虫,但并未观察到卵被排出体外,而是以幼虫钻出母体,即卵可以在母体内孵化,这种现象叫噬母现象[3],是指卵在种子怀卵成虫体内孵化,幼虫靠母体的营养物质存活,当母体营养物质被消耗完后钻出母体。这些幼虫大多为2 龄幼虫,或者2 龄幼虫至侵染期幼虫的过渡虫态,2 龄幼虫及过渡虫态在LB 培养基上存活并发育形成侵染期幼虫。由于LB 培养基上的菌落接种采用了Z 字形划线法,线虫会顺着菌落层层扩散分布,使得培养获得的侵染期幼虫龄期整齐。
综上所述,本研究中单菌线虫的制备利用了昆虫病原线虫与细菌共生关系的专化性,通过使用两个不同品系线虫的怀卵成虫,筛选到一种直接从大蜡螟体内分离怀卵成虫至纯化好的共生菌上孵育线虫,以快速准确获得整齐龄期的侵染期幼虫的简便方法,弥补了颜珣和韩日畴[1]研究的以不同的孵育液孵育小卷蛾斯氏线虫(S.carpocapsae)的怀卵成虫,找到可以简单快速从怀卵成虫直接获得整齐龄期的侵染期幼虫的方法中营养成分介于纯水和Ringer’s液中的生理盐水及PBS 中,无法诱导获得全部的侵染期幼虫的不足;明确了昆虫病原线虫与其共生菌的一一对应关系;为这两种品系线虫侵染期致病力的研究以及在草地害虫安全防治中的商业化生产提供技术支持;为产品的低成本生产、贮存技术、质量控制程序[18-19],为更多植物病虫害寻找新的防治途径[20]打下基础。为证实单菌线虫制备方法的意义,本研究也将在后续进行单菌线虫致病力测定。
猜你喜欢
今日农业(2022年14期)2022-09-15
新农村(浙江)(2022年7期)2022-07-07
当代水产(2022年1期)2022-04-26
中国生物防治学报(2022年1期)2022-04-22
中国瓜菜(2019年8期)2019-09-19
现代农业科技(2017年13期)2017-08-09
农业科技与装备(2017年1期)2017-06-09
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
现代园艺(2016年7期)2017-01-09
