
禁牧年限对温性荒漠草地群落特征及土壤因子的影响
2022-03-26 02:15王勇辉徐海量赵万羽刘星宏金额斯别克木哈德勒哈勒哈什唆坦哈机库丽沙依拉吾那提汗
草业科学 2022年3期
郎 鹏,王勇辉,徐海量,赵万羽,刘星宏,金额斯别克·木哈德勒,哈勒哈什·唆坦哈机,库丽沙依拉·吾那提汗
(1.新疆师范大学地理科学与旅游学院, 新疆 乌鲁木齐 830054;2.中国科学院新疆生态与地理研究所, 新疆 乌鲁木齐 830011;3.新疆干旱区湖泊环境与资源实验室, 新疆 乌鲁木齐 830054;4.阿勒泰地区富蕴县林业与草原局, 新疆 富蕴 836100)
新疆丰富的草地资源,是发展畜牧业的重要物质基础,同时对维持国家生态安全具有重要作用[1]。作为新疆畜牧业主要生产基地之一[2],阿勒泰地区由于过度放牧和气候变化等因素的影响,近年来草地生态系统平衡遭受破坏,草地生产力急剧下降,可食优良牧草比例降低,进而致使土壤退化,最终导致部分地区草地退化[3]。为此,当地政府采用围栏禁牧等措施实施治理,通过降低或阻断家畜对草地群落生态系统的影响,使其得以自然恢复[4]。同时,围栏封育下草地的物种多样性[5-6]、植被碳储量[7]、土壤理化性质[8]、土壤种子库[9]和草地生产力[10-11]、总生物量[12-13]等均出现明显变化。因此,围栏封育对退化草地的恢复具有重要的研究意义。上述研究表明围栏封育的效果是随着封育时间的变化而发生变化的,随着围栏措施的广泛应用,一些学者发现:围栏封育可以提高土壤有机质、碱解氮和速效钾的含量,改善地上植被状况[14-15],但较长时间的草地围封不利于优化草地群落结构,同时也会限制草地生态系统稳定性的维持[16-17]。
土壤作为草地群落植物主要的生命活动场所,对草地群落植被的生长发育起着极其重要的作用[18]。土壤的养分含量对草地群落植被的恢复和生长有直接影响。因此,监测不同围栏封育年限草地群落土壤养分指标的变化对于探讨适宜的封育措施具有重要意义[19]。基于此,本研究通过调查不同禁牧年限对草地群落高度、密度、盖度和生物量等的影响,分析不同禁牧年限对草地群落的生长特征,并通过测定不同禁牧年限草地的土壤全氮、全磷、全钾、土壤含水率、pH 和电导率,了解不同禁牧年限草地群落与土壤理化性质的关系,探讨禁牧适宜时间,以期为新疆阿勒泰地区温性荒漠草地的保护、恢复和利用提供参考依据。
1 研究区概况与研究方法
研究地点位于新疆维吾尔自治区阿勒泰地区富蕴县国家级草原固定监测点和常规观测点(88°40′18″~89°44′47″ E,46°40′31″~46°58′27″ N),海拔高度为1 020~1 451 m。该区域地属温带大陆干旱气候,冬季寒冷,夏季炎热,春秋季短暂,极端最高气温42.2 ℃,极端最低气温-51.5 ℃,平均气温3.0 ℃[20],日照较为丰富,年日照时数2 869.8 h。年平均降水量为158.3 mm,蒸发量较大,年蒸发量1 743 mm[21]。土壤类型主要为棕钙土,土层厚度为5~10 cm。草地类型为温性草原,轻度超载。植物群落以羊茅(Festuca ovina)、针茅(Stipa capillata)、白茎绢蒿(Seriphidium terrae-albae)、苔草(Carex thibtica)为主。富蕴县天然草地按照利用草地类型与传统牧业生产方式分为夏牧场、春秋牧场和冬季牧场。春秋牧场放牧155 d,退化较严重,所以选其进行探究。
1.1 试验设计
试验样地设在新疆维吾尔自治区阿勒泰地区富蕴县国家级草原固定监测点和常规观测点。样地内2013 年开始采用围栏形式进行全年禁牧;放牧区(放牧强度为重度,即牧草利用率为80%以上)在禁牧区围栏外,属于春秋牧场,放牧牲畜为羊。于2021 年7 月1 日,选择有3 种不同禁牧背景的草地作为研究样地,即未禁牧、禁牧5 年、禁牧9 年的样地,每个样地以对角线方式设置4 个1 m × 1 m 的样方,共计12 个样方(图1)。

图1 研究区概况图Figure 1 Overview of the study area
1.2 样品采集
植被调查为记录每个样方内的物种名称、数量、高度、冠幅和盖度。然后将样方内植物地上部分齐地面分种刈割,并装入样品袋,编号带回实验室,称取鲜重,装入纸袋中,通过烘箱在65 ℃环境下烘干至恒重,用电子天平称量干重(Mettler Toledo,PL2002,精准度为0.01 g),测定草地群落生物量。
土壤调查是在每个样方内,沿对角线方向,参考孔德庸等对表层土壤采样方法[22],利用土钻(直径5 cm)按0 - 10、10 - 20 和20 - 30 cm 土层分别采集3 个土壤样品,采集时除去掉落物,并将每层采集到的3 个土样混匀装在自封袋带回实验室。将一部分土样过0.25 mm 筛用于测定全氮、全磷、全钾;另一部分过1 mm 筛用于测定土壤电导率、pH。利用 1 ∶ 5 土水比悬液酸度计测定土壤pH[23];利用1 ∶ 5土水比浸提液测定土壤电导率[24];利用碱熔法测定土壤全磷[25];利用凯氏定氮法测定土壤全氮[26];利用NaOH 熔融-火焰光度法测定土壤全钾[27]。
1.3 数据处理
1.3.1 物种重要值和多样性指数
重要值(important value, ⅠV)既能表示群落中不同物种的分布情况,也可反映出物种在群落中的功能地位,常用来比较某一物种在群落中的重要性[28-29],其计算公式为:

式中:P为相对密度,H为相对高度,C为相对盖度。
群落物种多样性研究的测度常采用α-多样性和β-多样性体现[30]。α-多样性不仅能反映群落内物种组成和物种丰富度,还可反映物种在群落中分布的均匀程度。β-多样性反映两个群落中共有种与总物种数之间的比例关系,比值越大,则说明两个群落物种间的竞争越小,群落越稳定。本研究采用α-多样性中的Shannon-Wiener 指数(H')、Simpson 多样性指数(D)、Pielou 均匀度指数(J)、Margalef 丰富度指数(DMa)[31-34],比较各群落多样性的变化特征;采用β-多样性中的Sorenson’s 相似性指数(So)和Jaccard’s 相似性指数(Ja)[35-36],比较各群落间相似性特点,各指标计算公式为:
Shannon-Wiener 指数(H′) :

式中:S为样方中的总物种数;N为样方中的总个体数;Ni为第i种植物的个体数;Pi为i种植物个体数占总个体数比例。

式中:S12为两个群落都出现的物种数;S1和S2分别为群落1 和群落2 中的物种数。
采用SPSS 23.0 进行单因素方差分析(One-way ANOVA),并利用Origin 2018 进行制图。
1.3.2 统计分析
采用CANOCO 5.1 中的典范对应分析(CCA)法分析不同禁牧年限植物群落与土壤环境因子的关系。
2 结果与分析
2.1 不同禁牧年限对草地群落特征的影响
2.1.1 不同禁牧年限草地群落的物种组成与功能群重要值
调查发现,研究区共有植物种12 种,隶属6 科12 属,其中禾本科、藜科、豆科植物占所有物种的75%。禾本科植物羊茅(Festuca ovina)、针茅(Stipa capillata)、赖草(Leymus secalinus),藜科植物盐生假木贼(Anabasis salsa)、角果藜(Ceratocarpus arenarius)、猪毛菜(Salsola collina)、木地肤(Kochia prostrata),豆科植物准噶尔岩黄芪(Hedysarum songaricum)、新疆棘豆(Oxytropis sinkiangensis)等分布最广,数量最多。
从功能群的物种组成看(表1),多年生草本中,未禁牧样地纤细绢蒿的重要值较大,针茅的重要值较小。禁牧5 年样地羊茅的重要值较大,纤细绢蒿的重要值较小。禁牧9 年样地羊茅的重要值较大,金丝桃叶绣线菊重要值较小。一年生草本中,未禁牧样地仅有木地肤,重要值为0.04;禁牧5 年样地角果藜重要值较大,猪毛菜重要值较小。禁牧9 年样地无一年生草本植物。围栏封育后,一年生草本的退化指示物种所占比例降低,甚至逐渐消退,如木地肤、角果藜等。随着禁牧年限的增加多年生禾本科重要值所占比例增多,并占据优势地位,说明围栏封育后荒漠草原向着草原化恢复演替的方向发展。
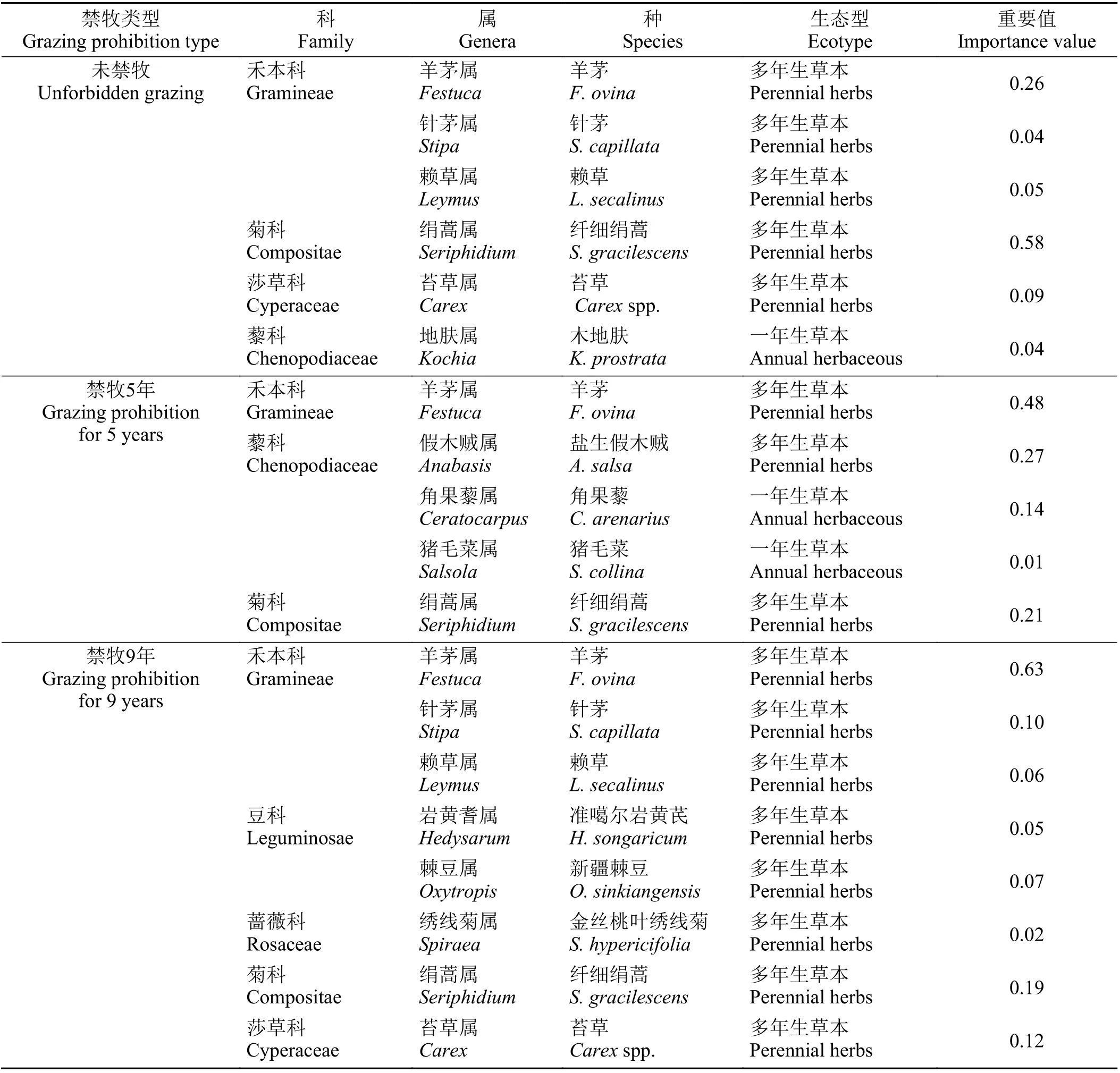
表1 不同禁牧年限草地群落主要物种名录Table 1 List of main species in grassland communities with different grazing prohibition periods
2.1.2 不同禁牧年限草地群落特征
草地群落的平均高度和平均盖度随着禁牧年限的增加有显著增加的趋势(图2),禁牧5 年和禁牧9 年草地群落的平均高度和平均盖度均显著高于未禁牧的草地群落(P< 0.05),禁牧5 年的草地群落平均高度比未禁牧的草地群落高9.89 cm;禁牧9年的草地群落平均高度比未禁牧的草地群落高18.11 cm。未禁牧的草地群落平均盖度最小,为36.25%,禁牧5 年的草地群落平均盖度为47.5%;禁牧9 年的草地群落平均盖度最大,为65%。草地群落的平均密度随禁牧年限的增加有显著降低的趋势,未禁牧的草地群落的平均密度显著高于禁牧5 年和禁牧9 年的草地群落(P< 0.05),未禁牧的草地群落平均密度比禁牧5 年和禁牧9 年的草地群落分别高36.22 和39.71 株·m-2。草地群落的生物量随禁牧年限的增加有显著增加的趋势,禁牧5 年和禁牧9 年的草地群落生物量显著高于未禁牧草地群落(P<0.05)。禁牧5 年的草地群落生物量最大,较未禁牧的草地群落增加60.40 g·m-2;禁牧9 年的草地群落生物量较未禁牧的草地群落增加54.93 g·m-2。

图2 不同禁牧年限对草地群落生态特征的影响Figure 2 Effects and important values of different grazing prohibition years on ecological characteristics of grassland communities
2.2 不同禁牧年限草地群落的多样性变化特征
未禁牧、禁牧5 年和禁牧9 年的草地群落Shannon-Wiener 多样性指数和Simpson 多样性指数整体变化一致,均呈上升趋势(图3)。Shannon-Wiener 多样性指数和Simpson 多样性指数变化表现为禁牧9 年 > 禁牧5 年 > 未禁牧。禁牧9 年与禁牧5 年草地群落的Shannon-Wiener 多样性指数显著高于未禁牧42.53%和27.59% (P< 0.05),Simpson多样性指数显著高于未禁牧47.73%和40.91% (P<0.05)。Pielou 均匀度指数变化表现为禁牧5 年 > 禁牧9 年 > 未禁牧,禁牧9 年与禁牧5 年草地群落的Pielou 均匀度指数显著高于未禁牧42.59%和57.41%(P< 0.05)。Margalef 丰富度指数变化表现为禁牧9 年 > 未禁牧 > 禁牧5 年,禁牧9 年Margalef 丰富度指数显著高于禁牧5 年和未禁牧40.63%和25.00%(P< 0.05)。

图3 不同禁牧方式对草地群落植物多样性的影响Figure 3 Effects of different grazing prohibition methods on plant diversity of grassland community
不同禁牧年限草地群落β-多样性分析结果显示:未禁牧和禁牧5 年草地群落的Sorenson’s 指数和Jaccard’s 指数分别是0.27 和0.22;禁牧5 年和禁牧9 年草地群落相似性最低,Sorenson’s 指数和Jaccard’s 指数分别是0.24 和0.18;未禁牧和禁牧9年的群落相似性最高,Sorenson’s 指数和Jaccard’s指数分别是0.42 和0.56,未禁牧和禁牧9 年的草地群落共有种多,草地群落物种间竞争小。
2.3 不同禁牧年限植物群落与土壤环境因子的关系
将土壤酸碱度、土壤电导率、土壤含水率等6 个环境因素进行最大值标准化预处理,采用物种 × 样地(12 × 12)和环境 × 样地(6 × 12)矩阵数据进行排序分析(图4),由CCA 排序结果发现,前两个轴特征值最大且所包含的信息最多,因此采用前两轴进行分析。

图4 物种数据与环境数据的二维空间排序图Figure 4 Two-dimensional spatial sorting diagram of species data and environmental data
由表2 中信息可知,CCA 排序中所有排序轴均通过蒙特卡洛(Monte Carlo)检验,所有典范轴均达到显著水平(P< 0.05)。由CCA 分析结果可知,土壤酸碱度与第1 轴呈显著负相关关系(P< 0.01),土壤电导率、土壤含水率、全钾和全磷与第1 轴呈极显著正相关关系(P< 0.001),第1 轴各因素相关性表现为土壤含水率 > 全磷 > 土壤电导率 > 全钾 > 土壤酸碱度,其主要反映了土壤含水率、全磷、土壤电导率、全钾、土壤酸碱度的变化,其中土壤含水率、全磷、土壤电导率、全钾对各样点分布具有较高的相关性;CCA 第2 轴和第3 轴与环境因素无相关性,CCA 排序分析第4 轴仅与全氮呈显著正相关关系(P< 0.05),相关系数为0.460 5。CCA 排序前二轴累计解释方差分别为57.40%和77.28%,由此可知,与第1 轴相关的环境因素,即对不同禁牧年限草地群落起主导作用的因素为土壤含水率、全磷和土壤电导率,其余环境因素起辅助作用。

表2 土壤环境因子与前 4 个排序轴的特征值、相关系数、解释方差排序结果Table 2 Comparison of soil environmental factors with eigenvalues, correlation coefficients and explanatory variance of the first four axes
为确定影响植物群落空间分布的主要因素,对6 种环境因素进一步分析(表3)。根据各因素对不同禁牧年限草地群落解释率的大小进行排序,典范对应分析中各环境因素重要性表现为土壤含水率 >土壤酸碱度 > 土壤电导率 > 全磷 > 全氮 > 全钾,其中土壤含水率通过蒙特卡洛置换检验(P< 0.05),各环境因素解释率之和为77.4%,其余22.6%为各因素间的交互作用。因此,土壤含水率(39.7%)为影响不同禁牧年限草地群落差异的主要因素,其次为土壤酸碱度(9.6%)。

表3 CCA 分析各环境因素解释率和显著性检验Table 3 CCA analysis of various environmental factors interpretation rate and significance test
3 讨论
3.1 不同禁牧年限对草地群落生长特征的影响
本研究发现,草地群落的高度、盖度和生物量随着禁牧年限的增加而显著提高(P< 0.05),这与前人的研究结果一致[37-38]。本次野外调查时也发现围栏内的植物普遍的比围栏外的植物个体高大,这可能是由于荒漠草原土壤水分、养分资源缺乏,植物的超补偿生长并不普遍[39]。因此,适度增长围栏封育期,通过草地自然修复,从而促进植被个体的生长发育。野外调查时也发现,随着禁牧年限的增加草地群落的密度显著降低(P< 0.05)。此结果与赵凌平等[40-41]研究的结果一致,随着封育年限的延长群落密度呈逐渐降低趋势,其原因可能是随着禁牧年限的增加,建群种竞争力的增强,进而引起角果藜等一年生草本植物逐渐消退;或者是枯枝落叶层的不断积累,阻碍草本植物的萌发,尤其是影响幼苗期草本植物的光合作用[42],从而导致密度降低。
3.2 不同禁牧年限对草地群落物种多样性的影响
本研究结果表明,禁牧年限与Shannon-Wiener和Simpson 多样性指数呈正相关关系,禁牧可以促进草地群落的多样性指数的增加,其主要原因是禁牧牛羊的啃食和践踏等干扰方式,使得禁牧前因被抑制无法生长的植物有足够的生长空间。随着禁牧年限的增加,草地群落的均匀度指数呈现先增加后减小的趋势,但总体高于未禁牧。有研究指出,禁牧后植物的Simpson 多样性指数和Pielou 均匀度指数比未禁牧区域均有增加[43-44],这与本研究所得出的随着禁牧年限增加草地群落的多样性也有所增加的结果基本一致。从禁牧开始到中期,在一定程度上降低了草地群落的丰富度,使得群落中某些一年生草本和竞争力较弱的植被减少,但后期随着禁牧年限的增加,草地群落的丰富度增加,这与左万庆等[45]研究禁牧对退化羊草草地植物群落物种丰富度随时间表现出先降低后升高趋势的结果一致。因此,禁牧是恢复退化草地的有效措施,能够明显改善植物的生境,为植物群落中消退的物种再次生长创造了条件。从不同禁牧年限草地群落β-多样性分析结果看,未禁牧和禁牧9 年的群落相似性最高,说明未禁牧和禁牧9 年的草地群落共有种多,群落环境较为相似,且草地群落物种间竞争小。
3.3 不同禁牧年限与土壤环境因素的关系
生物量的权衡机制不仅取决于气候条件,还受土壤性质的驱动[46]。通过研究结果发现,影响不同禁牧年限草地植被生长特征的主要土壤环境因素是土壤含水率,其次为土壤酸碱度、土壤电导率、全磷、全氮、全钾。前人的研究中有关于土样因子对植物生长研究的影响并未得到一致性的结论。有研究指出,土壤含水量是影响植物生长及生态系统恢复的关键因子[47-48],也有研究指出影响植物功能群主要因子是土壤全氮含量和土壤含水量[49],还有研究指出影响植物群落盖度的土壤因子为有机质、全氮、全磷[50]。本研究发现全氮不是影响不同禁牧年限草地群落的显著因子,其原因是研究对象不同,张江英等[50]研究湿地群落的植被生长,本身有优越的水源条件,因此含水率不是其植物群落生长的主要环境因子。
4 结论
禁牧使草地群落发生明显变化,其高度、盖度、生物量及生物多样性均明显提高,随着禁牧时间的延长,各项指标在禁牧9 年后比禁牧5 年高,说明本研究区的适宜禁牧时间要长于9 年;禁牧使得群落中一部分竞争力弱的物种消失,降低了群落的密度;从典范对应分析结果中可以看出土壤含水率是影响该区的主要土壤因子。
猜你喜欢
绿色天府(2022年9期)2022-11-19
农业工程学报(2022年13期)2022-10-09
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
物联网技术(2021年6期)2021-07-01
红蜻蜓(2020年12期)2020-06-20
好孩子画报(2019年8期)2019-09-19
人民长江(2019年7期)2019-09-10
发明与创新·大科技(2018年2期)2018-03-17
