
不同植被类型对不同土层黑土土壤微生物群落功能多样性的影响
2022-03-10 01:06邹狮严君陈旭陆欣春高瑞敏
土壤与作物 2022年1期
邹狮,严君,陈旭,陆欣春,高瑞敏,2
(1.中国科学院 东北地理与农业生态研究所,黑龙江 哈尔滨 150081;2.中国科学院大学,北京 100049)
0 引 言
土壤微生物在土壤生态系统中承担着重要角色,参与养分循环、转化和有机质分解等过程,对生态系统功能起着至关重要的作用[1-2]。土壤微生物群落功能多样性可以反映其对环境的响应和适应,并且能够反映出土壤中微生物群落生态系统功能的演化和土壤肥力的变化[3]。因此,研究土壤微生物的群落功能多样性对评估不同生态系统的稳定性非常必要[4]。
土壤微生物功能多样性是土壤中整体微生物多样性的一个方面,主要通过土壤微生物对碳源利用的情况对其进行衡量和评价[5],Biolog微平板法是目前评估土壤微生物功能多样性的一种简单有效的方法[6]。虽然国内外在利用Biolog方法研究土壤微生物对碳代谢上存在分歧,但它已被证明是区分不同环境土壤微生物功能多样性和群落干扰的有效方法[7-8]。土壤微生物群落功能多样性,受环境(非生物和生物)因素和人为管理措施的影响[9-10]。文东新等[11]对乔木、灌木和草本植被条件下土壤微生物群落功能多样性的研究表明土壤有机碳是影响土壤微生物代谢活性及功能多样性的重要因素[12]。Bertuni等[13]通过对人为扰动程度不同的南洋杉森林土壤微生物功能多样性的研究表明,与扰动和重新造林的南洋杉森林土壤微生物群落相比,原生南洋杉森林对土壤微生物群落的组成和功能影响更大。苜蓿和玉米种植条件下土壤微生物对糖类和氨基酸类碳源的利用能力显著不同[14]。此外,植被的多样性[15]、凋落物的数量[16]及种植密度[17]等因素亦会对土壤微生物的功能多样性产生影响。现有的研究大多是0~20 cm土层土壤微生物群落功能多样性的变化,而亚表层土壤中仍有大量的土壤微生物存在,可能含有一些特殊的针对其环境的微生物,从根本上区别于表层土壤微生物群落[18],推测其土壤微生物功能多样性亦与表层存在差异。因此本研究选用黑土区母质发育相同的土壤上长期无植被和有植被的(自然恢复、农作物及人工林)处理,通过对表层0~15 cm和亚表层15~35 cm土层中土壤微生物对碳源的利用情况,探索不同植被类型对不同土层微生物群落功能多样性的影响,揭示土壤微生物群落功能多样性与植被类型、土层深度之间的联系。
1 材料与方法
1.1 研究地区概况
研究区设置在中国科学院海伦农业生态试验站(47°26′N,126°38′E),该区位于黑土区中部的海伦市,平均海拔234 m,年均气温1.5 ℃,无霜期120~130 d,年均有效积温2 400 ℃,土壤为第四纪黄土状亚黏土发育而成的中厚黑土。
1.2 试验设计及样品采集
试验区大小为500 m×150 m,土地平坦,土壤基础肥力相对均匀。1985年在试验区设立4种不同植被类型处理,分别为:(1)自然恢复为草原化草甸植被(GL),植被类型丰富,有羊草(Leymuschinensis(Trin.)Tzvel.)、苔草(Carexspp.)、木贼(EquisetumhyemaleL.)和狗尾草(Setariaviridis(L.)Beauv.)等植被分布,面积104 m×16 m,无任何肥料投入,无耕作;(2)农作物(AL):当季作物为大豆,一年一熟小麦-玉米-大豆轮作种植,不施用肥料采用传统耕作方式,在作物生长季耙松4~6次,秋季作物收割后,移除作物地上部分,面积120 m×5.6 m;(3)人工落叶松林(FL):林分密度为2 100株·hm-2,株高约4.5 m,面积485 m×57 m;(4)无植被(BL):每年定期将地上部分铲除,始终保持无植被,面积30 m×16 m。由于每个样区的面积够大,所以将每个处理平均划分成三个小区作为重复。
2020年7月17日按5点法取样,在每个处理的样区内随机选取5个点分别采集0~15 cm和15~35 cm土壤样品,均匀混合后,收集在无菌自封袋中并带回实验室,去除肉眼可见的动植物残体和砾石,用2 mm筛子过筛后冷藏在4 ℃冰箱内备用,7 d内完成所指标的测定。
1.3 测定项目及方法
采用Biolog-Eco板,每个板内含96个小孔,设3次重复,每重复含1个空白对照和31种碳源。参照Qian[19]方法培养,具体步骤如下:称取相当于5 g烘干土的鲜土加入到装有45 mL灭菌的生理盐水(0.85%)的100 mL三角瓶中,在摇床上振荡约30 min,转速180 r·min-1,静置片刻。吸取一定量的上清液,将0.85%的灭菌生理盐水稀释至10-3倍,用八通道移液器吸取150 μL稀释后的上清液加入Biolog-Eco微孔板中,放在恒温培养箱中28 ℃恒温避光培养至168 h。每隔24 h用Biolog MicrostationTM酶标仪测定590 nm和750 nm处的吸光值。土壤微生物功能多样性各项指数参照Classen等[20]方法计算。
AWCD=∑(Ci-R)/n
式中:Ci为第i孔在590 nm和750 nm下的吸光值之差;R为对照孔的吸光值;n为Biolog-Eco微平板的碳源种类数(n=31)。利用各样品培养96 h的数据,计算其微生物功能多样性指数:
Shannon物种丰富度指数(H):H=-∑Pi/lnPi


Richness指数(S):S=碳源代谢孔的总数目(微孔的光密度值≥0.25,则认为该孔碳源被利用,即为碳源代谢孔)。
式中:Pi为第i孔的相对吸光度值与整个平板相对吸光度值总和的比率;ni为第i孔的相对吸光度值。
1.4 数据分析
采用SPSS 17.0软件对数据进行处理分析,对不同植被类型不同土层土壤微生物群落平均颜色变化率、功能多样性指数及碳源利用特征进行单因素方差分析,采用Duncan多重比较法在0.05水平上进行显著性差异检验。使用CANOCO 5.0软件对培养96 h 的土壤微生物进行主成分分析。
2 结果与分析
2.1 不同植被类型对不同土层土壤微生物群落平均颜色变化率(AWCD)的影响
不同植被类型下不同土层土壤微生物群落的AWCD有差异(图1),随着培养时间的延长均呈逐渐增长趋势。不同处理表层0~15 cm土壤微生物在培养的第120 h时趋于稳定,而15~35 cm土层在第168 h趋于稳定,且表层0~15 cm的AWCD值均显著高于对应处理15~35 cm土层。在整个培养时间内两个土层的AWCD值变化均表现为GL>AL>FL>BL。在培养的第96 h时,GL处理在0~15 cm土层的AWCD值比AL、FL和BL处理分别增加了4.2%、8.3%和131%,在15~35 cm土层中比AL、FL和BL分别增加了8.2%、15.2%和216%。对于同一处理,不同土层的AWCD值差异也很明显。96 h时,GL、AL、FL和BL处理在0~15 cm土层的AWCD值比对应处理15~35 cm土层分别增加了15.5%、19.9%、22.9%和57.7%。
2.2 不同植被类型对不同土层土壤微生物功能多样性的影响
利用培养第96小时的AWCD值(图1),计算土壤微生物的Shannon指数、Simpson′s指数、McIntosh指数和Richness指数,以反映土壤微生物碳源代谢功能多样性差异。不同植被类型对不同土层微生物群落的Shannon指数、Simpson′s指数、McIntosh指数和Richness指数的影响不同(表1)。GL处理在0~15 cm和15~35 cm土层4个多样性指数均最高,且0~15 cm土层土壤微生物群落的4个指数均高于15~35 cm土层,其中McIntosh指数和Richness指数之间差异显著(P<0.05),而Shannon指数和Simpson′s指数之间差异不显著。BL处理在0~15 cm和15~35 cm土层4个指数均最低,且BL处理0~15 cm的4个指数显著高于15~35 cm(P<0.05)。15~35 cm土层中AL处理的4个指数稍高于FL处理对应的指数,但无显著差异。

注:BL:无植被;AL:农作物;GL:自然恢复;FL:人工林。下同。Note:BL means no vegetation cover;AL means crops;GL means natural restoration of grass;FL means forests.The same is as below.图1 不同植被类型在0~15 cm(A)和15~35 cm(B)土层中土壤微生物群落平均颜色变化率(AWCD)Fig.1 Average Well Color Development (AWCD)in soil layers of 0~15 cm (A)and 15~35 cm (B)under different vegetation types
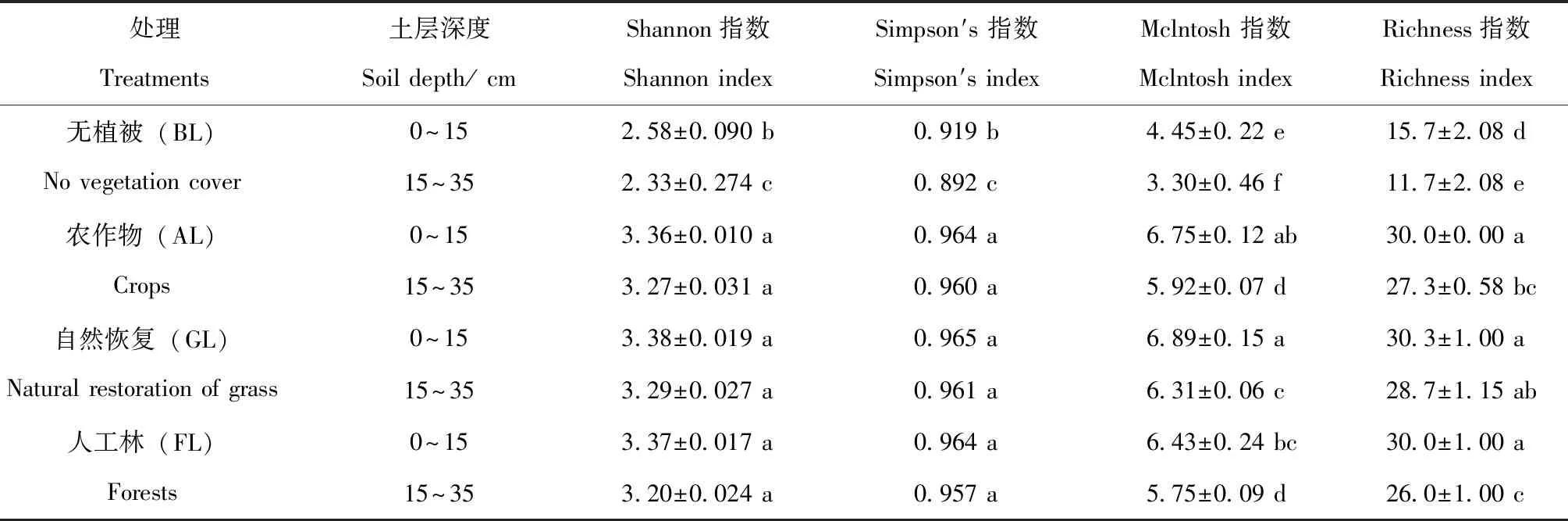
表1 不同植被类型在0~15 cm和15~35 cm土层中土壤微生物群落功能多样性指数的变化(96 h)Table 1 Variation of functional diversity index of soil microbial community in 0~15 cm and 15~35 cm soil layers of different vegetation types(96 h)
2.3 不同植被类型对不同土层土壤微生物群落对碳源的利用特征
不同植被类型土壤微生物群落对碳源总的利用能力表现为GL>AL>FL>BL,且对六种碳源的利用亦存在差异(图2)。在表层0~15 cm土层BL处理土壤微生物对氨基酸类碳源的利用高于对羧酸类、糖类及其它碳源,而AL和FL处理对羧酸类碳源利用最高,GL处理对氨基酸类利用最高。在15~35 cm土层不同植被类型土壤微生物对六类碳源总的利用能力整体低于0~15 cm土层对应的处理,且对碳源的利用种类与表层存在差异。BL处理对酚酸类利用最高,且显著高于该处理0~15 cm对酚酸类碳源的利用。AL和FL处理对氨基酸类利用最高,GL处理对羧酸类利用的最高。不同植被类型在不同土层碳源的利用能力存在差异,同时在同一土层中不同植被类型土壤微生物对六种碳源的利用也存在差异。

图2 不同植被类型下不同土层土壤微生物对六类碳源的利用情况Fig.2 Utilization of six kinds of carbon sources by soil microorganisms in different soil layers of different vegetation types
2.4 不同植被类型对不同土层土壤微生物对碳源的主成分对比分析
利用培养96 h标准化后的OD值,对BL、GL、AL和FL处理下土壤微生物对碳源的利用进行主成分分析,提取2个主成分,累计贡献率达到72.30%,其中第一主成分(PCA1)方差贡献率为59.22%,第二主成分(PCA2)方差贡献率为13.08%(图3)。不同植被类型土壤微生物对碳源的利用在PC轴上出现了明显的分布差异。表层0~15 cm中GL、AL和FL处理分布距离较近,土壤微生物利用的碳源相似,而BL处理与之显著不同。在15~35 cm土层不同植被类型聚成三组,GL和AL处理聚在一起,BL、FL单独聚类,表明GL和AL的土壤微生物利用的碳源相似,且与BL、FL显著不同。此外BL和FL处理0~15 cm和15~35 cm土层分布在不同区域,表明BL和FL处理不同土层土壤微生物群落代谢显著不同。
3 讨论
土壤微生物群落的功能多样性是衡量土壤微生物群落特征的一个重要指标,它反映了群落的动态变化,对评估土壤生态系统的稳定性具有重要的意义[21]。本研究中不同植被类型在0~15 cm和15~35 cm土层土壤微生物的AWCD值在整个培养期间均表现为自然恢复(GL)>农作物(AL)>人工林(FL)>无植被覆盖(BL)(图1)。自然恢复处理的AWCD值最高,这是由于自然恢复处理植被种类丰富,有羊草(Leymuschinensis(Trin.)Tzvel.)、苔草(Carexspp.)、木贼(EquisetumhyemaleL.)和狗尾草(Setariaviridis(L.)Beauv.)等植被分布,其凋落物和根系分泌物的数量和种类较丰富,所以土壤微生物对碳源的利用能力相对于其它处理高[22]。人工林处理以人工种植落叶松为主,其AWCD值稍低于农作物(图1),原因可能是相对于玉米和大豆等农作物的的凋落物,针叶类树种的凋落物中含有大量难分解的木质素和单宁酸,降低了土壤微生物对碳源的利用能力[23]。然而刘倩[24]在通过对内蒙古地区天然草地、水田、林地、开垦4年农田和开垦30年农田土壤微生物多样性研究,结果表明土壤微生物的AWCD值的变化表现为:4年农田>天然草地>林地>30年农田,与本研究的结果不同,这可能与本研究是所处的气候条件、土壤类型以及植被种植的年限不同造成的[25]。无植被覆盖处理土壤微生物代谢活性最低,主要是由于长期无植被种植所以几乎没有碳源的输入[26-27]。可见植被对土壤微生物碳源代谢的影响与植被丰富度、根系分泌物及凋落物等来源的碳源输入都有直接的关系。此外土壤深度和土壤有机质含量也是影响土壤微生物代谢能力的重要因素[28]。郝翔翔等[29]的研究表明,草地、农田、裸地和林地的土壤有机碳含量随着土壤深度的增加呈下降的变化趋势,而土壤有机碳含量与土壤微生物群落功能多样性具有正相关关系,即土壤微生物功能多样性随着土壤深度的增加呈下降的变化趋势[30]。本研究中不同植被类型下0~15 cm土层土壤微生物的AWCD值均高于15~35 cm土层,推测其产生原因是由于植被的存在使得土壤表层含有大量的凋落物促进了土壤的有机碳的形成;种植植被可以减少土壤团聚体的破碎,在一定程度上减少了土壤养分的分解[31],促进了土壤有机碳的积累和增加,而土壤有机碳含量高土壤微生物及其功能多样性就越高[32]。此外表层土壤的透气性一般较深层土壤更好,更有利于土壤微生物的繁殖和存活。陈超凡[31]等通过对4种阔叶人工林不同土层深度土壤微生物功能多样性的研究表明,随着土层深度的增加土壤微生物可利用的养分降低、根系分泌物量减少,导致微生物可利用的数量和种类下降,从而引起各林分土壤微生物对碳源的利用能力下降。健康良好的土壤环境可以促进土壤微生物的繁殖与生长,有利于保持和提高土壤微生物的活性和功能多样性,同时土壤微生物又会促进土壤养分积累、转化和土壤结构的改善,进而有利于土壤生态系统的稳定[11]。

注:空心代表0~15 cm土层,实心代表15~35 cm土层。Note:Hollow means 0~15 cm soil layer,solid means 15~35 cm soil layer.图3 不同植被类型下不同土层土壤微生物对31种碳源代谢主成分图Fig.3 Principal component analysis of 31 carbon sources metabolism by soil microorganisms in different soil layers of different vegetation types
不同植被类型下不同土层土壤微生物对碳源的利用有差异。本研究中自然恢复、农作物、人工林及无植被覆盖处理不同土层土壤微生物对碳源的利用存在偏好,农作物处理0~15 cm土层对羧酸类利用最高,这与孟庆杰等[27]在黑土农田上的研究结果相似。然而陆思旭等[33]研究表明,农田黑土土壤微生物利用最高的碳源是糖类,此外种植柑橘的土壤微生物对于多聚物和羧酸类碳源的利用率最高[34],产生差异的原因是由于植被、取样时期及土壤类型等不同引起了土壤中的微生物群落组成发生变化,从而导致土壤微生物对碳源的利用能力不同[35]。在本研究中不同植被类型条件下0~15 cm土层土壤微生物总的碳源利用能力均显著的高于对应处理在15~35 cm土层的土壤微生物,BL处理0~15 cm主要利用的碳源是氨基酸类,而15~35 cm则是酚酸类利用的最高。有植被存在的条件下不同处理对氨基酸类和羧酸类碳源利用的较多,但其利用能力显著高于无植被覆盖处理。有研究表明水稻根际微生物对氨基酸的利用率较高,而非根际土壤对羧酸类、酚酸类和胺类的利用率较高[36],可见植被的存在对土壤微生物群落功能多样性的影响作用显著,其内在机理还需要做进一步的深入研究。整体上有植被的自然恢复、农作物和人工林处理的土壤微生物碳源代谢功能相似,这可能由于Biolog法培养的都是土壤中快速繁殖的优势菌群,而土壤中的优势菌门一般不会随着植被和管理措施而发生显著变化[37-38]。虽然Biolog法能够通过碳源代谢能力反映土壤微生物群落的变化,但是仍然存在很多的不足,今后还需借助于高通量测序及磷脂脂肪酸PLFA等技术,结合合理的统计分析方法,从不同层面对土壤微生物功能多样性展开研究[39]。
4 结论
采用Biolog-Eco微平板法研究了自然恢复(GL)、农作物(AL)、人工林(FL)及无植被覆盖(BL)处理下不同土层土壤微生物群落功能多样性的变化,得到以下结论:
(1)不同处理在整个土层中AWCD值变化均表现为自然恢复>农作物>人工林>无植被覆盖,且表层0~15 cm的AWCD均显著高于15~35 cm。
(2)Shannon指数、McIntosh指数、Simpson′s指数和Richness指数均表现为自然恢复、农作物、人工林处理显著高于无植被覆盖处理,表层0~15 cm土壤微生物代谢活性和群落功能多样性均高于15~35 cm。
(3)两个土层中土壤微生物群落主要利用的碳源有氨基酸类、羧酸类、糖类和多聚物类。在0~15 cm土层BL处理土壤微生物对氨基酸类利用最高,而AL和FL处理对羧酸类碳源利用最高,自然恢复处理对氨基酸类利用最高;在15~35 cm土层无植被覆盖处理对酚酸类利用最高,农作物和人工林处理对氨基酸类利用最高,自然恢复处理对羧酸类利用的最高。
(4)主成分分析结果表明,自然恢复、农作物和人工林处理在0~15 cm土层的土壤微生物的碳源代谢功能相似;自然恢复和农作物处理在15~35 cm土层的土壤微生物的碳源利用方式相似,与无植被覆盖、人工林处理均显著不同。
总之不同植被类型下土壤微生物群落功能多样性的差异是植被类型与土层深度共同作用的结果,其中以自然恢复处理对黑土土壤微生物群落功能多样性影响最为显著,更有利于黑土土壤生态系统的稳定。
猜你喜欢
油气田地面工程(2022年9期)2022-10-26
当代水产(2022年8期)2022-09-20
江苏农业科学(2022年6期)2022-04-15
昆明医科大学学报(2022年2期)2022-03-29
铀矿冶(2022年1期)2022-02-17
乡村科技(2021年1期)2021-12-03
农村实用技术(2021年6期)2021-12-01
昆钢科技(2021年6期)2021-03-09
学校教育研究(2020年7期)2020-04-09
农民致富之友(2020年4期)2020-04-03
