
黔中北亚热带喀斯特次生林动态监测样地:物种组成与群落结构
2022-03-10 03:07:06张忠华刘立斌程安云吴洋洋倪健
生态学报 2022年2期
张忠华,胡 刚,刘立斌,程安云,胡 聪,吴洋洋,倪健,*
1 南宁师范大学环境与生命科学学院/北部湾环境演变与资源利用教育部重点实验室,南宁 530001 2 浙江师范大学化学与生命科学学院,金华 321004 3 中国科学院地球化学研究所环境地球化学国家重点实验室,贵阳 550081 4 中国科学院普定喀斯特生态系统观测研究站,普定 562100 5 贵州师范学院地理与资源学院,贵阳 550018
喀斯特地貌是世界上广泛分布的一种地质景观类型。我国喀斯特地貌区面积约344万km2,约占国土总面积的36%,占全球喀斯特面积的15.6%,西南地区有碳酸岩出露面积约51万km2,占该区域总面积的5.8%,其中贵州、云南和广西3个省份是西南喀斯特地貌最为典型的集中连片区域[1—2]。喀斯特生态系统具有土层浅薄、土被不连续、水土易于流失、树木不易定植等生态脆弱特性,加上西南地区持续的人为干扰和资源的不合理利用,导致喀斯特植被破坏严重,生态系统退化和石漠化等问题较为突出[3—6]。现存的西南喀斯特植被多为灌丛或灌草丛,而原生林则丧失殆尽,残存的森林常以遭受干扰后逐步恢复起来的次生林为主。喀斯特植被对维持生态系统结构和功能的稳定性极为重要,同时对探索岩溶地质背景下诸多生态过程与驱动机制具有重要科研价值[7]。我国西南喀斯特地区片段化分布的次生林是石漠化山地植被恢复的重要参考样本,对于检验生态恢复的理论框架和指导植被恢复的实践成效具有重要意义。
固定监测样地的建设为森林生物多样性定位研究提供了重要的平台[8—9]。目前为止,我国学者在温带、亚热带和热带气候区已建成了18个涵盖多种森林类型的大型动态监测样地和50多个1 hm2以上的辅助样地[10]。这些森林固定样地对于研究不同地区森林的群落结构与演替动态、物种空间分布格局及其群落多样性的维持机制起到了重要的推动作用,也为不同森林生态系统的结构、功能等比较研究提供了理想的平台[11]。喀斯特森林是独特的非地带性植被类型,为对该类森林进行定位研究,在广西弄岗和木论、贵州茂兰等地已建成有面积不等的固定监测样地,并在植物群落构建机制、动物的物种多样性等方面已取得了诸多可喜的成果[12—16]。以上喀斯特森林监测样地均为原生性较强的森林,而缺乏针对喀斯特次生林监测的固定样地,对次生演替过程中的诸多生态格局、过程与机制的研究还尚浅。对喀斯特次生林的研究可为区域性植被恢复与重建提供重要参考,因此亟待加强对喀斯特次生林的长期动态监测研究。
贵州是我国西南地区喀斯特地貌分布范围最广,面积比例最大的省份。位于贵州中部(简称黔中)的普定县是喀斯特高原石漠化的典型地区,同时也是开展石漠化防治研究的理想区域[6]。普定县境内的天龙山残存有该地区相对面积较大的次生常绿落叶阔叶混交林,是黔中典型的喀斯特森林植被类型之一,是开展喀斯特次生林定位研究的理想地点,对揭示该类型植被的结构、功能及其演变规律,提供喀斯特退化植被恢复与重建模式具有重要意义[17]。鉴于此,2012年中国科学院普定喀斯特生态系统观测研究站在天龙山南坡建立了一块面积2 hm2的固定森林动态监测样地,对天龙山次生常绿落叶阔叶混交林的生物多样性、生态系统结构和功能等进行长期监测研究,该样地的建立是对我国森林生物多样性监测网络的有益补充和完善。
针对北亚热带喀斯特次生林,在林分空间结构[18]、木本植物功能性状变异及其适应策略[19]、地上生物量结构[20—22]等方面已取得了一些研究成果。然而,对该次生林的物种组成、群落结构等基础群落信息尚不清晰。为此,本文以天龙山2 hm2森林监测样地的第一次本底调查数据为基础,分析了该样地次生林群落的物种组成、区系特征、径级结构及树木空间分布等基本特征,以期较为全面地展示黔中北亚热带喀斯特次生林的种群和群落结构特征,为今后开展喀斯特地区的植物群落构建机制、生物多样性与生态系统功能等研究提供参考资料。
1 材料与方法
1.1 研究区概况
天龙山位于贵州普定县马官镇下坝村,地理坐标26°14′40″ N,105°45′45″ E(图1),海拔约1550 m,该山处于典型的高原型峰丛洼地地貌类型之中,土壤类型为黑色石灰土,土层浅薄,地面岩石裸露明显,土被不连续,山体较为陡峭,平均坡度为31°。该地区属北亚热带季风湿润性气候,季风交替明显,全年气候温和,雨量较为充沛,日照少,无霜期长。年平均气温为15.1℃,极端最高温34.7℃,极端最低温-11.1℃,无霜期达289 d,年平均降水量1390 mm[18, 23]。
天龙山分布有典型的北亚热带喀斯特次生常绿落叶阔叶混交林,该林分平均高度7 m,郁闭度约0.75,林分成层现象较明显,可分为乔木层、亚乔木层、灌木层和草本层。林分物种组成和群落结构相对简单,乔木层常见种包括窄叶石栎(Lithocarpusconfinis)、化香树(Platycaryastrobilacea)、云南鼠刺(Iteayunnanensis)、安顺润楠(Machiluscavaleriei)、云贵鹅耳枥(Carpinuspubescens)、短萼海桐(Pittosporumbrevicalyx)、猫乳(Rhamnellafranguloides)等。灌木层以异叶鼠李(Rhamnusheterophylla)、倒卵叶旌节花(Stachyurusobovatus)、刺异叶花椒(Zanthoxylumovalifoliumvar.spinifolium)、薄叶鼠李(Rhamnusleptophylla)等为主;草本层有大披针薹草(Carexlanceolata)、矛叶荩草(Arthraxonlanceolatus)、阔叶山麦冬(Liriopeplatyphylla)、十字薹草(Carexcruciata)、野雉尾金粉蕨(Onychiumjaponicum)和对马耳蕨(Polystichumtsus-simense)等。藤本植物主要有香花鸡血藤(Millettiadielsiana)、藤黄檀(Dalbergiahancei)、钩刺雀梅藤(Sageretiahamosa)、长柄地锦(Parthenocissusfeddei)和常春藤(Hederanepalensisvar.sinensis)等。
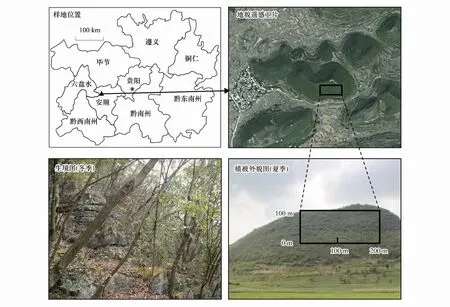
图1 天龙山样地的位置与植被外貌和生境照Fig.1 Location of the Tianlongshan plot and photos of vegetation phsiognomy and habitat
1.2 研究方法
1.2.1样地建立
除了广西弄岗、木论和贵州茂兰等自然保护区外,我国西南喀斯特地区较少有成片连续分布的喀斯特森林。黔中喀斯特山地强烈的人为干扰致使其山地以石漠化生境和灌草丛植被为主,封山育林后形成的小面积次生林在空间上呈现出明显的片段化和破碎化分布,因此,该区域不具备建设较大面积森林样地的可能性。2012年7—9月,中国科学院普定喀斯特生态系统观测研究站依据陆地生态系统生物观测规范[24]的要求,并参考美国Smithsonian热带研究所热带森林科学研究中心ForestGEO(Forest Global Earth Observatory,原CTFS)森林固定样地的建设标准[25],使用全站仪(Topcon GTS- 336,Topcon Positioning Systems,Tokyo,Japan)在天龙山南坡的次生常绿落叶阔叶混交林内建立了面积为2 hm2(东西长200 m,南北长100 m)的固定动态监测样地(图1)。该样地的海拔范围为1402—1512 m,平均坡度31°,岩石裸露率可达60%以上,土层厚度10—80cm,大部分地段土层厚度不超过50 cm。样地四周每间隔10 m用水泥桩作永久标记,用全站仪将整个样地划分为200个10 m × 10 m的调查单元,每个调查样方边角使用1 m高PVC管进行标记。对样地内所有胸径(Diameter at Breast Height,DBH)≥ 1 cm的木本植物进行挂牌、定位和鉴定,记录植物的编号、种名、胸径、树高、相对坐标和生长状况等。同时,也记录每个样方的海拔、坡度、坡向、岩石裸露率、地被物厚度等环境特征。
1.2.2数据分析
参照《贵州植物志》和Flora of China(www.efloras.org/)确定样地内被调查植物的科属种名称,编制天龙山样地植物名录并确定植物的生长型类别,包括乔木、小乔木、灌木以及藤本。按世界种子植物科和中国种子植物属的分布区类型划分标准统计样地内物种科、属的分布区类型[26—27]。计测植物物种和科的重要值,其方法为:
物种重要值(IV)=(相对频度+相对多度+相对优势度)/3,其中,相对频度= 某个物种在所有样方内出现的次数/所有种出现的总次数 × 100;相对多度= 某个物种的个体数/全部物种的个体数 ×100;相对优势度= 某个种的胸高断面积之和/所有种的胸高断面积之和 × 100[28]。
科的重要值(FIV)=(相对多样性+相对密度+相对优势度)/3,其中,相对多样性为某个科的树种数占总树种数的百分比;相对密度为某个科的树种密度占总树种密度的百分比;相对优势度为某个科的树种胸高断面积和占总树种胸高断面积和的百分比[29]。
种-面积曲线采用组合样方法绘制;种-多度曲线以多度为纵坐标,以树种多度从大到小排序为横坐标绘制;径级分布图采用的径级间隔大小为5 cm。
稀有种和偶见种的划分参照Hubbell和Foster[30]的定义,即稀有种为每公顷个体数≤1株的物种,偶见种为每公顷个体数1—10株的物种。
利用Excel 2010和R 3.4.4对数据进行整理分析和作图。
2 结果
2.1 物种组成
样地内共有DBH≥1 cm木本植物66种,其中乔木33种,灌木24种,藤本9种,隶属于34科55属。从科的分布水平来看,蔷薇科植物最为丰富,共有7属9种,豆科次之,有6属6种,樟科有3属4种,鼠李科有3属4种。从属的分布水平来看,花椒属的植物最多,有3种。另外,有64.7%的科及72.7%的属仅有一个物种分布。个体数和优势度最多的是壳斗科,分别占总个体数和优势度的24.40%和36.97%,其次是胡桃科,分别占总个体数和优势度的17.80%和25.88%。样地内物种数最多的10个科所含物种数占物种总数的50%,所含物种个体数占个体总数的89.18%(表1)。
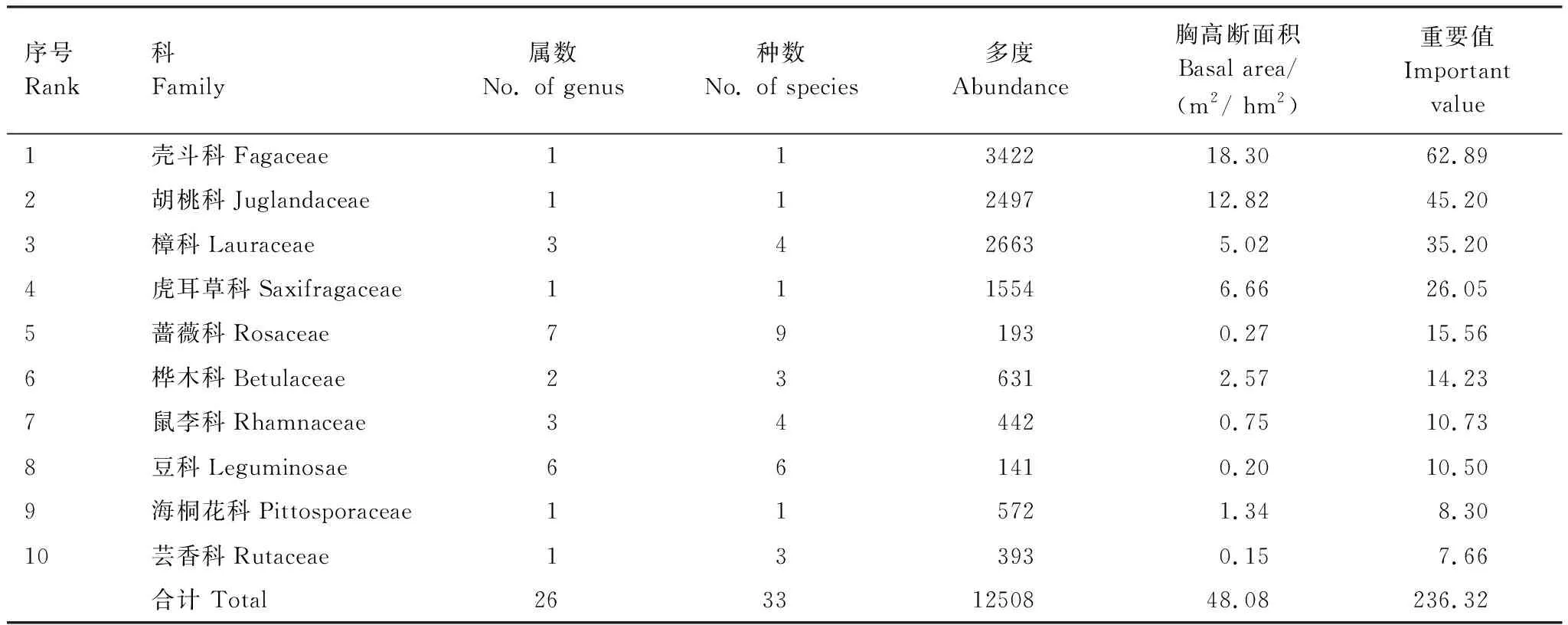
表1 天龙山样地木本植物中重要值前10位的科
2.2 区系成分
参照吴征镒[26—27]对我国植物科、属地理区系成分的划分,天龙山样地木本植物区系特征分析结果如表2所示。样地内木本植物的科分为7个分布型,4个亚型;属可分为13个分布型,2个亚型。热带性质的科占总科数的44.1%,其中泛热带分布型科数最多,占35.3%。温带性质的科占总科数的29.4%,其中北温带分布型科数最多,占23.5%。样地内热带区系的属占总属数的40.0%,其中泛热带分布的属比例最高,占16.4%;温带性质的属占总属数的52.7%,其中北温带分布的属数最多,占18.2%。世界分布科有9科,世界广布属有3属,中国特有属有1属。根据植物区系地理成分分析,表明研究样地植物区系的热带—亚热带性质,也反映出样地内植物区系由热带向温带过渡的特点。
2.3 多度格局
样地内独立个体数为14,025株(包括分枝、萌枝为17585株),单位面积个体数为7013株/ hm2。样地内个体数大于500的植物有7种,仅占总种数的10.61%,但个体数量占80.11%。个体数量最多的为窄叶石栎,占样地总个体数的24.4%,随后依次为化香树、安顺润楠、云南鼠刺,分别占总个体数的17.8%、14.6%和11.1%(表3)。样地内稀有种和偶见种分别有21种和18种,分别占总种数的31.82%和27.27%,分别占个体总数的0.19%和1.52%(图2)。
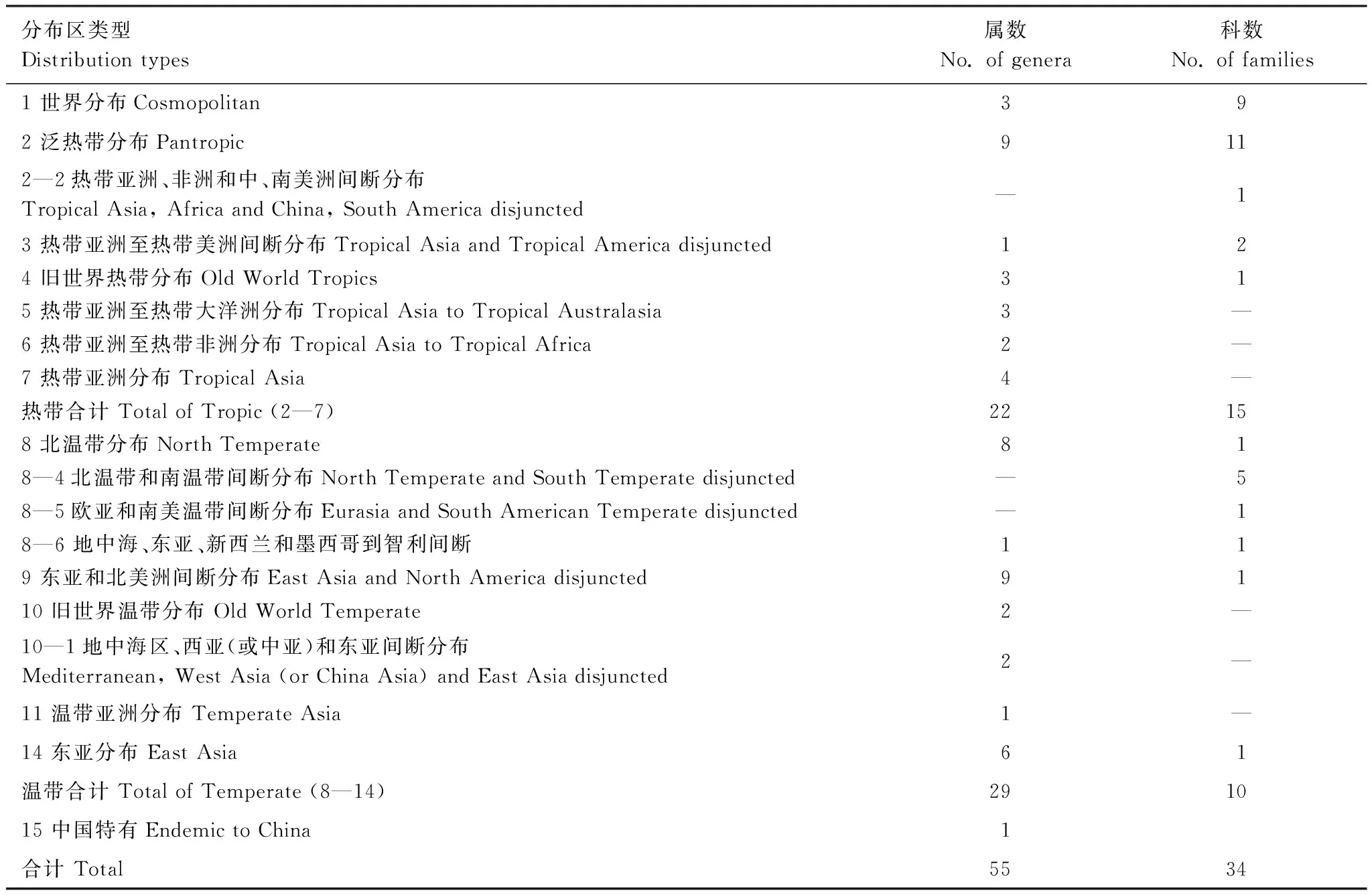
表2 天龙山样地木本植物的地理成分

表3 天龙山样地重要值≥1的优势物种组成
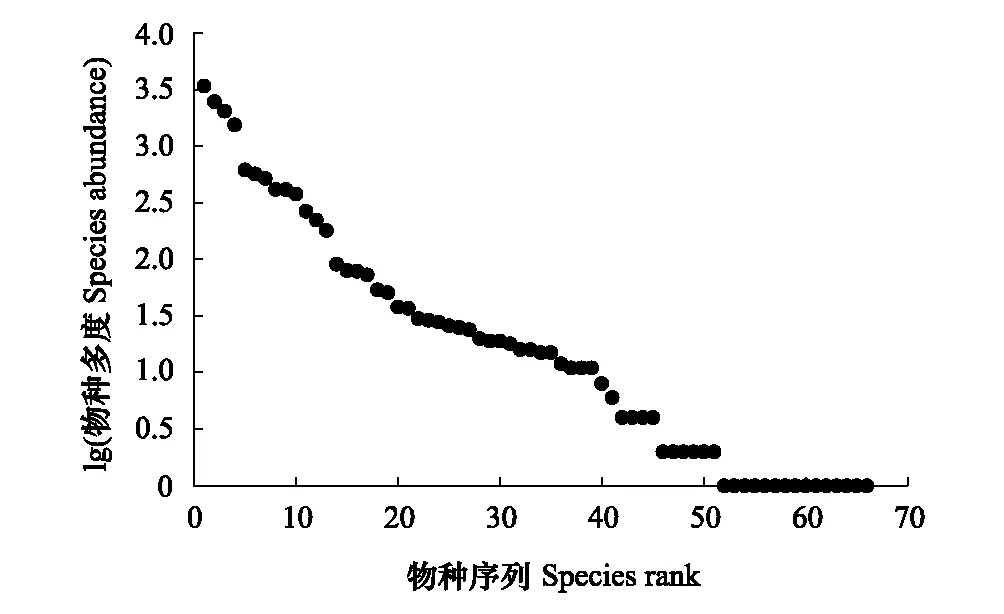
图2 天龙山样地的物种多度序列图 Fig.2 Sequence curve of species abundance in the Tianlongshan plot
2.4 优势种分析
样地内木本植物的优势种明显,样地内重要值≥1的物种有13种,占总物种数的19.70%,其重要值之和为88.81(表3)。窄叶石栎的重要值最大,有3422个个体,最大胸径达47.90cm,胸高断面积为9.15 m2/ hm2,占总胸高断面积的36.97%,是样地内胸高断面积最大的树种,也是乔木上层的优势种。其次分别为化香树(17.20%)、云南鼠刺(10.79%)、安顺润楠(10.09%)和云贵鹅耳枥(4.40%),这5个物种的重要值之和超过了65.00%,胸高断面积之和占样地总胸高断面积的89.21%,在群落中占据了绝对的优势。重要值第6—8位的物种分别是短萼海桐、香叶树和朴树(Celtissinensis),为样地亚乔木层的优势种;刺异叶花椒和倒卵叶旌节花是灌木层的优势种,胸径和胸高断面积较小,重要值位列第9和10位。
2.5 种-面积格局
天龙山样地物种的种-面积分析表明(图3),种-面积曲线的斜率在开始阶段随取样面积的增加而迅速降低。样地在取样面积较小的初始阶段物种数迅速上升,当取样面积达到0.42 hm2时,有45个种,约占样地总种数的70%,当取样面积达到1.3 hm2时,涵盖了样地90%的物种。种-面积关系表现出随着取样面积的增加,物种数先迅速增加后趋于稳定,后期随着面积增大,物种数变化幅度很小,这表明2 hm2的样地面积能够较客观体现出天龙山喀斯特森林群落的物种丰富度。
2.6 径级结构
样地内所有个体的平均胸径为5.12 cm,DBH≥1cm的个体的总胸高断面积为24.75 m2/ hm2。全部个体的径级分布呈现明显的倒“J”字型(图4),小径级的数量占较大优势,胸径1—5 cm的个体为8648株,占总株数的61.66%;5—10 cm的个体为3607株,占总株数25.72%;DBH≥10 cm的个体共1770株,占12.62%;DBH≥20 cm的共145株,占1.03%;DBH≥30 cm的个体数仅有9株,仅占总个体数的0.06%。倒“J”形的径级结构表明样地中有丰富的幼树储备,群落更新良好,呈稳定生长状态。
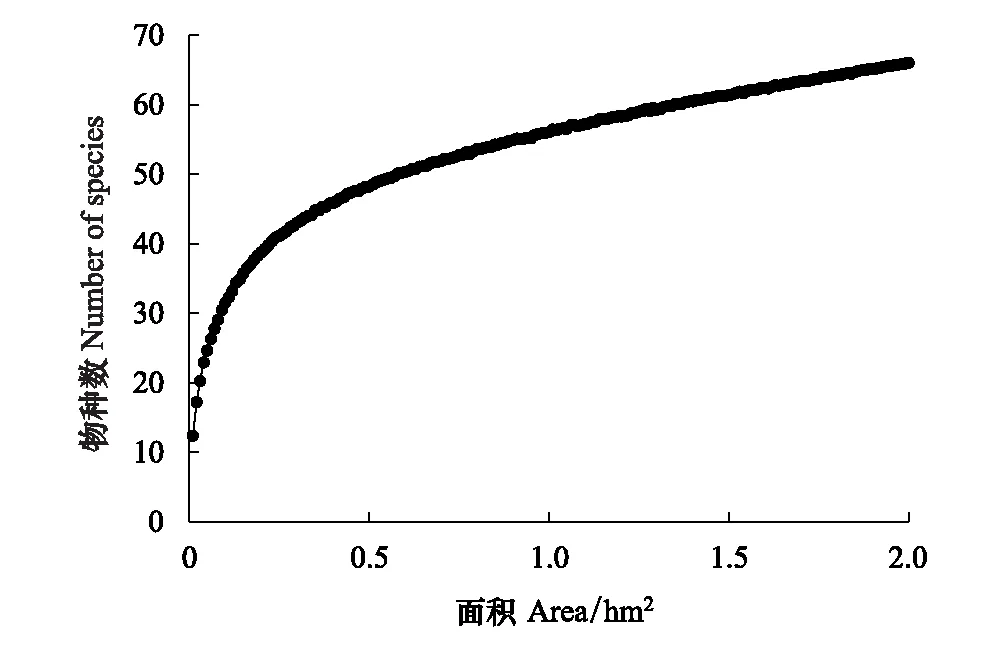
图3 天龙山森林动态监测样地的种-面积曲线Fig.3 Species-area curve in the Tianlongshan plot

图4 天龙山样地木本植物的径级分布图 Fig.4 Size-class distribution for woody species in the Tianlongshan plot
对样地内8个主要乔木树种的径级结构分析表明(图5),主要树种的径级结构可以归纳为3种类型:(1)倒“J”型分布,以1 cm ≤ DBH ≤ 5 cm的个体数最多,而后随着径级增大个体数逐渐下降,如窄叶石栎、安顺润楠、云贵鹅耳枥和短萼海桐;(2)“泊松”型分布,如化香树和云南鼠刺,DBH在5—10 cm的个体数最多,几乎没有大径级;(3)“L”型分布,它们多是亚冠层的优势物种,小径级的个体占绝对优势,而大径级的个体极少或无,如香叶树和朴树。

图5 天龙山样地8种优势树种的径级结构Fig.5 Size-class distribution of eight dominant tree species in the Tianlongshan plot
2.7 垂直结构
群落垂直成层明显,乔木层可分为2层,上层高达12 m,优势种比较明显,主要由窄叶石栎、化香树、云贵鹅耳枥等高大乔木组成,种类相对较少。亚乔木层高约7 m,以小乔木为主,常见种有安顺润楠、云南鼠刺、短萼海桐、珊瑚冬青、朴树、香叶树、猫乳等树种,数量较多,郁闭度达0.6。灌木层平均高约3 m,平均盖度为30%,主要有刺异叶花椒、倒卵叶旌节花、异叶鼠李、安坪十大功劳(Mahoniaeurybracteata)等组成,此外还有一些乔木的幼树。层间植物种类丰富,主要以粗大的木质藤本植物为主,包括藤黄檀、香花鸡血藤、钩刺雀梅藤、小果蔷薇(Rosacymosa)、柱果铁线莲(Clematisuncinata)、黑龙骨(Periplocaforrestii)等。样地内层次清晰,其中乔木上层和亚乔木层在垂直结构中占主要部分,灌木层和层间植物的稀有种和偶见种比例较高。
2.8 优势种的空间分布
从8种优势树种的空间分布看(图6),窄叶石栎的个体数明显高于其他树种,它在不同的海拔或坡位均有分布。8个主要优势树种均表现出不同程度的聚集分布特征,且与生境条件密切相关。如化香树在样地的低海拔区域分布较多,而短萼海桐更趋向于分布在样地的高海拔地段,云贵鹅耳枥主要聚集分布在样地的东北方向海拔偏高的区域,香叶树在样地的中部区域分布较多,云南鼠刺多分布于样地的西南方向。
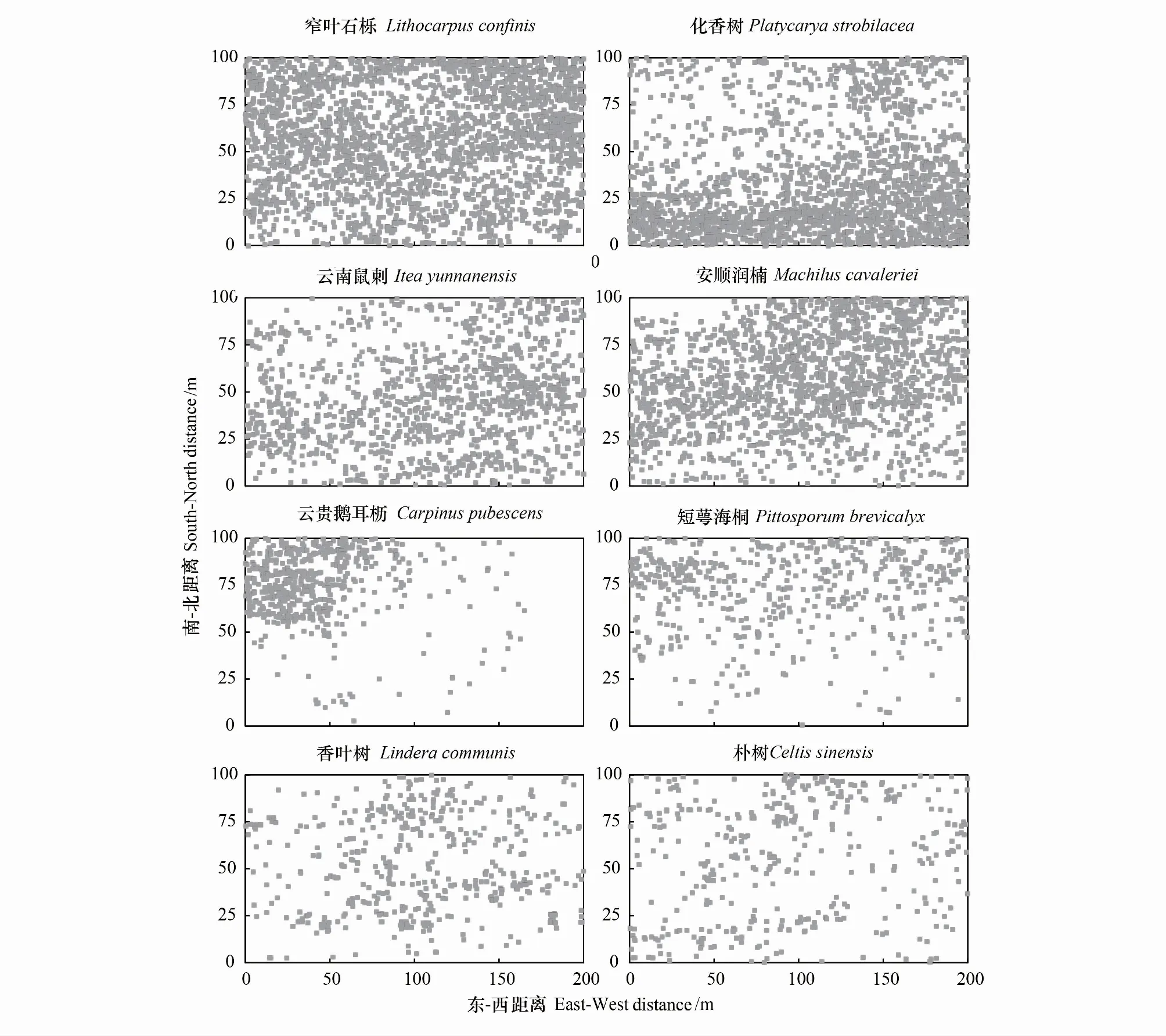
图6 天龙山样地8种优势树种的空间分布Fig.6 Spatial distribution maps of eight dominant tree species in the Tianlongshan plot
3 讨论
天龙山、弄岗、木论和茂兰等地均分布有我国西南地区典型的喀斯特森林,对比4个区域的植物群落结构特征能更全面地反映它们间的共性与差异。天龙山样地内DBH≥1 cm的木本植物合计有66种,隶属于34科55属,平均每公顷木本植物为55种。物种数量上远低于茂兰样地(199种/ hm2)、木论样地(124种/ hm2)和弄岗样地(121种/hm2),而单位面积的个体数上,天龙山样地却明显高于其他森林样地(表4)。天龙山为北亚热带喀斯特次生常绿落叶阔叶混交林,广西木论和贵州茂兰拥有中亚热带喀斯特常绿落叶阔叶混交林[12, 14],而广西弄岗分布有北热带喀斯特季节性雨林[11]。显然,天龙山木本植物多样性较低与样地所处纬度和海拔高度所形成的水热条件差异有密切关系,符合物种丰富度随纬度和海拔的升高而降低这一普遍规律。苏宗明和李先琨[31]认为非地带性的喀斯特植被具有自身起源与分布特点,同时带有植被地带性分布规律的烙印,我们的研究结果印证了这一观点。弄岗、木论和茂兰在成立自然保护区前后均长期处于良好的自然封育状态,因此除受水热条件影响外,黔中喀斯特地区高强度的人为干扰使得森林更具有次生性质,这也是天龙山物种丰富度较低而幼树个体数较高的重要原因之一。从物种科属组成来看,天龙山样地蔷薇科、豆科和樟科物种比较丰富,这点与弄岗北热带喀斯特森林样地树种以大戟科、马鞭草科和梧桐科为主明显不同[11],而与以樟科、蔷薇科、茜草科物种最丰富的茂兰样地具有一定的相似性[32],这体现了气候和地形地貌等生态要素在系统发育层面上过滤局域性种库,环境筛选作用决定了物种组成在科、属或种层面上的相似度。
天龙山样地稀有种占比为31.82%,其比例稍低于木论和弄岗样地(表4)。影响稀有种比例的原因可能有物种本身的种群特征与分布特性、生境异质性、森林类型镶嵌、干扰、区系的交汇和地形限制等[28, 33—34]。天龙山样地的纬度和海拔高度高于弄岗和木论样地,物种数较少(66种)且优势种明显,并且取样面积相对较小(仅2 hm2),这些均可能是样地所包含稀有物种的比例相对较少的原因。此外,天龙山样地稀有种比例相对于亚热带非喀斯特森林样地也少得多[28, 34—35],这可能与喀斯特地貌与微地形以及物种对喀斯特生境的适应性有关。喀斯特植物往往具备耐旱、耐钙、耐碱等共同特性[36],区域种库中的物种往往难以适应喀斯特这一特殊生境,同时黔中破碎化的生境和高度异质的喀斯特地形容易阻断种子长距离扩散,因此可能造成新物种的种源扩散受限[37]。此外,人类活动在很大程度上决定了物种的分布,特别是对生境要求更为“敏感”的狭域种和稀有种[38—39],因此,强烈人为干扰下的黔中喀斯特地区片段化森林中的稀有种相对于常见种可能更易于遭受负面影响。总的来看,天龙山样地稀有种的形成可能由气候与地形、生物地理过程和人为干扰等多维度因素共同决定的。上述推论仍需要进一步检验。
天龙山样地木本植物区系地理成分中,热带和温带分布的科分别占总科数的44.1%和29.4%。从物种属的分布区类型看,天龙山样地有55个属,热带成分占40.0%,温带成分占52.7%,温带成分大于热带成分,反映出样地内植物区系由热带向温带过渡的特点。从不同喀斯特森林监测样地内属的分布比例来看,茂兰样地的热带成分占67.1%,温带成分占29.3%[32];木论样地的热带成分占70.8%,温带成分占24.3%[14];弄岗样地的热带成分达88.39%[11]。这种区系成分有规律的变化,表明区域尺度上的植物区系组成与其所在的地理纬度和海拔有密切关系。天龙山样地所处的纬度和海拔较高,热带性质属的比例逐步减少,温带性质属的比例逐步增加,因此,喀斯特植被在区系成分变化上具有地带性植被的分布规律。
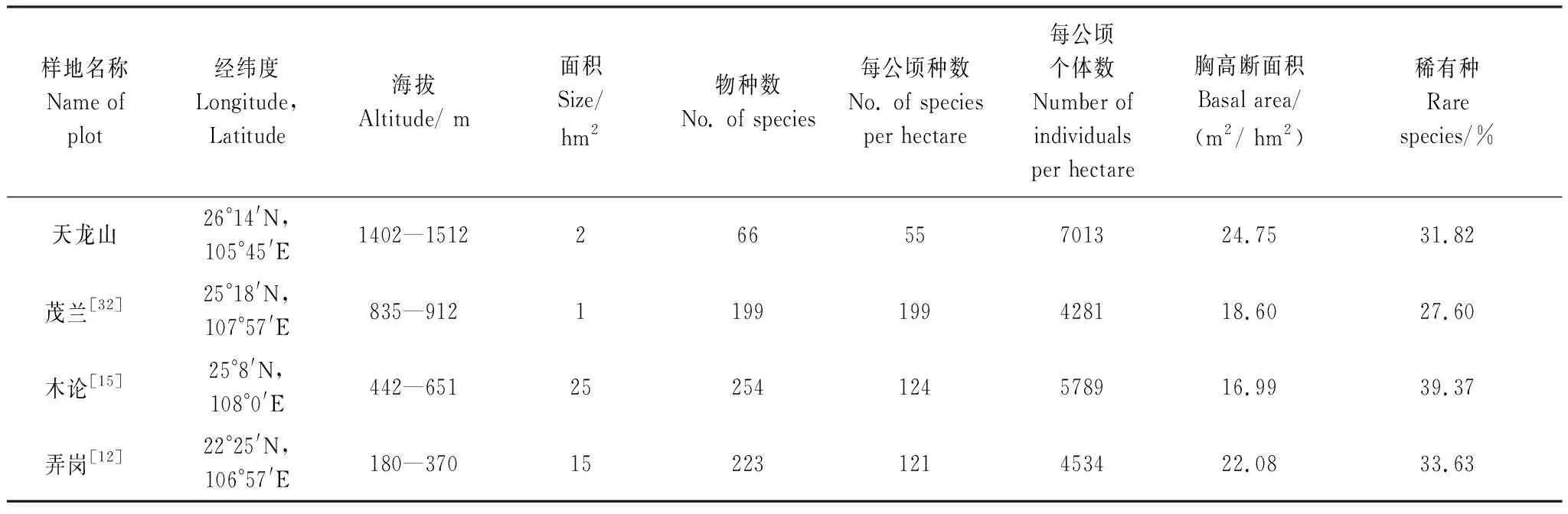
表4 天龙山样地与其他3个喀斯特森林动态样地特征的比较
天龙山样地优势种较明显,在所有物种中,窄叶石栎的重要值最高(23.15),其个体数和胸高断面积也最高(3422,9.15 m2/ hm2),其次是化香树(2497,6.41 m2/ hm2)、云南鼠刺(1554,3.33 m2/ hm2)和安顺润楠(2052,1.91m2/ hm2),这些物种均为乔木层树种,在样地内占绝对优势,属于该群落的优势种,这一群落结构特征与弄岗和茂兰森林群落不同[11, 32]。弄岗的季雨林和茂兰的常绿落叶阔叶混交林均无明显的优势种,多个优势种共存是这两个森林类型物种组成的重要特点之一。纬度和海拔决定的水热条件差异是驱动喀斯特森林优势种格局变化的主要原因。此外,天龙山样地为人为干扰破坏后形成的次生林,物种组成和群落结构相对简单,处于主林层的物种数量有限且个体数量较多,形成了明显的优势种,而处于长期封育的弄岗和茂兰喀斯特森林,同种以及近缘种间资源生态位维度的相似性以及长期累积的“天敌”—植食性昆虫和土壤病原菌等可能加剧了同种以及近缘种间排斥作用[40—43],因而长期封育的喀斯特森林具有更为复杂的群落结构和物种多样性。上述推论仍需要开展长期的定位观测和控制实验来加以验证。
径级结构是植物群落稳定性和生长发育状况的重要指标[28]。样地内所有个体径级结构呈倒“J”型(图4),表明群落整体更新良好。样地内DBH≥1 cm的树木平均DBH为5.12 cm,并且样地的个体数量较高,单位面积个体数为7013株/ hm2,高于弄岗样地、木论样地和茂兰样地(表4),这意味着该区域森林存在较多中小径级的植株,大径级的植株较少,反映了天龙山样地次生常绿落叶阔叶混交林个体密度较高这一特性。8个优势种的径级结构呈现倒“J”型、“泊松”型和“L”型分布,表明优势种均具有较为丰富的幼树补充,更新良好。总体来看,天龙山样地次生林优势种更新和林分发育良好。
从优势物种空间格局看,8个优势树种空间分布均具有一定的聚集性,且表现出生境关联性。影响森林物种空间分布格局的因素较多,如物种的环境适应性、生境异质性、种内与种间的相互竞争、物种生活史策略等[44—47]。弄岗、木论和茂兰样地中树种的分布格局均以聚集分布为主,生境异质性是影响其物种空间分布的重要因素[11—12, 14]。因此,生境异质性也可能是造成天龙山样地主要优势树种聚集格局的重要原因之一。由于岩石裸露度高、地形起伏较大,微地形组合复杂,土壤分布极度不均,光照、土壤水分和养分等生境特征的不均匀性、不连续性和复杂多样组合,致使同一地段、坡位、坡向的小生境,其生境特征也可能存在十分明显的差异,这样复杂的生境变化可能会导致物种分布上的专化。天龙山样地优势树种的聚集分布体现了物种有喜好的生境,如化香树和云贵鹅耳枥为喜钙、耐旱、耐贫瘠的树种,多分布在岩石裸露较高和坡度较大的生境生长,短萼海桐更倾向于分布在样地高海拔区域岩石裸露相对较少和缓坡的生境。
4 展望
本研究分析了黔中天龙山2 hm2喀斯特次生林样地中木本植物群落的物种组成、区系特征、径级结构和主要优势种的空间分布格局,同时推测了区系分布、物种组成和空间分布形成的潜在机制,研究结果对掌握北亚热带喀斯特次生林的构建过程、对比分析不同区域喀斯特森林结构与动态以及指导植被恢复与重建具有较为重要价值。目前,对黔中北亚热带喀斯特次生林的研究仍然较少,对其诸多的生态格局与过程的理解仍然不清,在北亚热带喀斯特植物多样性的形成及其维持机制、进化过程与群落生态过程及其之间的关系、地上与地下生态过程的耦联关系、喀斯特植被对气候变化的响应与适应等方面亟待深入研究[7, 17]。天龙山次生林动态监测样地的建立为上述研究工作的开展奠定了基础,为区域性植被恢复与重建提供了可参照的模板,是北亚热带喀斯特森林生物多样性重要的监测和研究平台。
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
阅读(低年级)(2020年8期)2020-11-06 06:23:19
乡村地理(2019年2期)2019-11-16 08:49:32
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
西北植物学报(2015年9期)2015-07-05 07:03:42
