
绒螨目线粒体基因组研究进展
2022-03-16 09:34:36冯蕊方颖方瑜褚凌渺苏璇刘璐瑶李飞燕左泽涛丁岚胡彩笑吴默闻孙恩涛
热带病与寄生虫学 2022年1期
冯蕊,方颖,方瑜,褚凌渺,苏璇,刘璐瑶,李飞燕,左泽涛,丁岚,胡彩笑,吴默闻,孙恩涛
1.皖南医学院临床医学院,安徽 芜湖 241002;2.皖南医学院检验学院
绒螨目(Trombidiformes)亦作恙螨目,属节肢动物门(Arthropoda)、蛛形纲(Arachnida)、蜱螨亚纲(Acari)、真螨总目(Acariformes),是一个数量庞大且分散的类群,近年来仍不断发现新种[1]。许多绒螨目螨类具有医学和经济学意义,如皮脂蠕形螨和毛囊蠕形螨可引起皮肤组织的炎性反应,导致毛囊炎、痤疮、酒糟鼻、眼睑缘炎等疾病的发生[2];二斑叶螨在农业生产中危害严重且防治困难,是世界性的重要害虫[3]。
线粒体基因组具有分子量小、基因组成稳定、普遍为母系遗传、较少发生重组和在细胞中拷贝多等特点[4-6],被广泛用于种群遗传学、谱系地理学、分子进化、种系发生和比较及进化基因组学等多方面的研究[7-9],极大地加深了人们对物种进化现象及其过程机制的理解,为系统发育等研究提供了丰富的分子标记。
线粒体基因组的祖征及其系统发育研究对物种的分类归属和进化的决策起着至关重要的作用。而在绒螨目螨类研究中存在着研究类群不全面、重要类群缺少、数据信息发掘不到位等方面的不足。目前在Genbank数据库中,仅报道了28种84条绒螨目螨类线粒体基因组全序列,包括同一物种重复报道的序列以及叶螨总科不同的单倍型序列,见表1。另外,该目的分科分属和系统发育问题正引发学术界的激烈讨论。本文根据绒螨目线粒体基因组研究的现有成果,总结分析了绒螨目线粒体基因组的分子结构特征,综述了绒螨目的系统发育关系并讨论了存在的争议。
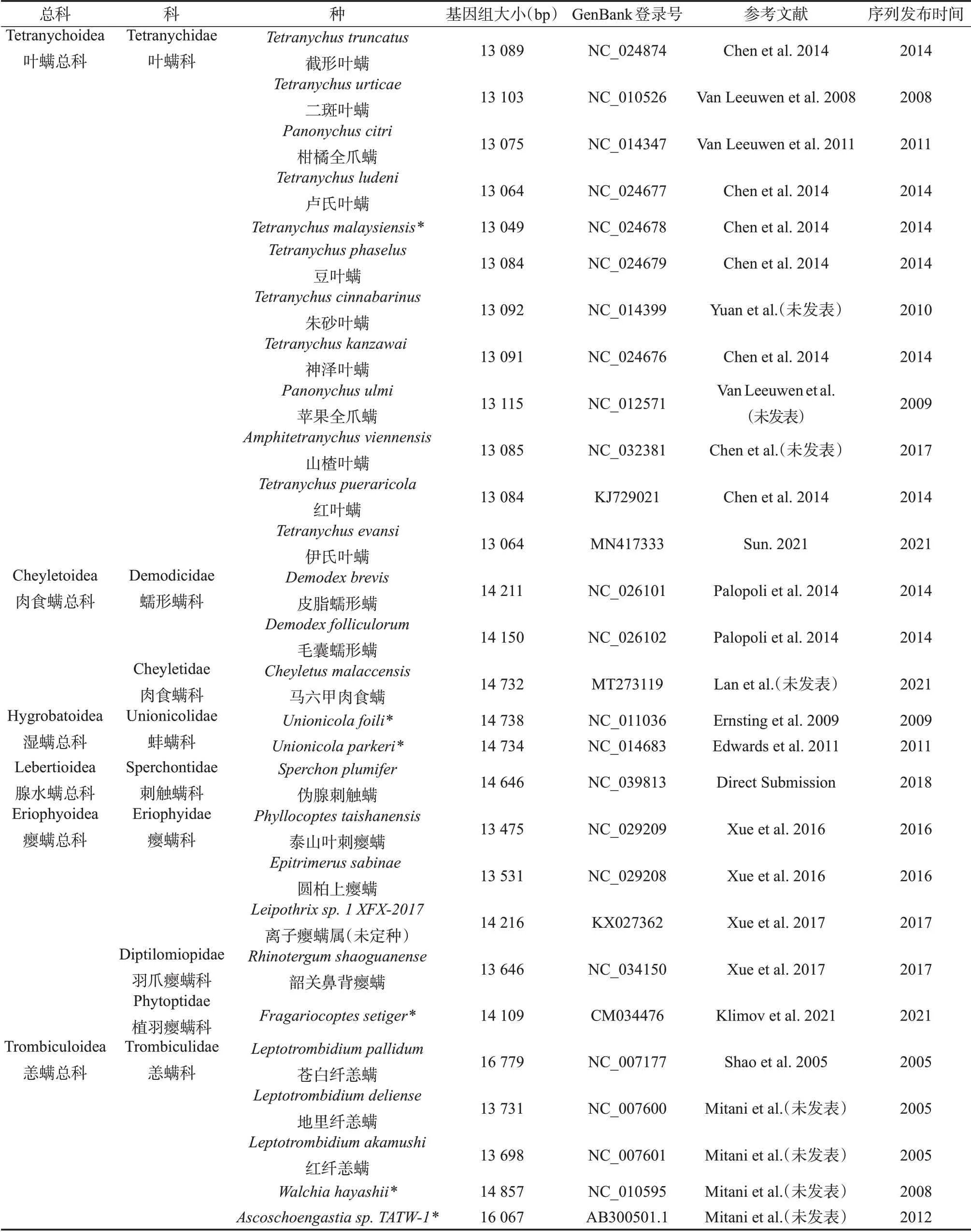
表1 绒螨目已报道种类
1 绒螨目线粒体基因组一般特征及重排分析
绒螨目螨类线粒体基因组长度在13049~16779 bp之间,较已获得的蛛形纲线粒体基因组略小。其中,恙螨科(Trombiculidae)的苍白纤恙螨(Leptotrombidium pallidum)的线粒体基因组由于含有2个rrnL基因和1个PrrnS序列(rrnS的假基因),长达16779 bp[10]。叶螨科(Tetranychidae)的线粒体基因组则属于绒螨目中较小的一类,长度为13049~13115 bp[11]。
与其他后生动物相似,绒螨目螨类线粒体基因组在碱基组成上偏向A和T。其中,红纤恙螨(Leptotrombidium akamushi)A+T含量最低(67.5%)[12],苹果全爪螨(Panonychus ulmi)最高(85.6%)[11]。此外,A+T含量通常在同科或同属物种间相近,如叶螨科4种叶螨线粒体基因组的A+T含量平均值为84.9%±0.66%[11]。
绒螨目螨类线粒体基因组为共价闭合的双链环状结构,与典型后生动物相似,多数具有37 个基因,包括13 个蛋白质编码基因(protein-coding genes,PCGs),2 个rRNA 基因和22 个tRNA 基因。在大部分后生动物中,37个线粒体基因的排列顺序相对保守,但在绒螨目已测序的螨类中,其基因组呈现出高度的重排,甚至在同一属内的不同种之间也存在重排现象。如在恙螨科中,与节肢动物祖先相比,红纤恙螨和地里纤恙螨(Leptotrombidium Deliense)的17 个基因发生了易位,8 个基因同时发生易位和倒位。而苍白纤恙螨线粒体基因组是一种较为特殊的类型,除具有37 个基因外,还有四个控制区以及额外的rrnL基因和PrrnS基因,Shao 等[12]在复制-删除模型基础上结合非同源性的基因组间重组模型,推测苍白纤恙螨的线粒体基因组可从其他2 种纤恙螨属(Leptotrombidium)基因组演变而来。此外,已公布的恙螨科2 种非纤恙螨线粒体基因(Walchia hayashii,Ascoschoengastia sp.)排序各不相同,各自代表一种排列方式。在蚌螨科(Unionicolidae)中,两种水螨(Unionicola foili,Unionicola parkeri)基因组相较节肢动物祖先也发生了高度重排。且这两种水螨的基因排列模式在属内也存在不同,与U.foili相比,U.parkeri的trnW和trnC基因的排序发生了易位[13-14]。研究发现蚌螨属(Unionicola)存在高度分歧的cox1 序列,推测该属可能存在独特的基因排列特征[13]。而在叶螨科中,虽然相较于祖先线粒体基因重排程度很高,但在科内保持一致[11,15]。瘿螨总科(Eriophyoidea)则具有相对于节肢动物线粒体基因祖先排列模式的最小重排程度和最大保留程度[16-17]。见图1。
2 绒螨目线粒体蛋白质编码基因特征
绒螨目螨类PCGs 广泛使用ATN(ATG,ATT,ATA,ATC)作为起始密码子,少数使用非标准的起始密码子,如cox1 基因(Leipothrix sp.)、nad5 基因(U. parkeri, U. foili)、nad4L基因(Leipothrix sp.)使用TTG,cox1 基因(Phyllocoptes taishanensis)、atp8基因(P. taishanensis)使用CTG。终止密码子大多为TAA或TAG,部分螨类在蛋白质翻译时还使用T或TA 作为转录终止信号,后者在转录后可能经聚腺苷酸化最终形成完整的终止密码子[18]。然而,在U. foilii的nad4L基因中未发现终止密码子,Ernsting 等[13]推测在U. foilii线粒体基因组中nad4L和nad5组成融合基因,进行融合蛋白的表达。密码子偏性(codon bias)在后生动物中普遍存在,这种偏性也在已测得的绒螨目螨类线粒体基因组中发现。例如,柑橘全爪螨(Panonychus citri)相对同义密码子频率(RSCU)显示,第三位为A或T的密码子总是过度使用[11]。
3 绒螨目线粒体rRNA基因特征
通常,基因簇rrnS-trnV-rrnL在许多后生动物中高度保守。但在目前已测得的绒螨目螨类中,只有瘿螨总科保持着这种基因簇的形式,而恙螨科3 种纤恙螨的rrnL和rrnS直接相连,蚌螨科(U. foili和U. parkeri)和恙螨科(W. hayashii和A. sp.)则显示rrnL和rrnS之间伴有nad1 基因和1~2个tRNA的插入,在恙螨科的苍白纤恙螨中,还额外发现了一个rrnL和PrrnS[12]。而叶螨科的4 种叶螨的2 个rRNA基因相距甚远,插入了5 个PCGs 和5 个tRNA 基因[11],见 图1。Edwards 等[14]认 为2 个rRNA 之 间nad1基因的插入,可作为绒螨目中蚌螨科与恙螨科之间的共同衍征,但未发现该特征存在于叶螨科和恙螨科纤恙螨属的螨类中。同时,研究发现多数节肢动物线粒体rrnL和rrnS基因均由N-链编码[19-22]。相反,在绒螨目中,恙螨科纤恙螨属(L.pallidum、L. akamushi和L. deliense)[10]和叶螨科(Tetranychus urticae,Tetranychus cinnabarinus,P. ulmi和P.citri)的rRNA 基因均在J-链上编码[11]。此外,由于rRNA 的二级结构主要由多种茎区和环区组成,在不同的物种中茎环的数目、大小和位置存在差异,因此可通过预测其二级结构,选择合适的基因区段来进行同源性比较,然而关于这方面研究在绒螨目中仍很匮乏。

图1 绒螨目线粒体基因组的排列
4 绒螨目线粒体tRNA基因特征
在后生动物中,典型的线粒体tRNA 基因具有三叶草型二级结构,即包括7 bp 的氨基酸接受茎,5 bp 的反密码子茎,T 臂和D 臂。真螨总目中,tRNA二级结构呈现出大量的D臂或者T臂缺失,在绒螨目中这种现象尤为显著:叶螨科的22个线粒体tRNA 中有19 个二级结构缺少D臂或T臂,甚至在一些种中,trnF和trnN同时缺少D臂和T臂[11,15];蠕形螨科(Demodicidae)有15 个tRNA 缺少D臂或T臂,有5 个同时缺少D臂和T臂[23];恙螨科有18 个tRNA缺少D臂或T臂[10,12-13];蚌螨科有15个tRNA缺臂[13-14]。瘿螨总科中,四个已报道的种都有缺臂的tRNA,虽然其重排程度在真螨总目中最低,但缺臂程度却是最高的。四种瘿螨的22 个tRNA 中16 个缺失D臂或T臂。另外,在绒螨目中,缺臂的特征在科内基本上保持一致,而科间存在差异。
5 绒螨目线粒体控制区
控制区是后生动物线粒体基因组中富含AT 的最大非编码区(large non-coding region,LNR),通常与rrnS基因相邻,形成多个潜在的茎环结构。因大多数节肢动物的AT富集区广泛存在一系列保守的序列元件和二级结构特征,这段区域被广泛认为是线粒体基因组复制和转录相关的控制元件[24-25]。
线粒体基因组中非编码区的保守性普遍低于编码区。在已测得的绒螨目线粒体基因组中,LNR的长度、位置和序列变异显著。真螨总目螨类大多含有一个LNR,而绒螨目3 种螨类(L. akamushi、L.deliense和U.foili)具有2个LNRs,苍白纤恙螨则有4个LNRs。LNR序列种内相似性大于种间相似性,证明同一物种的多个LNRs 之间为协同进化[12]。但值得注意的是U.foili的2 个LNRs 序列之间差异显著。此外,复制-删除模型无法解释纤恙螨属线粒体基因组中转录方向相反的LNR 之间的协同进化现象,而基因转换可对此作出较好解释[12]。多数真螨总目螨类线粒体控制区均可形成茎环结构,或存在一些保守序列,但4 种叶螨的控制区序列存在明显差异,仅有数个较短保守序列。同时,4 种叶螨的AT富集区均可形成稳定茎环的二级结构,全爪螨属(P. citri和P. ulmi)有2 个稳定的茎环结构,而叶螨属(T.urticae和T.cinnabarinus)仅有1 个。比较发现,LNRs 区域的保守序列或茎环结构即使在绒螨目亲缘关系很近的物种间也少有相似性。绒螨目螨类LNR 中较长的有W.hayashii(1403 bp)、A.sp.(1205~1234 bp)和U.parkeri(977 bp),但叶螨科4种叶螨的AT 富集区较短,仅44~57 bp,这种长度的变异在绒螨目中十分明显。除LNR大小外,其相对位置在绒螨目基因组中也具有一定变异:叶螨科4种叶螨线粒体AT 富集区位于cox1 和nad3 基因之间,蚌螨科2 种蚌螨和恙螨科中W.hayashii的LNR两侧分别是trnL1 和trnH基因以及trnS1 和trnT基因[26],见图1。
6 绒螨目系统发育关系
系统发育可以阐述物种之间的进化和亲缘关系。近年来,分子技术的高速发展使系统发育关系倾向于用分子建树阐述。由于线粒体基因组具有分子量小、拷贝数多等特点,常常被用于系统发育研究。同时可以结合形态学特征,探讨物种的分类归属问题。
蜱螨亚纲在较高水平(目或总股)的亲缘关系一直存在争议。有学者将蜱螨亚纲分为真螨总目(Acariformes)和寄螨总目(Parasitiformes),大致对应着两个总目螨类的生活方式和生活环境,但是很多蜱螨学家认为两个谱系之间并没有很深的密切联系,支持蜱螨亚纲为多系。在之前研究中,真螨总目、寄螨总目常常恢复为单系,但蜱螨亚纲的单系没有得到恢复[27-28]。
真螨总目下设绒螨目和疥螨目(Sarcoptiformes)。根据最新的分类系统,绒螨目分类见图2。

图2 绒螨目已测序种类分类
研究表明,在瘿螨总科线粒体基因组被测序之前,由于绒螨目序列信息不足以及关键物种分类序列缺失,导致在之前的系统发育研究中,绒螨目往往被恢复为单系[15,23]。但在包括了瘿螨的蜱螨亚纲系统发育分析中,在真螨总目内,支持疥螨目的单系性却不支持绒螨目的单系性,且瘿螨单独聚为一支[29]。
瘿螨总科属绒螨目(Trombidiformes)、前气门亚目(Prostigmata)、真足螨总股(Eupodides),现有4种序列在Genbank上报道。大多数瘿螨有较高的寄主植物特异性,目前的形态学和分子研究都支持瘿螨总科的单系性,但瘿螨总科在蜱螨亚纲的系统发育地位仍然存在争议。由于具有不愈合的螯肢、退化的口针结构、大体缺乏成对的侧腺体、侧毛只有1~2对等特征,认为瘿螨的类群是绒螨目下的一个总科,设为瘿螨总科。但在某种意义上,瘿螨归入疥螨目也可以说得通。即使没有作为疥螨目鉴定特征的气门系统,瘿螨也有很多疥螨目的特征:比如足有羽状爪,无气管系统,以及某些原始种群中缺少侧腺体等[29]。
Xue等[17]为了推测瘿螨总科在蜱螨亚纲中的系统发育地位,选取了包括瘿螨在内的110 个蜱螨种类的线粒体基因组序列,构建了蜱螨亚纲高级阶元的系统发育树。此外,还选取231个蜱螨种类(包括25 个瘿螨种类)的18S 核基因序列构建系统发育树。两套数据的分析结果都支持以往研究中瘿螨总科的单系性,但不支持绒螨目的单系性。依据不同的数据分析方法,得到两种略有差异的结果:一种是支持瘿螨和疥螨目聚类,另一种则支持瘿螨是“疥螨目+非瘿螨的绒螨目”的姐妹群。但这两种结果都表示瘿螨总科可能不适于放在绒螨目下。还有一些研究基于线粒体基因组氨基酸序列所建的系统发育树恢复了传统的绒螨目祖先分支关系,但在这种情况下,瘿螨总科会和异气门总股(Eleutherengonides)聚合[30]。
有学者提出一种假说,将瘿螨总科置于内气门亚目(Endeostigmata)下,和线美螨科(Nematalycoidae)位置相邻[31-32]。但内气门亚目由几种寄居在土壤的科属组成,和瘿螨的生活环境大相径庭。Arribas等[30]基于瘿螨总科引起的绒螨目的分类争议,采用蜱螨亚纲一百多条具有代表性的线粒体基因组,进一步探讨蜱螨亚纲系统发育关系时,其一种系统发育树显示,瘿螨总科从真螨总目中最先分离,而后分为绒螨目和疥螨目,反对绒螨目的单系性。研究还提出除去瘿螨总科的绒螨目内部总股之间的分类也只是部分符合之前所述的形态假设,需要更深地了解整个瘿螨-绒螨目关系的复杂性,同时也要注重真足螨总股(Eupodides)和大赤螨总股(Anystides)的非单系性。
7 展望
由于在动物进化过程中,线粒体基因很少出现重排。当重排出现时,可为阐明绒螨目物种间深层次的系统发生关系提供重要参考。例如,绒螨目的蚌螨科和恙螨科(除A.sp.外)共享nad4-trnR-trnVnad6-cytB基因的排列,以该区域为基础的基因结构支持他们的近缘关系[14]。
在已测得的绒螨目线粒体基因组tRNA二级结构的氨基酸接受臂、D臂、反密码子臂和T臂均存在一定数量的碱基错配。这些错配严重影响到tRNA基因的预测,因为错配的tRNA基因与随机DNA序列差异不明显,导致tRNA 基因的准确注释存在较大困难[26]。而且,高度截断的tRNA 也大大增加了注释的工作难度,很可能在软件预测中出现难以置信的二级结构或者在预测过程中被忽略。后生动物线粒体基因组中tRNA的二级结构和反密码子在低级分类(科或属)相对保守。因此,除线粒体基因序列特征外,tRNA 二级结构特征也具有潜在的系统进化信息,尤其在非典型tRNA 二级结构广泛存在的绒螨目螨类中。例如,通过对蚌螨科2 种蚌螨(U. foili、U. parkeri)和恙螨科2 种恙螨(L. akamushi、W.hayashii)tRNA 基因比对发现,多数tRNA 具有保守的反密码环序列,且同一科内所得的二级结构完全一致[14]。同时,利用线粒体基因组tRNA 基因的二级结构和反密码子在低级分类的保守性,可避免近缘种中tRNA基因的错误注释。
对于绒螨目瘿螨总科的研究已经持续了一个多世纪,但其系统发育研究较少,瘿螨总科的分类地位在形态祖征和分子信息上缺少足够的系统发育证据。绒螨目的研究也主要集中于新种的描述,其系统发育关系以及分类问题也因为缺乏重要分类类群的物种信息而困难重重,如吸螨总科尚未见全序列报道。因此,加强绒螨目线粒体功能基因组学的研究,获得新的重要类群物种信息对于阐明瘿螨总科系统发育位置以及解决绒螨目的系统发育问题具有重要意义。
猜你喜欢
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
华东师范大学学报(自然科学版)(2021年6期)2021-01-01 11:52:58
生物学通报(2020年11期)2020-10-22 01:20:20
能源研究与信息(2020年4期)2020-02-19 06:45:06
中成药(2018年7期)2018-08-04 06:04:10
中国中医药图书情报(2014年6期)2015-03-30 11:55:50
生命科学研究(2014年1期)2014-04-29 00:44:03
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
中国烟草学报(2012年2期)2012-04-09 06:44:56
