
番茄双链RNA绑定蛋白(SlDRB)基因家族鉴定及抗TYLCV防御反应分析
2022-03-11 06:56:32方远鹏岳宁波李云洲
核农学报 2022年2期
黄 鑫 方远鹏 岳宁波 张 龙 吴 丹 李云洲
(贵州大学农学院,贵州 贵阳 550025)
番茄(Solanumlycopersicom)为茄科番茄属园艺作物,含有丰富的番茄红素、维生素C、类胡萝卜素等营养元素[1]。目前,番茄病虫害严重限制了番茄的高产和优质生产,其中病毒病害影响严重且不易防控,威胁着番茄的大面积推广和种植[2]。为解决病毒侵染带来的生产限制,目前可采用的方法包括增强植物植株先天抗性[3]、抗病毒药剂[4]的开发以及病毒侵染介质的防控[5],其中最为有效和经济的抗病毒策略是增强植物植株先天抗性,即抗病毒育种。植物体的先天抗病毒策略主要有两种,即R基因介导的抗病毒途经与RNA干扰信号途经[6]。
RNA干扰(RNA interference,RNAi)是植物抗病毒的重要机制,参与RNAi机制的蛋白包括核酸内切酶DCL(Dicer-like)、RNA依赖的RNA聚合酶RDR(RNA-dependent RNA Polymerase)和AGO蛋白(Argonaute),DCL主要通过剪切双链RNA(double strand RNA,dsRNA)形成初级小干扰RNA(small interference RNA,siRNA),RDR可以将siRNA重新反转录成为新的dsRNA,新合成dsRNA再次经过DCL剪切形成次级siRNA,AGO蛋白与siRNA结合形成RNA诱导的沉默复合体(RNA induced silencing complex,RISC),通过碱基互补配对方式降解外源RNA或病毒[7]。在此过程中,产生的大量次要蛋白也可以有效促进反应的完成及信号增强。双链RNA结合蛋白(dsRNA-binding proteins, DRB或dsRBPs)是一种参与RNAi通路的重要蛋白[8],在动物[9]、微生物[10]、植物[11]中广泛存在,该蛋白能参与抗病毒粒子的形成、RNA的加工形成、病毒信号识别等过程[12]。
DRBs作为辅助因子,可同DCLs在促进小干扰RNA的产生直到RISC的组装中,始终促进RNAi的发生[13]。DRBs最初被认为具备两个双链RNA结构域(DRB4D1和DRB4D2,又称DSRM),典型的DRB蛋白包含约70个氨基酸组成的α1-β1-β2-β3-α2结构[14]。但是不同分支下的DRB蛋白存在明显差异,如结构域的分布位置以及数量等[15]。目前DRB蛋白在拟南芥(Arabidopsisthaliana)、水稻(Oryzasativa)等植物基因组的成员数大多为10个以内,并且对DCL的活性至关重要[15]。前人研究表明拟南芥中DRB4介导了番茄斑点枯萎病病毒(tomato spotted wilt virus, TSWV)过程中21 nt病毒siRNA的合成[13];另外芜菁花叶病毒(turnip yellow mosaic virus, TYMV)感染寄主后,会诱导寄主DRB4表达,从而抑制病毒外壳蛋白的积累[16]。DRB4蛋白参与芜菁皱纹病毒(turnip crinkle virus, TCV)的识别过程,使R基因介导的特异性免疫能正常识别病毒外壳蛋白(coat protein, CP),DRB4促进植株对TCV病毒的超敏反应从而形成稳定的复合物[17]。烟草DRB2参与马铃薯X病毒(potato virus X,PVX)侵染过程并引起病原体相关分子模式(pathogen associated molecular patterns, PAMPs)植物防御反应,从而产生免疫反应(PAMPs triggered immunity,PTI)诱导组织坏死[18]。
DRB基因家族参与RNAi过程,促进植物病毒的降解,在RNAi抗病毒过程中发挥重要作用。本研究利用生物信息学的方法,鉴定出番茄SlDRB基因家族,并对其性质和抗病毒防御反应进行分析,旨在为番茄DRB抗病毒研究奠定基础,同时为研究SlDRB基因家族在番茄RNAi抗病毒研究中的功能提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为矮番茄[19],源自西北农林科技大学园艺学院番茄种质创新实验室,TYLCV侵染性克隆来自中国农业大学植物保护学院周涛实验室。
1.2 序列获取和DRB基因家族的鉴定
拟南芥、番茄序列来自TAIR(https://www.arabidopsis.org/)及Sol Genomics Network(https://solgenomics.net/)数据库。采用拟南芥DRB基因(AtDRB)家族全部蛋白序列对番茄蛋白组进行鉴定,将合并去重获得的全部基因置于NCBI-CD数据库确认结构域(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi),保留其具备DSRM结构域的蛋白基因。基于番茄基因组注释文件获得SlDRB基因的基本信息。最后利用在线工具ProtParam(https://web.expasy.org/protparam/)和PSORT(http://psort.ims.u-tokyo.ac.jp/form.html)进行基础理化性质和亚细胞定位分析[20]。
1.3 番茄DRB蛋白质系统发生分析
根据DNAMAN软件对拟南芥与番茄的多序列比对结束,进行番茄DRB基因的命名。然后根据拟南芥、番茄DRB家族蛋白序列通过Clustal X2.1软件进行多重比对分析,基于比对结果,利用MEGA7.0软件采用相邻连接法(neighbor-joining,NJ)构建进化树,设置bootstrap为1000[21]。
1.4 保守基序的鉴定及结构分析
利用 MEME (http://meme.Nbcr.net/meme/intro.html)[22]在线平台进行番茄 DRB蛋白中保守基序(motif)分析,最大motif检索数值为15,E-value < 0.05。采用SOPMA2.0和Phyer2.0网站进行蛋白高级结构的预测,并用pyMOL软件[23]进行蛋白质三级结构的绘制(http://npsa-pbil.ibcp.fr/cgi-bin/npsa-automat.plpage=npsa-sopma.html、http://www.sbg.bio.ic.ac.uk/phyre2/)。
1.5 基因结构和染色体定位、组织表达分析
根据染色体注释信息中包含的DRB基因的结构注释,利用TBtools v1.068对DRB基因的基因结构和染色体定位进行绘制。并通过BAR ePlant(http://bar.utoronto.ca/eplant_tomato/)检查番茄DRB(SlDRB)基因在不同组织的表达特征[24-26]。
1.6 DRB基因表达分析
精选矮番茄种子,温汤浸种后,放置于人工气候箱进行催芽,3 d后种子露白播种于灭菌基质,置于人工气候室进行培养。选取生长一致的5片真叶的幼苗进行接种番茄黄叶曲叶病毒(TYLCV)侵染性克隆,对照接种空载体,接种方法参考文献[27]。取样时间分别为接种后0、7、14 d。采用 Trizol法提取总RNA,经DNaseI处理去除基因组DNA,2 μg RNA经TaKaRa公司(大连)Primescript RT Reagent Kit试剂盒反转录合成cDNA第一链,稀释后做模板。根据番茄数据库基因序列,利用Primer 3.0设计引物(表1),由生工生物工程(上海)股份有限公司合成。荧光实时定量PCR(quantitative real time PCR, qRT-PCR)反应试剂盒购自南京诺维赞生物科技有限公司SYBR Master mix(TaKaRa,大连),反应体系为20.0 μL:SYBR Premix Ex TaqTM Ⅱ(TliRNaseH Plus)(2×)10.0 μL,引物(10 μmol·L-1)各0.8 μL,DNA模板2.0 μL,无菌双蒸馏水6.4 μL。反应条件为:95℃预变性30 s;95℃变性5 s,60℃退火20 s,40个循环;融解曲线分析95℃ 0 s,65℃ 15 s,95℃ 0 s,在CFX96TM Real - time System荧光定量PCR仪(美国Bio-Rad公司)上进行。采用2-ΔΔCt方法计算基因相对表达量,每个基因的表达反应重复3次,以Actin作为内参基因。

表1 研究所用的PCR引物Table 1 The sequence of PCR primers in this study
2 结果与分析
2.1 番茄DRB基因家族鉴定及基本理化性质分析
基于拟南芥序列Blastp结果和结构域特征获取番茄基因组的SlDRB基因,共得到8个含有DRB结构域的番茄SlDRB蛋白编码基因,根据它们同拟南芥序列的多序列比对同源性的相似度进行命名,相似度过低的以基因所处位置从SlDRB6开始依次往后命名(表2)。所有SlDRB蛋白编码基因的长度不固定,范围从SlDRB7的378 bp到SlDRB3的1 662 bp。所编码蛋白的长度为125~553 aa,分子量为14.2~60.2 kDa。另外,8个基因等电点均处于5.28~9.57之间,碱性蛋白包括SlDRB2、SlDRB3、SlDRB5,酸性蛋白包括SlDRB1、SlDRB4、SlDRB6、SlDRB8,近中性蛋白(6.5 表2 DRB基因家族基本理化性质Table 2 The Basic physical and chemical properties of DRB protein family 基于注释文件中对SlDRB基因的定位信息,通过TBtools绘制SlDRB基因的染色体定位及基因结构图,进而综合分析番茄SlDRB基因的改变情况。染色体定位显示8个SlDRB基因分布在番茄6条染色体上。其中,1号染色体上含有的SlDRB基因最多,包含SlDRB4、SlDRB6、SlDRB7,2、3、4、5、11号染色体各分布1个SlDRB基因。另外,SlDRB1、SlDRB2、SlDRB3、SlDRB5、SlDRB8染色体分布位置具有共同的特征,即均处于染色体下端,SlDRB6处于染色体上端,不同的是SlDRB4和SlDRB7成簇分布于1号染色体的中部(表2、图1-C)。 根据AtDRB和SlDRB氨基酸序列建立的进化树,可将DRB分为3个分支,即AtDRB2/3/5和SlDRB2/3、SlDRB6/7、AtDRB1/4和SlDRB1/4/5/8三支。其中AtDRB2/3/5和SlDRB2/3分支、AtDRB1/4和SlDRB1/4/5/8分支均包含2~3个内含子,SlDRB6/7分支包含0~1个内含子。SlDRB6是唯一缺乏3′端UTR的DRB基因,SlDRB4、SlDRB5则缺乏5′端UTR(图1-A、B)。 图1 拟南芥、番茄DRB蛋白的进化树及编码基因的基因结构(A)、染色体定位关系(B)Fig.1 Phylogenetic tree of DRB protein in Arabidopsis and tomato and gene structure (A), chromosomal localization relationship (B) of coding gene 基于NCBI-CD网站预测结果,显示SlDRB蛋白可以分成4类,第一类包括5个成员:SlDRB1、SlDRB2、SlDRB3、SlDRB4、SlDRB5,C端具2个DSRM结构域的DRB蛋白基本特征;第二类仅有SlDRB6,包含3个DSRM结构域,1个DSRM结构域保留在C端,2个DSRM结构域处于近中央的位置;第三类仅有1个成员SlDRB7,在C端出现一个DSRM结构域;第四类只有SlDRB8,在N端出现一个DSRM结构域。但是第三类SlDRB7和第四类SlDRB8蛋白结构不完整,暗示SlDRB7、SlDRB8的功能与其他有差异。 利用MEME网站预测番茄DRB蛋白、番茄及拟南芥DRB蛋白全部保守基序。结果显示,番茄DRB蛋白发现4个高度保守基序,其中motif A1和motif A2、motif A3和motif A4分别代表DSRM结构域的保守结构(图2-A)。在2种物种的保守基序中共发现9个高度保守基序,除了几个与motif A1~A4相对应的motif B1~B5外,SlDRB2和SlDRB3蛋白还存在4个新的保守基序,即motif B6~B9(图2-A、B)。 图2 番茄DRB蛋白的保守基序(A)、番茄及拟南芥DRB蛋白的保守基序(B)、保守基序图标(C)分析Fig.2 Analysis of conserved motif of DRB protein in tomato (A), motif of DRB protein in tomato and arabidopsis (B), motif graph (C) 另外,在SlDRB和AtDRB蛋白序列中第1和第2个DSRM结构域维持了两段保守的基序,分别为‘K*LQ*Y*G*H*F*V’和‘A*A’基序、‘K*L*E*P*Y’和‘G*K*A*A’基序。在番茄内还存在P、G、A等氨基酸,这些氨基酸在8个DRB蛋白的DSRM结构域区域内保持稳定(图2-C)。其次在PSORT的预测中,8条SlDRB蛋白被定位为胞外、细胞质、细胞核3个场所,其中胞外的最多,共5个,分别为SlDRB1、SlDRB5、SlDRB6、SlDRB7、SlDRB8,其次SlDRB2、SlDRB3被定位于细胞质,SlDRB4蛋白被定位于细胞核(表3)。 表3 DRB家族蛋白二级结构及亚细胞定位分析Table 3 Analysis of the secondary structure and subcellular localization of DRB family proteins 利用SOPMA 2.0网站对SlDRB蛋白质的二级结构进行预测,结果表明大部分SlDRB蛋白质二级结构以无规则卷曲为主,如SlDRB2、SlDRB3、SlDRB4、SlDRB6、SlDRB7、SlDRB8,其次是α-螺旋和延伸链,三者占比分别为46.45%~65.46%、20.80%~36.11%、8.45%~22.4%。SlDRB1蛋白主要以延伸链为主,其次是α-螺旋和无规则卷曲;SlDRB5的无规则卷曲与α-螺旋区域均为43.92%,延伸链占14.14%(表3)。 通过Phyer 2.0构建8条SlDRB蛋白的三级结构,结果显示不同的SlDRB蛋白三级结构差距较大。大部分SlDRB蛋白具备丰富的α-螺旋区域和延伸链,构型差距大,成圈状、椅状、钳状等多种构型。但是SlDRB1、SlDRB5的结构与二级结构预测结果存在差距,以无规则卷曲为主(图3、表3)。 注:红色、黄色、绿色分别代表α-螺旋、延伸链、无规则卷曲。Note: The red, yellow, green represent α-helix, sheet, loop, respectively.图3 番茄DRB蛋白三级结构分析Fig.3 Tertiary structure analysis of tomato DRB protein 基于ePlant提供的番茄组织表达分析,得到8个SlDRB基因在10种不同阶段或组织中的基因表达谱。综合分析组织表达的卡通热图和普通热图,获得SlDRB家族的组织特异性表达图谱(图4)。分析显示,SlDRB5和SlDRB6在植株中整体表达较低,而其余6个SlDRB基因(SlDRB1~4、SlDRB7~8)在所有组织的各个阶段均有较高水平表达。SlDRB5在开放的花和叶片中表达水平最高,其次是果(3 cm直径)和花蕾,在根和幼果不表达;SlDRB6仅在花蕾中表达,在开放的花和果实中表达非常低。此外,8个SlDRB基因均在叶片组织中表达,大多数基因都在根、花、果组织中达到最高的表达水平。 图4 番茄DRB基因的组织表达分析Fig.4 Expression analysis of the DRB gene family in different tissues 通过组织表达的分析,初步了解到了番茄DRB基因的特异性。为进一步了解不同阶段的基因功能,筛选参与抗TYLCV防御反应的SlDRB蛋白,通过实时荧光定量检测TYLCV侵染番茄植株不同阶段SlDRB的表达情况。由图5可知,8个SlDRB基因在未处理时相对表达水平较低,TYLCV侵染后不同的SlDRB表达均有升高。其中,SlDRB7、SlDRB8在7 d和14 d时间表达持续升高,相对表达水平始终高于0 d。SlDRB2、SlDRB3在病毒侵染第7天时表达显著上调,病毒侵染第14天时表达下调,但表达水平与0 d相比仍然显著上调。SlDRB1、SlDRB4在初次显现症状时(7 d)表达量变化不显著,但是在14 d时表达水平显著上调。SlDRB5、SlDRB6在组织表达分析中整体的表达量偏低,在病毒侵染的7 d时表达水平显著升高,但14 d时表达水平恢复至初始水平。 图5 DRB基因家族接种TYLCV后不同时间段的表达分析Fig.5 Expression analysis of different time periods after the DRB gene family was vaccinated against TYLCV 病毒病是威胁蔬菜以及其他农作物安全生产的重要病害之一,由于病毒易突变以及传播介体昆虫难控制等因素,导致病毒病成为农业生产中最难防控的一类病害[28]。RNAi是植物抗病毒的重要机制,是植物抗病毒研究以及抗病育种的重要方向[29],同时也广泛应用于其他生物逆境,如真菌、昆虫等病虫害[30],并且可拓展延伸至基因编辑效率的研究[31]。而DRB蛋白作为RNAi路径中的又一重要元件,不仅可以参与病毒粒子的识别和结合,而且可以促进小RNA(sRNA)和RISC的形成[17,32-34]。另外,不同DRB蛋白存在功能分化和拮抗效应[35]。 本研究基于番茄基因组鉴定出8个编码DRB蛋白的基因家族成员,这些番茄DRB家族(SlDRBs)含有DSRM结构域,其数量和位置在染色体上并不固定。SlDRBs主要分布于染色体的两端,包括SlDRB1、SlDRB2、SlDRB3、SlDRB5、SlDRB8、SlDRB6,仅SlDRB4和SlDRB7成簇存在于1号染色体的中部。以前的研究中,DRB主要含两大类,即DRB1、DRB2-3-5(DRB2,DRB3-5);DRB4(DRB4A、DRB4B)、DRB6,且DRBs往往具备不定量和不同位置的DSRM结构域[14-15]。在拟南芥和番茄的DRB蛋白进化树中,主要分出3个支,即DRB2/3和AtDRB5分支、SlDRB6/7分支、DRB1/4和SlDRB5/8分支,不同的支内含子数量和蛋白结构分布有所差异。蛋白的亚细胞定位包括胞外、细胞质、细胞核,这些结果表明拟南芥与番茄SlDRB基因的相似度较低,虽然部分番茄DRB蛋白与拟南芥DRB蛋白归为一支,但大多难以明确分出单个类蛋白。因此,DRB蛋白的功能有待进一步研究分析。 多数基因家族研究中,不同的基因会产生组织特异性表达差异,且同类基因在不同的物种以及基因簇均会产生巨大的差异[36-38]。例如RNAi机制的主要蛋白AGO、DCL、RDR在不同组织器官中同样表现出组织特异性[39-40],本研究发现番茄RNAi机制又一重要蛋白DRB家族同样存在组织特异性表达。本试验结果显示6个SlDRB基因(SlDRB1~4、SlDRB7~8)为组成型表达,并均产生独特的组织表达差异。另外2个SlDRB基因(SlDRB5和SlDRB6)组织选择性极强,SlDRB5在根和幼果中表达非常低,SlDRB6在开放的花和果实中表达非常低,但所有的SlDRBs基因在叶组织的表达适中,在根、花、果组织中表达较高,由此可以推测SlDRB可能在根、花、果发育过程发挥作用。 DRB基因对于植物抗病毒过程十分重要,前人研究表明DRB2及DRB4蛋白可参与RNA病毒抗病毒防御[13,16-18]。本研究同样发现番茄DRB可参与抗DNA病毒TYLCV的防御反应。在拟南芥研究中同样存在着独特的表达特征,Qu等[41]在拟南芥研究中发现DRB4对于有效的抗病毒沉默是必需的。虽然番茄幼苗DRB基因均受TYLCV侵染后产生表达水平上调,但显症初期(7 d)和显症后期(14 d)产生了3类变化趋势,包括始终表达上调的SlDRB7、SlDRB8,先表达上调后下调的SlDRB2、SlDRB3,仅在7 d或14 d表达上调的SlDRB1、SlDRB4和SlDRB5、SlDRB6。这一现象指示了DRB基因对不同病程阶段的差异性表达,然而其分子机制、选择性上调的原因均有待进一步探索。 本研究通过生物信息学分析鉴定到番茄DRB基因家族中共有8个家族成员,分布于6条染色体上。根据氨基酸同源性DRB基因家族可分出3个分支,内含子数量和蛋白结构分布有所差异。亚细胞定位预测细胞质DRB蛋白有SlDRB2和SlDRB3,细胞核DRB蛋白有SlDRB4,其余均为胞外蛋白。不同DRB基因的组织表达均有不同,番茄DRB基因在叶组织的表达都相对适中,大多数基因都在根、花、果组织中取得最高的表达水平。8个DRB基因均参与TYLCV诱导的防御反应,其中SlDRB7、SlDRB8表达水平随着TYLCV侵染时间持续上调,可知番茄DRB基因家族不同抗病毒阶段的功能不同。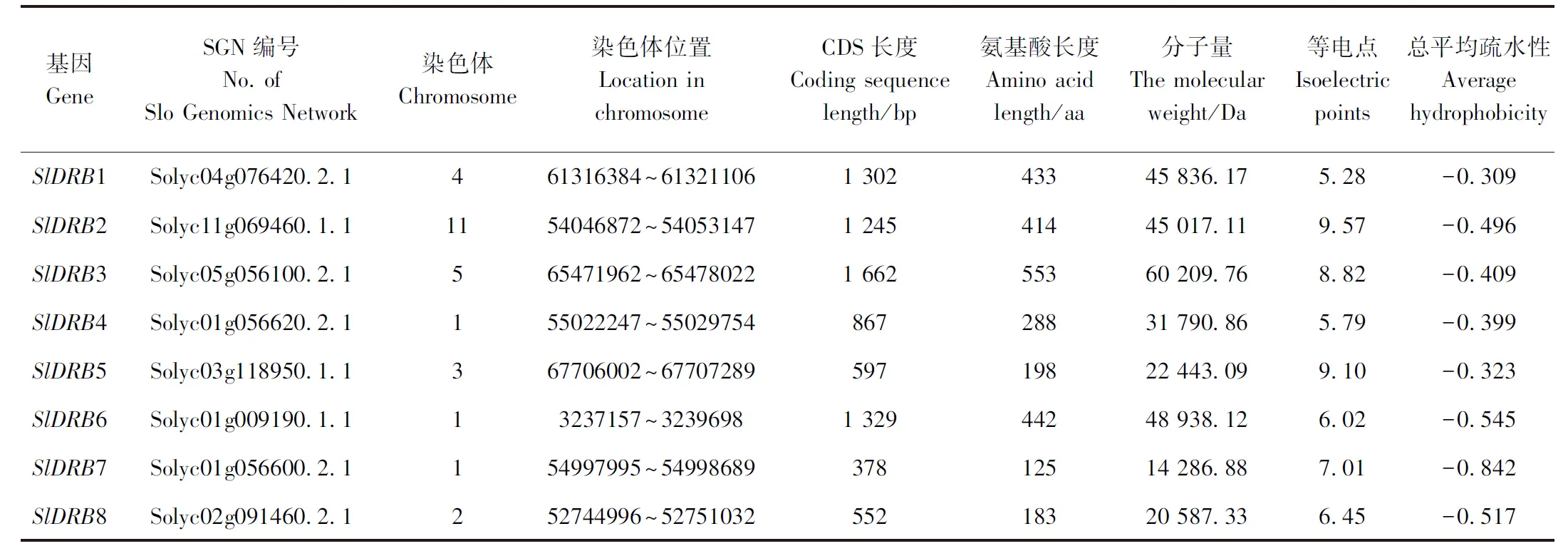
2.2 拟南芥、番茄DRB基因的结构、染色体定位分析

2.3 番茄DRB蛋白特性分析

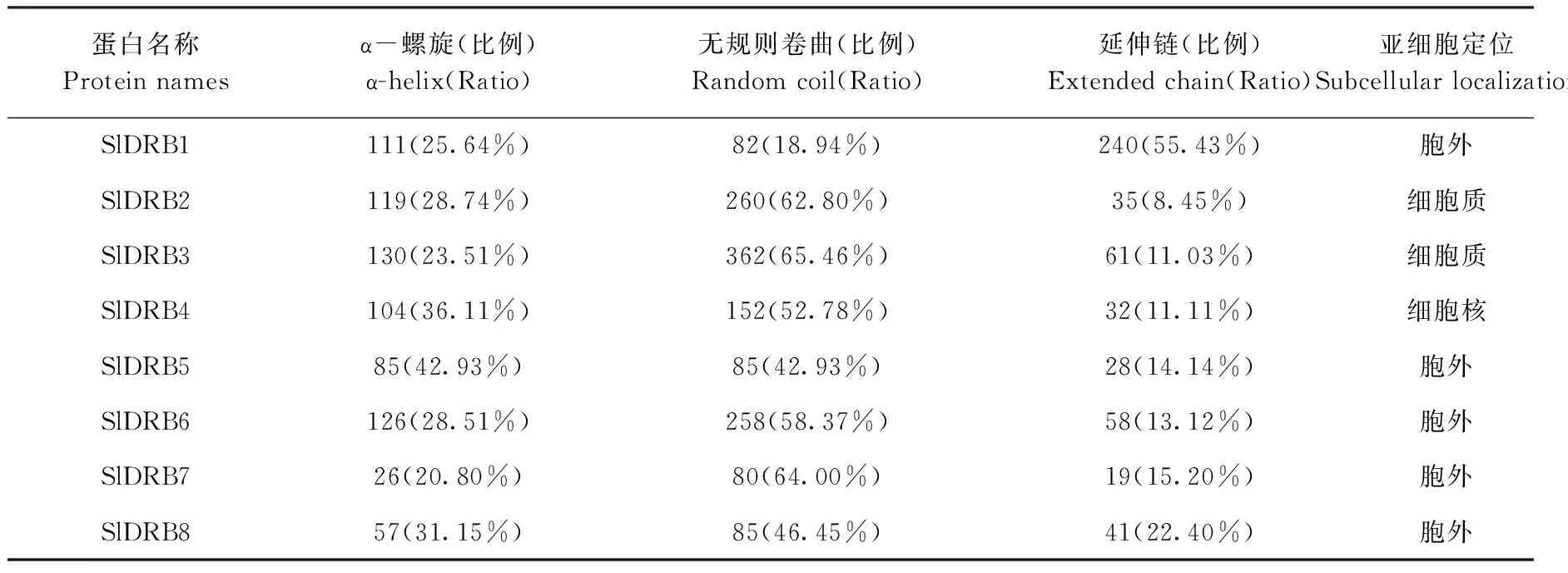
2.4 番茄DRB蛋白结构分析

2.5 番茄DRB基因组织表达分析

2.6 番茄DRB基因抗TYLCV侵染的表达分析
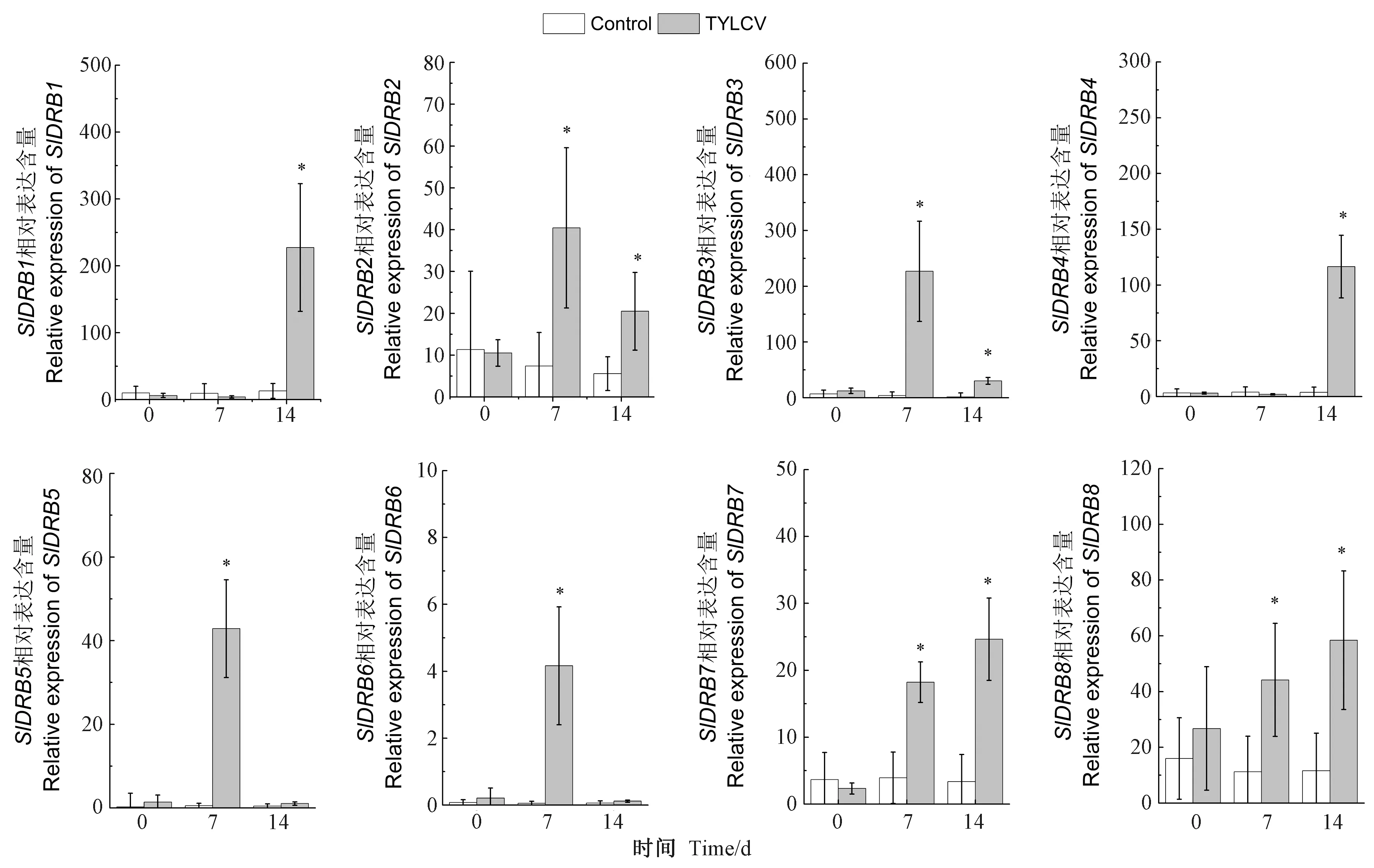
3 讨论
4 结论
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
肝博士(2022年3期)2022-06-30 02:48:52
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
肝博士(2021年1期)2021-03-29 02:32:14
肝博士(2020年4期)2020-09-24 09:21:26
疯狂英语·初中天地(2020年3期)2020-05-21 03:36:52
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
