
饲料中添加苯甲地那铵对肉仔鸡骨骼肌发育的影响
2022-03-12 07:57:00黄振吾胡晨辉杨艳艳吕增鹏
饲料工业 2022年4期
■金 松 黄振吾 胡晨辉 杨艳艳 吕增鹏
(1.常州市动物疫病预防控制中心,江苏 常州 213000;2.南京农业大学动物科学技术学院,江苏 南京 210095;3.北京市饲料工业协会,北京 100107;4.中国农业大学动物科学技术学院,北京 100193)
随着饲用抗生素在动物生产中的限制使用,越来越多的植物提取物被应用于家禽饲料中。植物提取物中大多含有苦味物质,鸟类能够很好地识别食物中富含的多酚类苦味有毒物质[1-4]。虽然家禽的味觉在很大程度上演变为对饲料营养的自然反应,但是家禽在被动的学习逃避试验中能够很好地识别邻氨基苯甲酸甲酯(苦味剂)。因此,研究苦味物质对肉仔鸡骨骼肌发育和生长性能的影响,对于指导具有苦味的植物提取物添加剂在家禽饲料中的合理运用具有重要意义。苯甲地那铵(DB)是一种苦味激动剂,被广泛用于苦味生理效应相关研究[5]。尽管鸡只有三种苦味受体:ggTas2R1、ggTas2R2和ggTas2R7,但同样表现出苦味敏感性[6]。前期研究表明,DB和大豆提取物——染料木黄酮均可以激活心脏、肝脏等器官的苦味受体表达[7]。并且,饲料中添加DB能够损伤空肠上皮的形态,诱导苦味受体相关的细胞凋亡,对肉仔鸡肠道的发育产生不利影响[8]。据此,推测DB可能也会对肉仔鸡的骨骼肌发育和蛋白质代谢产生不利影响。本试验在日粮中添加梯度浓度的DB,旨在阐明其对肉仔鸡的骨骼肌发育的作用,研究对于富含苦味物质的植物提取物在家禽饲料中的应用具有重要参考价值。
1 材料与方法
1.1 试验材料
苯甲地那铵:纯度为98%(阿达玛斯试剂有限公司,南京)。
1.2 试验动物及日粮
试验动物选用1日龄黄羽肉鸡,参照中国鸡饲养标准(2004)配制0~4 周龄和5~8 周龄合成粉状全价料,基础日粮配方及营养水平见表1。
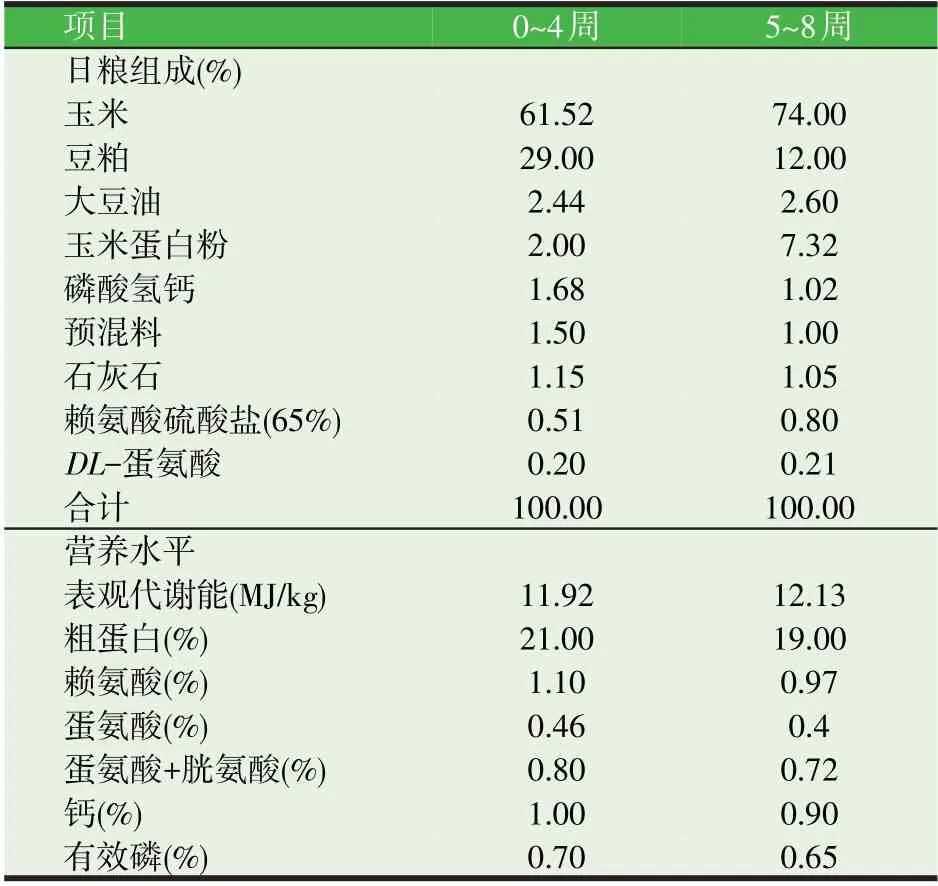
表1 基础日粮组成和营养水平(风干基础)
1.3 试验设计与饲养管理
动物饲养试验地点为南京农业大学动物实验基地,试验流程严格按照南京农业大学动物福利委员会相关规定进行。试验选用体重相近的1日龄黄羽雄性肉仔鸡240只,随机分为4组:对照组(CON);低剂量苯甲地那铵组(LDB);中剂量苯甲地那铵组(MDB);高剂量苯甲地那铵组(HDB),每组5个重复,每个重复12只鸡。所有肉仔鸡在负压通风的鸡舍中笼养,每笼12只鸡。对照组饲喂玉米-豆粕型基础日粮,LDB、MDB组和HDB组在基础日粮中分别添加5、20 mg/kg和100 mg/kg DB。试验周期为8周。肉仔鸡在试验期间自由饮水,饲养管理按快大型黄羽肉鸡管理的常规程序进行。
1.4 样品采集
试验第8 周结束,所有鸡称量体重后,每个重复中随机选择2 只鸡于翅静脉处采集血液。收集的血液4 ℃静置,12 h后在低温离心机中以3 000 r/min离心10 min得到血清,随后将血清样品保存在-20 ℃直至分析。肉鸡采集完血液后通过颈静脉放血处死,立即分离右侧胸肌组织后称重,用于计算胸肌指数。于右侧胸肌组织相同位置分别取大小相近的组织块固定在4%多聚甲醛溶液中,待组织学分析。采集肉鸡左侧相同位置胸肌组织立即置于液氮中冻存,随后保存于-80 ℃至样品检测。
胸肌指数(%)=右侧胸肌重(g)/活体重(g)×100
1.5 血清生化指标测定
使用Spark 20 M酶标仪(Tecan公司,瑞士)和商用试剂盒(A113-1-1、A112-1-1、C010-2-1、C009-2-1、A028-2-1、C012-2-1、C011-2-1、A111-1-1 、A110-1-1、A045-2-2,南京建成生物工程研究所,南京)检测低密度脂蛋白胆固醇(LDL-C)、高密度脂蛋白胆固醇(HDL-C)、谷草转氨酶(AST)、谷丙转氨酶(ALT)、白蛋白(ALB)、尿酸(UA)、肌酐(CREA)、总胆固醇(TCHO)、三酰甘油(TG)和总蛋白(TP)等血清生化指标。
1.6 胸肌组织学分析
4%多聚甲醛固定的胸肌组织经修整后,用梯度浓度的乙醇脱水,二甲苯透明,并进行石蜡包埋。用切片机(Leica Biosystems, Buffalo Grove, 美国)从每个石蜡块中获得5 μm的切片。用二甲苯除去石蜡,梯度乙醇复水后,苏木精和伊红(H.E.)染色。使用Velleman和Nestor 描述的方法测量组织切片的肌纤维宽度[9]。使用带有Olympus Magna Fire数码相机的Olympus XI 70显微镜拍摄,并使用Image-Proplus 6.0进行分析。
1.7 基因表达检测
取冻存的100 mg 胸肌组织样品加入1 mL Trizol(Invitrogen Life Technologies,Carlsbad,美国)提取液,严格按照试剂盒说明书提取总RNA。提取的总RNA用核酸分光光度计(AG 22331,Eppendorf,Hamburg,德国)检测其纯度和浓度,在260/280 nm 下测定值大于1.8认为RNA提取质量较好。根据试剂盒说明书采用M-MLV cDNA试剂盒(Invitrogen Life Technologies)进行反转录反应。反转录产物按照1∶1 稀释后备用。选用SYBR®Select Master Mix PCR Kit(R323-01,诺唯赞生物公司,南京)进行RT-PCR(Real-TimePolymerase Chain Reaction)检测,反应仪器为7500荧光检测系统(Applied Biosystems,Foster City,California,美国)。目标基因和内参基因GADPH的引物由生工生物工程(上海)有限公司合成,引物序列如表2所示。PCR反应程序如下:变性95 ℃30 s,循环次数40,熔解曲线:95 ℃15 s,60 ℃60 s,95 ℃15 s。使用熔解曲线验证引物的扩增效率后,针对内参基因GADPH 标准化,并通过2-ΔΔCt方法分析相对基因表达水平。

表2 基因的引物序列
1.8 数据处理
采用SPSS(version 25.0)软件中的one-way ANOVA程序进行统计分析。组间差异显著时,以Duncan’s法进行多重比较,P<0.05 为差异显著,P<0.10 判定为具有差异的趋势。试验结果描述为“平均值±标准误”。
2 结果与分析
2.1 生长性能和胸肌指标(见表3)
如表3所示,试验第8周末,相比于CON组,饲料中添加5、20 mg/kg 和100 mg/kg DB 显著降低肉仔鸡的体重和胸肌重(P<0.05)。然而与CON组相比,饲料中添加DB对胸肌指数没有显著差异。

表3 DB对肉仔鸡生长性能和胸肌指数的影响(n=10)
2.2 血清生化指标(见表4)
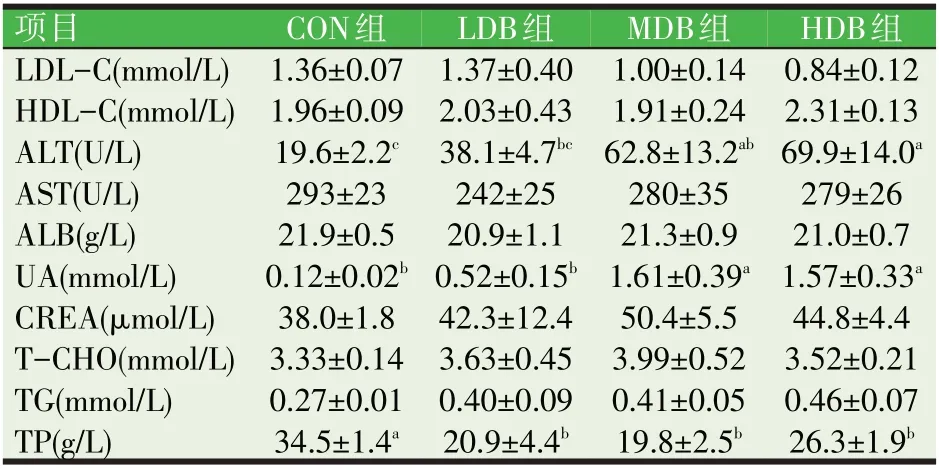
表4 DB对肉仔鸡血清生化指标的影响
如表4所示,与CON组相比,随着饲料中DB添加量的增加,肉鸡血清中谷丙转氨酶(AST)和尿酸(UA)水平逐渐升高后下降。MDB组和HDB组血清中尿酸水平均显著高于CON组和LDB组(P<0.05)。然而,随着饲料中DB添加量的增加,肉鸡血清中总蛋白(TP)逐渐降低后升高。饲料中添加5、20 mg/kg和100 mg/kg DB显著降低肉仔鸡血清中总蛋白水平(P<0.05)。此外,与CON组相比,饲料中补充不同含量DB,并未显著改变肉鸡血清中其他生化指标。
2.3 胸肌组织学分析(见图1)
如图1A所示,饲料中添加DB可使肉仔鸡胸肌纤维间隙增大,肌纤维宽度明显降低。统计结果表明,LDB 组胸肌纤维直径较对照组显著降低,MDB 组和HDB 组胸肌纤维直径较CON 组和LDB 组显著降低(P<0.05,图1B)。然而DB对肉仔鸡胸肌单位面积(胸肌纤维密度)没有显著影响(图1C)。

图1 DB对肉仔鸡胸肌组织学形态的影响
2.4 胸肌苦味受体相关基因mRNA表达(见图2)
如图2所示,饲料中添加DB对胸肌苦味受体基因ggTas2R1、ggTas2R2的mRNA表达水平没有显著影响。LDB组和CON组胸肌ggTas2R7基因mRNA表达水平没有显著差异,然而MDB 组和HDB 组ggTas2R7基因mRNA 表达水平较CON 组和LDB 组显著提高(P<0.05)。HDB组胸肌苦味受体下游PLCβ2基因mRNA表达水平较CON、LDB组和MDB组有显著提高(P<0.05)。同时,LDB、MDB组和HDB组处理相对于CON组显著提高肉仔鸡胸肌中IP3R3基因mRNA表达水平(P<0.05)。
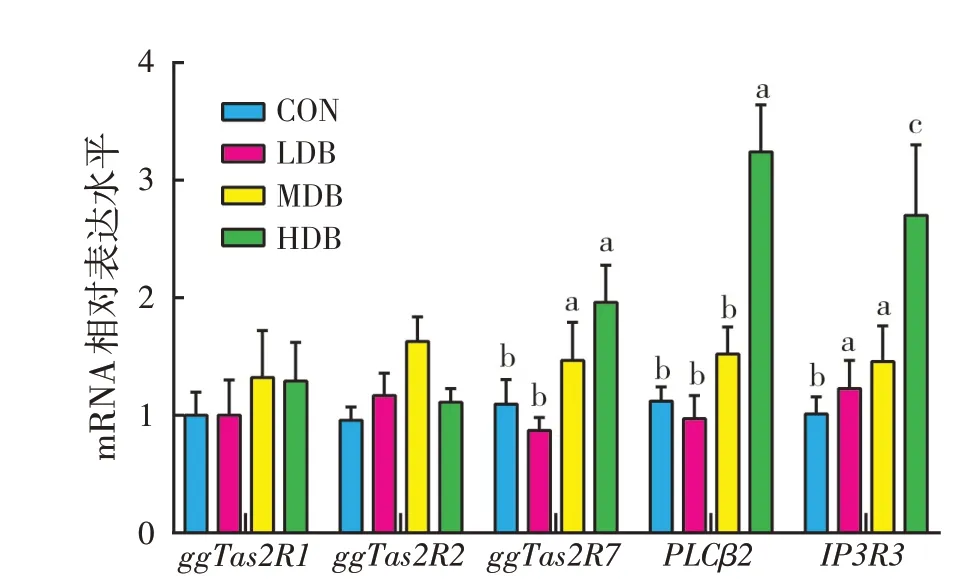
图2 DB对肉仔鸡胸肌苦味受体相关基因mRNA表达的影响
2.5 胸肌肌纤维发育和蛋白质代谢相关基因mRNA表达(见图3)

图3 BD对肉仔鸡胸肌肌纤维发育和蛋白代谢相关mRNA表达的影响
如图3A 所示,与对照组相比,饲料中添加DB 对肉仔鸡胸肌基因MSTN、Pax-7、MyoG和MyoD的mRNA 相对表达水平没有显著影响。如图3B 所示,MDB 组和HDB 组相比于CON 组显著提高胸肌中Atrogin-1 的mRNA 表达水平。HDB 组胸肌中MuRF1的mRNA 表达水平较CON 组和LDB 组显著上升(P<0.05)。与CON 组相比,所有DB 组对肉仔鸡胸肌中INSR、m-TOR、AKT的mRNA表达没有显著影响。
3 讨论
3.1 DB对肉仔鸡生长性能的影响
酚类、黄酮类、萜类和硫苷类等功能性植物提取物具有苦味[10]。植物进化从自我保护角度合成苦味物质,其目的是本身不被吃掉,因此这些味道对动物来说是难以接受[11]。大多研究往往关注这些功能性添加剂的有益生物学功能,但高剂量的苦味物质能够通过激活苦味受体对机体发育造成不利影响[12]。DB是化工合成的苦味剂,作为添加剂已被广泛应用于饲料生产中。研究发现,长期摄入低剂量DB 不会对大鼠或猴子表现出相关毒性[13]。本研究中使用安全剂量的DB作为苦味受体激动剂,探究其通过苦味受体信号通路对肉仔鸡骨骼肌发育与代谢的调控作用,对于苦味性添加剂在家禽饲料中应用具有指导价值。Avau等[14]研究发现,胃内给药DB 能够减轻肥胖小鼠的体重,这与饲粮中添加DB显著降低了雄性肉仔鸡的生长性能结果一致。Avau等[14]认为DB暴露导致体重减轻的可能原因是DB抑制平滑肌收缩,从而影响消化率和营养吸收。Jiang等[8]研究发现,饲粮中添加DB会激活肠道苦味受体,并损伤仔鸡肠道发育,可能是使仔鸡生长性能降低的原因之一。骨骼肌是动物机体的重要组成部分,也是肉鸡产生经济效益的主要部位[15]。仔鸡肌纤维数量在胚胎期就已经确定,孵化后肌肉发育主要依赖于蛋白沉积引起的肌纤维直径增加[9]。本研究发现,饲粮中添加DB会显著降低肉仔鸡胸肌重和肌纤维直径。推测DB可能通过影响蛋白质代谢过程抑制肉仔鸡骨骼肌发育,降低生长性能。
3.2 DB对肉仔鸡血清生化指标的影响
血清生化成分的变化可以反映机体代谢和健康状况[16]。在家禽中,尿酸(UA)是蛋白质代谢的终产物,血清中UA 的水平可以作为反映体内蛋白质代谢的指标[17]。在本研究中,饲粮中添加DB 使肉仔鸡血清中UA 浓度明显上升,表明机体蛋白质分解增强。血清白蛋白(ALB)和总蛋白(TP)是反映机体蛋白合成能力的两个重要指标[18]。本研究发现,饲粮中添加DB 显著降低了血清TP 水平,这表明DB 能够减弱肉仔鸡蛋白质合成过程。以上结果表明,DB 可以通过抑制肉仔鸡体内蛋白质合成,促进蛋白质分解,这与胸肌重量降低相一致。谷草转氨酶(AST)和谷丙转氨酶(ALT)是参与机体蛋白质和氨基酸代谢的两种重要转氨酶,其活性可以反映肝脏健康状况。Dai等[19]研究表明家禽肝脏功能受损时血清AST 和ALT水平会显著升高。本试验发现,饲粮中添加20 mg/kg和100 mg/kg的DB使血清中ALT含量较对照组显著提高。这表明饲料中添加高剂量DB似乎会对肉仔鸡肝脏代谢造成不利影响,但具体机制仍需要进一步研究。
3.3 DB对肉仔鸡骨骼肌苦味受体基因表达的影响
苦味感知可以帮助动物从正常食物中分辨出潜在的有毒和有害物质[20]。动物的苦味受体除了在味蕾中表达,还广泛存在于体内其他组织中[3-4]。Kok等[21]研究发现,激活肥胖小鼠的肠道苦味受体能够调控激素分泌和胆汁酸代谢,从而减轻一些代谢疾病的相关症状。Hamdard等[7]研究表明,苦味受体基因的激活,能够诱导肉仔鸡心脏和肾脏的氧化损伤进程。以上研究表明,苦味受体在不同组织器官中可能存在不同生理功能。家禽中共有三种苦味受体,分别是gg-Tas2r1、ggTas2r2和ggTas2r7[22]。由于苦味受体数量较少,家禽是研究非味觉组织中苦味受体功能的一个适宜的模型[23]。本试验中,饲粮中添加100 mg/kg的DB,显著上调苦味受体基因ggTas2r7在肉仔鸡骨骼肌中的表达水平。磷脂酶Cβ2(PLCβ2)和3 型肌醇-1,4,5-三磷酸受体(IP3R3)是苦味受体的下游基因,能够参与肌肉代谢的过程[24]。与苦味受体变化一致的是,高剂量的DB 能显著提升PLCβ2 和IP3R3在肌肉中的表达水平。以上结果表明,DB 能够通过激活苦味受体,参与骨骼肌的代谢。在本试验中,饲粮中添加5 mg/kg 和10 mg/kg DB 并未对骨骼肌中苦味受体及下游基因的表达产生显著差异,但具有上升的趋势。这表明DB对于肉仔鸡苦味受体通路的激活可能具有剂量效应。相较于人类和啮齿动物的8 000个味蕾,而鸡只有200~300个[25]。同时,鸡味蕾的大小似乎小于人类,表明鸡的味觉灵敏度不如人类[8,26]。这也可能是低剂量DB对骨骼肌苦味受体通路基因表达水平未产生显著差异的原因。
3.4 DB对肉仔鸡肌肉发育和代谢相关基因表达的影响
肉仔鸡骨骼肌肥大与机体的蛋白质合成和分解代谢密切相关。当骨骼肌中蛋白质的合成速率高于分解速率,可促进肌纤维中粗、细肌丝的增加,使得骨骼肌肥大;反之,则导致骨骼肌萎缩[27]。雷帕霉素靶蛋白(m-TOR)信号通路和泛素-蛋白酶体通路途径是调节蛋白质合成与分解的主要信号通路[28-29]。本研究中,检测m-TOR 信号通路相关基因(INSR、m-TOR和AKT)的表达无显著差异,表明DB对骨骼肌蛋白质合成无显著影响。萎缩素1(Atrogin-1)和肌肉环状指基因1(MuRF-1)是编码泛素-蛋白连接酶的基因,他们的表达是激活泛素蛋白酶体系统的重要原因之一[30-31]。在本试验中,高剂量DB可以显著增加肌肉中Atrogin-1 和MuRF-1 基因的表达。Glass 等[32]研究发现,肌动蛋白、肌球蛋白重链、肌球蛋白轻链和肌球蛋白结合蛋白C等主要肌原纤维蛋白可以被MuRF-1分解。本试验结果表明,饲粮中添加DB 可以使骨骼肌中蛋白质的分解增强,与血清中尿酸水平升高的结果一致,这可能是导致肉仔鸡体增重和骨骼肌重量降低的原因。Cheled-shoval 等[6]认为苦味受体下游基因IP3R3表达的上调将导致Ca2+从IP3敏感的Ca2+储存中释放,从而增加基础胞质Ca2+浓度。当基础胞质Ca2+浓度增加时,叉头盒转录因子O 型(FOXO)去磷酸化增加[33]。FOXO 作为Atrogin-1 和MuRF-1 的转录因子,随着FOXO去磷酸化增加,将上调其下游基因Atrogin-1和MuRF-1 的表达[34-35]。以上结果表明,饲粮中添加DB可能通过激活的苦味受体和泛素-蛋白酶体通路参与骨骼肌发育的调节。
4 结论
本试验条件下,饲粮中添加DB 会降低肉鸡的生产性能。其可能的机制是通过激活骨骼肌中苦味受体和泛素-蛋白酶体通路途径促进蛋白质水解进程,进而减少骨骼肌中蛋白质沉积,抑制骨骼肌发育。本研究结果有助于更深入地揭示鸡苦味受体在骨骼肌发育中的潜在调控机制,为苦味添加剂在鸡饲料中的应用提供参考依据。
猜你喜欢
职工法律天地·上半月(2022年3期)2022-04-15 17:43:16
养殖与饲料(2021年5期)2021-11-30 20:05:15
现代畜牧科技(2021年10期)2021-11-19 08:42:20
老年博览·上半月(2019年8期)2019-10-21 08:49:38
兽医导刊(2019年20期)2019-08-19 10:39:18
中成药(2018年8期)2018-08-29 01:28:10
今日畜牧兽医(2018年9期)2018-02-13 09:05:24
儿童故事画报·智力大王(2017年2期)2017-06-05 13:59:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
