
不同中华绒螯蟹放养密度对寒区稻蟹综合种养生产性能及环境因子的影响
2022-03-23 12:30刘金雨赵志刚王世会白庆利李海涛1李明帅
淡水渔业 2022年2期
刘金雨,赵志刚,罗 亮,王世会,张 瑞,郭 坤,白庆利,李海涛1,,李明帅,3
(1.上海海洋大学水产与生命学院,上海 201306;2.中国水产科学研究院黑龙江水产研究所,黑龙江省冷水性鱼类种质资源及增养殖重点开放实验室,哈尔滨 150070;3.东北农业大学动物科学技术学院,哈尔滨 150030)
稻渔综合种养是将水产养殖与水稻种植相结合,基于生物种间互惠互利,生态优势互补,可以提高资源利用率的一种生态循环农业模式。该模式具有稳粮、促渔、提质、增效、环境友好、发展可持续的特点。目前,稻渔综合种养已形成了稻虾、稻鱼、稻蟹、稻鳖、稻鳅五大类典型模式,稻蟹综合种养作为稻渔综合种养的重要模式之一,2019年在全国的应用面积约为1.37×10hm,主要分布在我国北方地区。
黑龙江省是我国水稻种植第一大省,发展稻渔综合种养产业潜力巨大。尽管黑龙江省稻田资源丰富,但其地处高寒地区,水产动物在稻田中的生长周期短,导致适合稻渔综合种养模式的养殖品种较少。中华绒螯蟹()作为我国重要的水产经济动物,是现今黑龙江省发展稻渔综合种养产业的主要养殖品种之一。目前,关于稻蟹综合种养方面的研究主要集中在通过机体生长、水质理化因子、浮游生物、微生物等参数来评价稻蟹综合种养过程中水稻的种植方式、施肥方式和投喂技术等方面的研究。然而,对于黑龙江等高寒地区稻蟹综合种养过程中的相关技术参数尚未见报道。水质和土壤理化因子作为稻蟹共作模式下的重要环境因子,探明其在稻蟹综合种养过程中的变化规律,对于解析该模式在高寒地区的生态效益具有重要意义。为此,本研究通过设置不同扣蟹放养密度,研究寒区稻蟹综合种养模式的生产性能及环境因子变化,旨在为构建寒区稻蟹综合种养模式提供技术支撑和理论依据。
1 材料与方法
1.1 实验材料
实验于黑龙江省绥化市国家级稻渔综合种养示范区进行。水稻品种为“绥粳4号”,实验所用的蟹种来自辽宁盘锦,土壤施用有机肥作为基肥。
1.2 实验设计
根据中华绒螯蟹不同放养密度,设置5个密度处理组,即低密度组(200只/667m),中密度组(350和500只/667m)和高密度组(650和800只/667m),水稻单作为对照组,每组设3次重复,共18个围隔,每个围隔面积为300 m。处理组各围隔间筑田埂,设置高出地面50 cm塑料防逃墙,围隔内距防逃墙0.6 m处开挖环沟,环沟口宽0.5 m,底宽0.3 m,深0.4 m,沟田比为10%,并在对角设进排水口(图1)。扣蟹于2020年6月25日放入围隔稻田,初始放养规格为(21.45±1.45)g。成蟹收获时间为2020年9月10日,水稻收割时间为2020年9月20日。实验期间,每天下午16:00投喂中华绒螯蟹商品配合饲料,投喂量为体重的2%~3%。各密度处理组水稻均采用大垄双行、边行加密的栽培方式进行栽种,实验期间不使用农药。
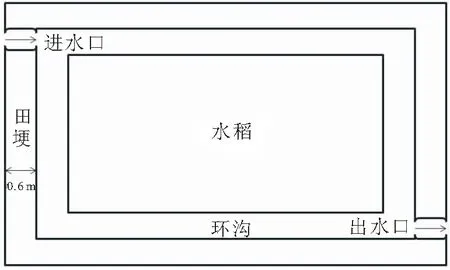
图1 田间工程示意图
1.3 指标测定
1.3.1 水稻的生长参数
从每个围隔随机选取2 m区域收割水稻,稻谷通过脱粒机脱粒晾干后称重。同时,从每个围隔中随机选择1 m区域测量水稻的株高(plant height,PH,cm)、穗长(panicle length,PL,cm)、穗粒数(grain number per rice ears,GR)和千粒重(1 000-grain weight,TKW)。
1.3.2 中华绒螯蟹的生长参数
每个围隔捕获15只雌蟹和15只雄蟹,擦干表面水分进行称重(精确度=0.01 g),计算中华绒螯蟹的增重率(weight gain rate ,WGR)和特定生长率(specific growth rate,SGR)。计算方法如下:
WGR=(-)/×100%
SGR=(ln-ln)/×100%
式中,为扣蟹初始体重(g),为成蟹终末体重(g),为养殖时间(d)。
1.3.3 土壤理化因子
水稻收割后,五点法采集每个围隔的土壤,混匀后带回实验室,风干、研磨、过筛后保存在干燥器中用于测定。总磷和总氮分别采用酸溶-钼锑抗比色法和半微量凯氏法测定,总碳使用N/C分析仪(multi N/C 3100)进行测定。
1.3.4 水体理化因子
水体样品每两周采集测定一次,采样时间为上午10:00~11:00。温度(T)、pH和溶解氧(DO)使用便携式水质分析仪(YSI Professional Plus)进行现场测定,采样水质按照《水和废水监测分析方法》进行测定。化学需氧量(COD)采用酸性高锰酸钾滴定法测定、总磷(TP)含量采用钼酸铵分光光度法测定、总氮(TN)含量采用碱性过硫酸钾消解紫外分光光度法测定、总氨氮(TAN)含量采用纳氏试剂比色法测定、硝酸态氮(NO-N)含量采用紫外分光光度法测定、亚硝态氮(NO-N)含量采用重氮偶合分光光度计法测定、叶绿素a(Chl.)含量采用分光光度法测定。
非离子氨(NH)的计算公式为:
(NH-N)=(100/(1 + EXP(-))/100×;
=10.056 0-0.032 5×(0<<40),
式中,为温度(℃),为水体pH,为水体总氮浓度(mg/L),(NH)为非离子氨的浓度(mg/L),为温度系数。
总无机氮的计算公式为:
(TIN)=(TAN)+(NO-N)+(NO-N),
式中,(TIN)为水体总无机氮浓度(mg/L),(NO-N)为水体硝酸态氮浓度(mg/L),(NO-N)为水体亚硝酸态氮浓度(mg/L)。
1.4 统计分析
实验数据均以平均值±标准误差(Mean±SE)表示,进行单因素方差(ANOVA)分析并进行Duncan氏多重比较,显著性水平=0.05。
2 结果与分析
2.1 中华绒螯蟹生长参数
不同处理组中华绒螯蟹的生长性能见表1。随着放养密度的增加,中华绒螯蟹的终末体重(W)、WGR和SGR整体均呈下降趋势。200只/667m和350只/667m密度组中华绒螯蟹的W、WGR和SGR均显著高于其他处理组。
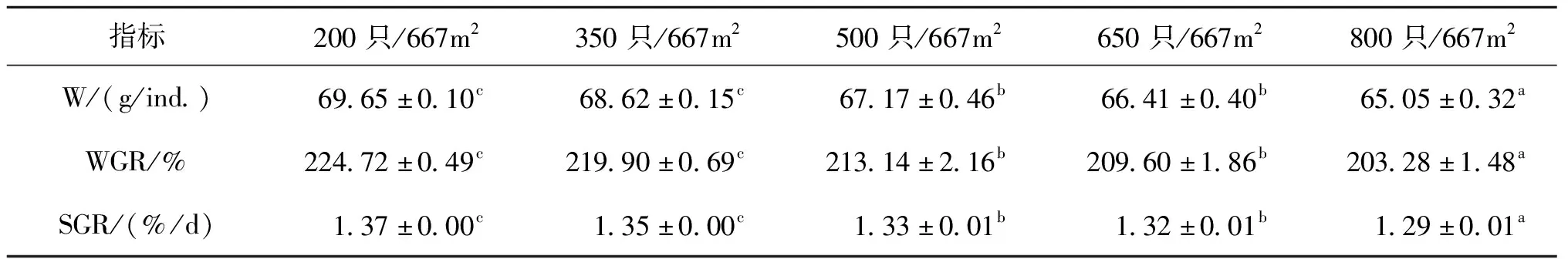
表1 不同处理组中华绒螯蟹的生长性能
2.2 水稻生长性能
不同放养密度条件下水稻的生长性能见表2。水稻的PH、PL、GR及TKW在各组之间均未表现出显著性差异,350只/667m处理组水稻的结实率和产量均显著高于水稻单作组和其他处理组。
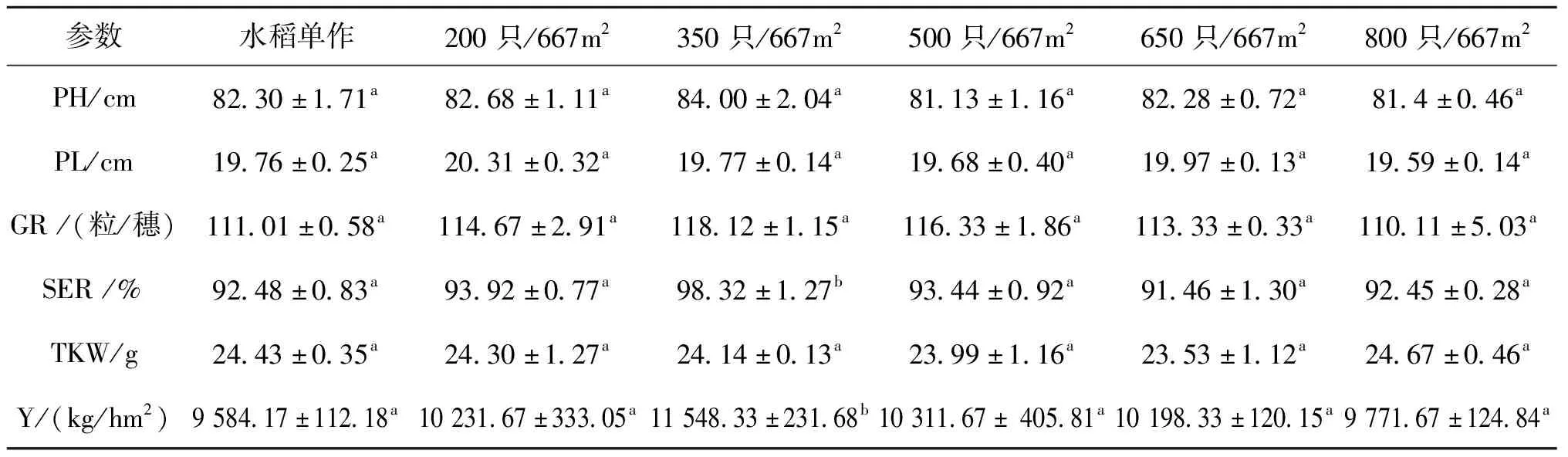
表2 不同组水稻的生长性能
2.3 土壤参数
与水稻单作组相比,各处理组稻田土壤的TP、TN和TC含量均未表现出显著差异(表3)。随着中华绒螯蟹放养密度的增加,各处理组土壤的总磷含量均呈逐渐上升趋势。800只/667m处理组总氮和总磷含量均显著高于350只/667m和500只/667m处理组,与其它组差异均不显著。
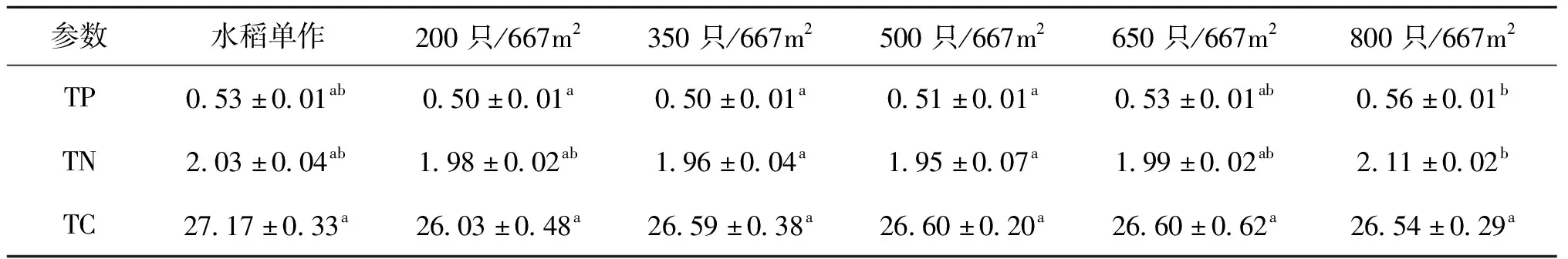
表3 不同组土壤的总磷、总氮和总碳含量(风干重)
2.4 水环境因子
实验期间,各组水体T均呈先上升后下降的趋势,T变化范围17.0~26.0 ℃(图2)。各处理组水体pH总体呈先下降后上升的趋势,波动范围在6.5~7.5,8月3日各处理组水体的pH显著高于水稻单作组(图3)。

图2 各组稻田水体T变化
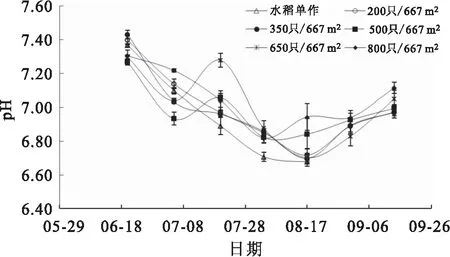
图3 各组稻田水体pH变化
实验期间各组水体DO含量波动较大,各组均在8月份的高温季节处于较低水平,此后DO含量逐渐上升(图4)。7月5日和8月3日,各处理组水体的DO显著低于水稻单作组。8月17日,高密度组(650只/667m和800只/667m)水体的DO显著低于其他组。

图4 各组稻田水体DO变化
稻田水体NO-N和NO-N含量变化分别见图5和图6。养殖前期各组水体的NO-N含量波动较大,波动范围为0.01~0.08 mg/L,从养殖中期开始,各组NO-N含量整体呈显著降低趋势。养殖前期各组稻田水体的NO-N含量均处于较高水平,从6月底开始显著下降,逐渐稳定在0~0.30 mg/L之间。
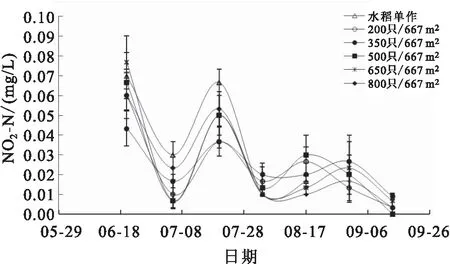
图5 各组稻田水体NO2-N变化
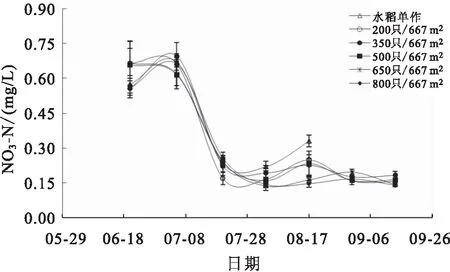
图6 各组稻田水体NO3-N变化
各组稻田水体的TAN、TIN、NH-N及TN含量在实验期间均呈相同变化趋势,养殖前期各项参数显著高于养殖后期,从6月底开始,各项参数均维持在较低水平(图7;图8;图9和图10)。8月17日,高密度组(650只/667m和800只/667m)稻田水体的TAN和TIN含量均显著高于水稻单作组。8月31日,800只/667m处理组稻田水体的NH-N和TN含量均显著高于其他各组。
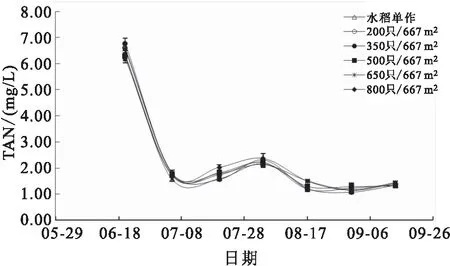
图7 各组稻田水体TAN变化
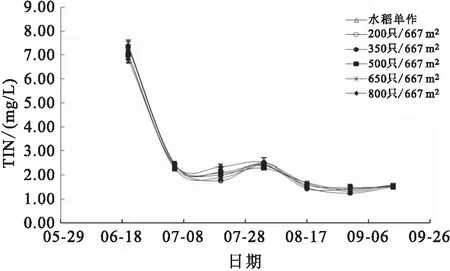
图8 各组稻田水体TIN变化
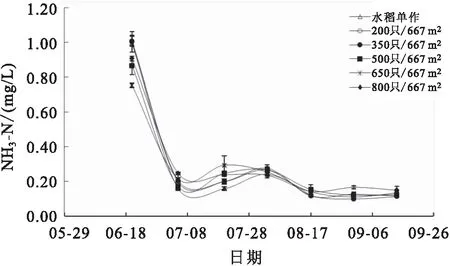
图9 各组稻田水体NH3-N变化
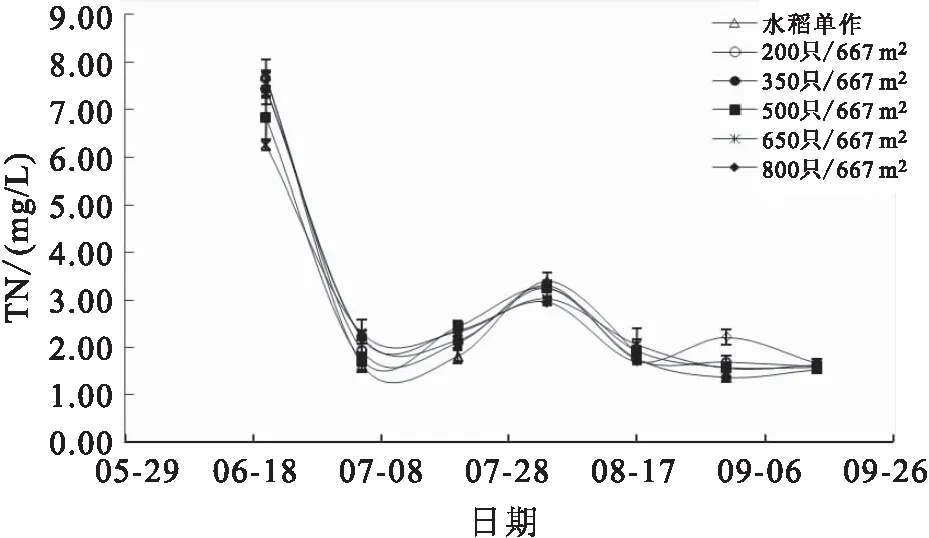
图10 各组稻田水体TN变化
实验期间稻田水体的TP、COD和Chl.a含量变化分别见图11、图12和图13。各组稻田水体总磷含量在实验期间均呈先上升后下降的趋势,且均在8月初上升至最高值,8月中旬之后逐渐稳定在0.2~0.4 mg/L。各组稻田水体COD含量在整个实验过程中变化较大,7月5日、8月3日和8月17日,200只/667m和350只/667m处理组稻田水体COD含量均显著低于水稻单作组。实验期间各组水体的Chl.a含量波动变化较大,各组均未呈现明显的变化规律。

图11 各组稻田水体TP变化

图12 各组稻田水体COD变化
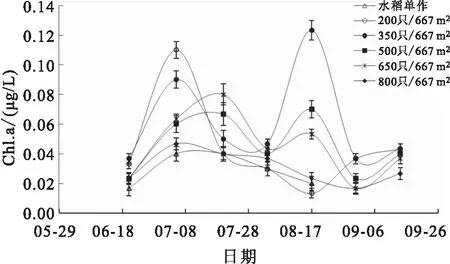
图13 各组稻田水体Chl.a变化
3 讨论
稻蟹综合种养是将水稻种植和中华绒螯蟹养殖相结合的复合农业生产方式,从生态学上来讲,是在稻田生态系统中引入中华绒螯蟹种群后形成的以稻、蟹为主导生物的稻-蟹共生生态系统。水稻与中华绒螯蟹的结合可充分提升系统中物质转化和能量传递的效率。在稻蟹综合种养过程中,根据实际情况投喂饲料,利用稻田的浅水环境,辅以工程技术设施,既种植水稻,又养殖中华绒螯蟹,使稻田的水资源、杂草、小型水生动植物、昆虫以及其他物质和能源充分地被中华绒螯蟹利用,通过中华绒螯蟹的活动,达到为稻田除草、灭虫、疏土和增肥的目的。
本实验结果显示,养殖密度会对中华绒螯蟹的生长产生显著影响(见表1),这与邓梦颖等研究结果一致。本研究采用辽河水系中华绒螯蟹,在黑龙江高寒地区,经过75 d的稻田养殖,500只/667m密度组的中华绒螯蟹从(21.45±1.45)g生长至(66.91±0.78)g,特定生长率为(1.33±0.01)%/d,而吕东峰等在辽宁地区经过120 d的稻田养殖,同样放养密度的中华绒螯蟹从(7.07±0.25)g生长至(96.67±7.64)g,特定生长率为(2.18±0.06)%/d,可能主要因生长周期和温度等气候条件的不同所致。本研究结果显示,稻蟹处理组的水稻产量均高于水稻单作组,这与胡宗云等和安辉等的研究一致。影响水稻产量的因素较多,主要包括遗传因素和环境因素等。本研究结果为放养中华绒螯蟹密度为350只/667m时其水稻产量最高,而郑岩等认为放养密度为696只/667m时水稻产量最高。主要可能因不同区域的实验条件和气候特征差异所致。有相关研究表明,稻蟹共作有利于改善稻田土壤结构。本研究结果显示,中华绒螯蟹放养密度会影响土壤中的总氮和总磷含量,且随着中华绒螯蟹放养密度的增加,总氮和总磷含量逐渐升高,主要原因可能由于中华绒螯蟹活动对土壤的扰动作用及中华绒螯蟹的排泄作用所致。中华绒螯蟹养殖过程中残饵、粪便、脱壳等因素均会导致土壤中的氮磷含量增加,且中华绒螯蟹活动对土壤的扰动造成浑水效应,降低了水体浮游植物含量,一定程度上减少了植物对氮磷的吸收。土壤总碳含量并未随着中华绒螯蟹放养密度的增加而显著上升,可能与土壤微生物对有机质分解导致CO的排放有关。
与湖泊、池塘等水体相比,稻田水体的深度相对较小,其水质环境更易受到气候和人为干扰的影响。水温是水生动物生命活动的重要环境因子,中华绒螯蟹作为变温动物,体温会随环境温度而变化。研究表明,中华绒螯蟹的最适生长温度为20~26 ℃,低于10 ℃或高于30 ℃时摄食活动减弱,生长受限。在实验期间,各组稻田水温均在17.0~26.0 ℃波动,整体处于中华绒螯蟹适宜生长的温度范围。即使在7、8月份的高温季节,水温也未超过30 ℃,一方面原因在于黑龙江地区的高温期较短,另一方面在于高温时期水稻叶片遮盖水面,可有效遮挡太阳光直射,从而避免水体温度的大幅度升高。pH同样作为一项重要的水质理化因子,其稳定性对于养殖和种植的影响较大,过高和过低的pH都会对中华绒螯蟹的生长造成不利影响。中华绒螯蟹适宜的生长pH范围为7.5~8.5,本实验各组稻田水体的pH值在6.5~7.5之间,略低于中华绒螯蟹的适宜生长范围。7月中旬至8月中旬,各组稻田水体的pH值显著下降,该时期为高温季节,在持续降雨,浮游植物的光合作用减弱,种养系统中中华绒螯蟹等生物的呼吸作用导致了pH下降。
溶解氧作为影响水生动物生命活动的关键因子,在养殖过程中其重要性不言而喻。稻蟹综合种养系统中水体的溶解氧来源主要通过日间水生植物的光合作用以及空气中氧气的溶解,而对溶解氧的消耗主要来自中华绒螯蟹、水稻根系、水生植物及微生物的呼吸作用。养殖前期,各处理组水体的溶解氧含量均显著低于水稻单作组,可能与中华绒螯蟹的呼吸作用及微生物对残饵粪便的分解耗氧有关。在养殖过程中,高密度组(650只/667m和800只/667m)水体溶解氧含量在多个时间点均显著低于其他处理组,表明中华绒螯蟹高放养密度会显著降低稻田水体溶解氧含量。渔业水质标准中规定,水产动物在养殖过程中溶解氧含量在任何时候不得低于3 mg/L,而在本实验稻田养殖过程中,在7、8月份高温多雨季节,溶解氧含量有时会有低于3 mg/L的情况出现。因此,在寒区稻蟹综合种养过程中,高温季节可能会有中华绒螯蟹缺氧的现象存在,可通过及时换水等方式进行缓解。中华绒螯蟹在生长过程中除了依赖水体溶氧外,还可从空气中获取氧气,该行为可在一定程度上缓解因水体缺氧导致的生理机能受阻。
水体中的总无机氮包括氨氮、硝酸态氮和亚硝酸态氮,三者之间存在转化关系,其相互转化过程与水体溶解氧含量密切相关。氨氮可通过硝化作用转化为硝酸态氮和亚硝酸态氮,硝酸态氮也可经反硝化作用转化为亚硝酸态氮和氨氮。本研究中,在种养殖初期(7月前),各处理组稻田水体的总无机氮、氨氮和硝酸态氮含量均显著高于其余时间点(见图6,图7,图8),最高时氨氮含量可达6 mg/L以上,可能是因为前期土壤施用基肥所致,随后各项指标因水稻和水生植物的吸收而逐渐降低。整个实验过程,各组之间的总无机氮、氨氮、硝酸态氮和亚硝酸态氮含量均未表现出显著差异,表明在北方寒区的温度条件下,各处理组因投喂饵料导致残饵、粪便等氮营养物质向水体溶解释放的量是有限的,并未因投饵量的增加而显著上升。叶绿素a含量可在一定程度上表征水体中浮游植物的生长和繁殖,本实验中,350只/667m密度组稻田水体叶绿素a含量显著高于水稻单作,表明稻田中放养适当密度的中华绒螯蟹,其残饵粪便和活动行为可显著促进水体浮游植物的生长。水体中氨氮(非离子氨)和亚硝酸态氮含量过高会对养殖动物产生危害,实验期间,相比各处理组养殖初期水体的氨氮(非离子氨)含量和亚硝酸态氮含量,其余时间均处于相对较低水平,但在特定时间内,水体的非离子氨含量仍高于渔业水质标准。在整个养殖过程中可通过适量换水的方式缓解氨氮和亚硝酸态氮对中华绒螯蟹造成的危害。研究表明,水体温度越高,氨氮的毒害作用越强,因此,在高温季节可通过适当增加换水或添加微生态制剂的方式对稻田水质进行调控,降低水体有害氮含量。
氮、磷作为基础物质,在生物代谢过程中发挥着重要作用。化学需氧量可以指示水样中有机物的含量,稻田水体中总氮、总磷及化学需氧量来源主要包括溶解性物质以及浮游生物、残饵、粪便、水稻和水生植物脱落的有机碎屑等。整个实验期间,稻田水体的总氮、总磷及化学需氧量在各组之间差异并不显著,表明稻田中放养一定量的中华绒螯蟹在整个种养过程中不会影响水体主要营养物质的变化。稻田水中总氮、总磷及化学需氧量在不同组之间的个别时间点存在显著性差异,可能与中华绒螯蟹活动及残饵粪便分解导致的差异有关。
4 结论
本研究结果表明,扣蟹放养密度为350只/667m时,中华绒螯蟹和水稻的生长性能最佳、稻田的水质环境较好。本研究认为寒区稻蟹综合种养中扣蟹的最佳放养密度为350只/667m。
猜你喜欢
电脑迷(2022年6期)2022-11-08
澳门月刊(2020年7期)2020-07-14
农村百事通(2019年16期)2019-09-02
农民致富之友(2019年6期)2019-03-25
作文大王·低年级(2019年2期)2019-01-23
投资北京(2018年10期)2018-12-29
学苑创造·A版(2018年9期)2018-10-22
创新作文(1-2年级)(2018年7期)2018-01-22
分析化学(2017年12期)2017-12-25
东坡赤壁诗词(2017年3期)2017-07-05
