
黄土高原苹果叶片光合作用限制形式研究
2022-03-08 09:20王姝婷李伟高冠龙
山西大学学报(自然科学版) 2022年1期
王姝婷,李伟,高冠龙,2,3,4*
(1.山西大学 环境与资源学院,山西 太原 030006;2.长安大学 陕西省土地整治重点实验室,陕西 西安 710064;3.中国科学院 西北生态环境资源研究院,甘肃 兰州 730000;4.山西省黄河实验室,山西 太原 030006)
0 引言
黄土高原是我国著名的苹果产区,具备了生产优质苹果的自然条件(海拔高、光照足、昼夜温差大、污染轻)。然而,黄土高原作为我国半干旱地区的主要组成部分,缺乏充足的水分来源,加之特殊的气候条件,土壤水分成为了制约苹果树生长的主要因素。因此,黄土高原半干旱地区苹果树的光合作用就容易受到明显的限制。
水分胁迫对苹果树光合作用的限制形式主要包括气孔限制和非气孔限制,两者的共同作用使得苹果叶片对光能的利用率减弱,影响苹果树的生长。判别植物受到气孔限制或者非气孔限制的核心就是Ci值的变化。不少学者们针对水分胁迫下植物生长的限制形式进行了研究,得出了一个普遍结论:在轻度、中度水分胁迫的环境下,植物光合作用的限制形式主要是气孔限制,而在重度水分胁迫的环境下,主要是非气孔限制,在一些特殊条件下,二者又相伴而生,共同影响着植物的光合作用[1-3]。
国内对苹果树叶片光合作用的研究[4]多在不同环境条件下进行,主要集中在干旱胁迫[5]、盐碱复合胁迫[6]、蓄水坑灌[7]、外源激素[8]及地面覆盖[9]等方面,缺乏对自然条件下苹果树叶片光合作用限制因子的研究。因此,本文在自然环境条件下,以黄土高原苹果树叶片为试验对象,研究果实膨大期和成熟期光合作用的限制形式及其影响因子,以期了解自然环境条件下苹果树叶片光合作用的限制机理,为黄土高原半干旱地区苹果树的生长和管理提供科学依据。
1 材料与方法
1.1 研究区概况
试验区位于山西省农科院植物保护研究所东阳试验研究基地(地理坐标:37°32′N、112°41′E),地处山西省中部。气候类型属于温带季风气候,四季变化明显,雨热同期,夏秋季降水集中,冬春季较干旱,平均年降水量为450 mm,空气相对湿度达60%;年均气温为9.5℃,年均风速为2.4 m/s,最少日照数为253.5 h。试验区内苹果树平均冠幅为316 cm×318 cm,平均地径为10.9 cm。
1.2 试验过程与方法
试验于2019年7月-10月苹果生长的两个主要阶段(果实膨大期和成熟期)内进行,苹果树的生长季划分参考党宏忠的研究结果[10],具体见表1。选择3株长势较好的苹果树,采用美国LI-COR公司生产的LI-6400便携式光合作用测量仪,选择3片成熟叶片并始终保持叶面垂向于太阳,每片叶片记录三次数据,测定苹果树叶片的气体交换特性参数,包括净光合速率(Pn,μmol·m-2·s-1)、气孔导度(gs,mol·m-2·s-1)、蒸腾速率(Tr,μmol·m-2·s-1)、Ci(μmol·mol-1)和大气中 CO2浓度(Ca,mmol·mol-1),观测时段为8:00-18:00,每隔 1 h测量 1次。此外,基地内设有小型气象站以观测各气象因子,包括光合有效辐射(PAR,μmol·m-2·s-1)、饱和水汽压差(VPD,kPa)、空气温度(Ta,℃)和空气湿度(RH,%)。

表1 黄土高原地区苹果树生育期划分标准Table 1 Division standard of apple tree growth period on Loess Plateau
1.3 气孔与非气孔限制值的计算
气孔限制值(Ls)反映了因气孔开度减小,气孔阻力增加[11],进入叶片内的CO2减少而引起光合速率减弱的气孔因子限制情况。目前,计算Ls的公式为[12]:
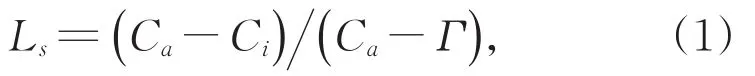
式中:Γ为CO2补偿点。但实际计算中Γ常被忽略,表达式又可以写成:

非气孔限制值反映了光合器官的变化引起的光合作用降低的非气孔因子限制情况,常用Ci/gs来表征[2-3]。
2 结果与分析
2.1 不同生长期苹果树环境因子日变化
在苹果膨大期和成熟期各选取4 d(7月31日、8月13日、8月20日、8月28日和9月1日、9月8日、9月22日、10月19日)的数据,进行环境因子和生理因子的日变化分析,结果见图1。
由图 1可以看出,PAR(图 1a)、VPD(图1b)、Ta(图1c)的日变化都呈先增大、后减小的趋势,PAR的峰值出现在中午12:00-13:00,而 VPD、Ta的峰值均出现在 12:00-17:00,且中午、下午明显较大。RH的日变化趋势(图1d)与PAR、VPD、Ta相反(上午增大,随后逐渐降低,最低值出现在 13:00-17:00,之后逐渐增大)。果实膨大期和成熟期内PAR和VPD都很 大,PAR 峰 值 为 1 154.369 μmol·m-2·s-1~1 473.263 μmol·m-2·s-1,平 均 值 为 1 065.893 μmol·m-2·s-1,VPD 峰 值 为 2.563 kPa~4.149 kPa,平均值为2.489 kPa。在水资源相对短缺的半干旱地区,VPD作为判断叶片需水量的重要因子,其值越大,表明叶片缺水越严重。两个生长期内Ta很高,中午时段的Ta值在35℃左右甚至更高,因此导致叶片水分蒸发较快,RH值降低。Ta和RH的变化趋势相反,二者在果实膨大期和成熟期出现峰值和谷值的时间不同,果实膨大期均早于果实成熟期。与此同时,VPD也从侧面反映出Ta和RH对苹果树叶片的影响。当Ta越大时,RH就越小,叶片对水分的需求就越大,而此时的VPD也越大。
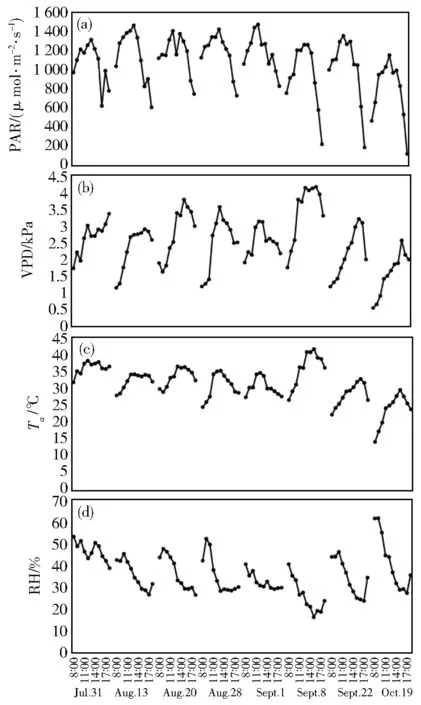
图1 (a)光合有效辐射(PAR)、(b)叶片饱和水汽压差(VPD)、(c)空气温度(Ta)、(d)空气相对湿度(RH)的日变化Fig.1 Diurnal variations of(a)photosynthetically active ra⁃diation(PAR),(b)vapor pressure deficit(VPD),(c)air temperature(Ta)and(d)relative humidity(RH)
2.2 不同生长期苹果树叶片生理因子日变化
图2为不同生长期苹果树叶片生理因子日变化趋势。Pn在果实膨大期和成熟期大致都呈先升高后降低的变化趋势(图2a),且都有“双峰”,出现了短暂的“光合午休”现象。gs在两个生长期呈现出与Pn相同的变化规律(图2b),gs值在12:00左右波动幅度较大,均出现了低谷,是由于午间太阳辐射强,空气温度高,叶片因水汽亏缺大而关闭气孔防止水分流失;此外,上午的gs值整体上要比下午的大。Tr的变化规律与Pn和gs的规律相似(图2c),整体上呈现出先升高后降低的变化趋势,也有“双峰”。午后Tr降低,是受到气孔关闭的影响,而下午Tr的减小也离不开太阳辐射减弱的影响。Ci的日变化趋势与Pn和gs的相反(图2d),在两个生长期内的变化趋势都是早晚大,下午小,呈现出“V”型的变化规律,最小值出现在14:00-17:00,整体变化差异较大。

图2 (a)净光合速率(Pn)、(b)气孔导度(gs)、(c)蒸腾速率(Tr)和(d)胞间CO2浓度(Ci)的日变化Fig.2 Diurnal variations of(a)net photosynthesis(Pn),(b)stomatal(gs),(c)transpiration rate(Tr)and(d)substomatal CO2concentration(Ci)
2.3 苹果树叶片光合生理特征对环境因子的响应
VPD是表征叶片水汽亏损的重要指标,对气孔的大小能够产生很大的影响。图3为gs与VPD的线性关系图,可以看出:在苹果树两个生长期内,gs与VPD成负相关关系,gs随VPD的增大而逐渐减小。整体来看,上午Ta较低,RH较大,叶片饱和水汽压差小,gs值也相对较大;下午随着Ta升高,VPD逐渐增大,受其影响gs逐渐下降,此后VPD虽有所降低,但其值仍高于上午,而gs一直处于下降趋势,这表明半干旱地区苹果树叶片在果实膨大期和成熟期的午间时段,由于空气温度增高,叶片失水过快而导致气孔关闭。
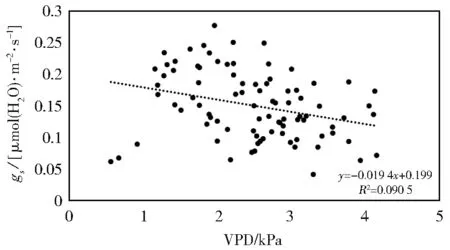
图3 苹果树叶片气孔导度(gs)和饱和水汽压差(VPD)的关系Fig.3 Relationship between stomatal conductance(gs)and vapor pressure deficit(VPD)of apple tree leaves
图4为苹果树叶片两个生长期内Pn对gs的响应关系,从而可以看出,Pn与gs呈对数型曲线,Pearson相关系数为 0.547**(P<0.01),Pn随gs的增大而增大,但是增长速度却随gs的增大而逐渐放缓。在两个生长期的上午,Pn随gs的增大而增大,随后二者都达到峰值,但之后Pn受气孔逐渐关闭的影响逐渐减小,出现“光合午休”。
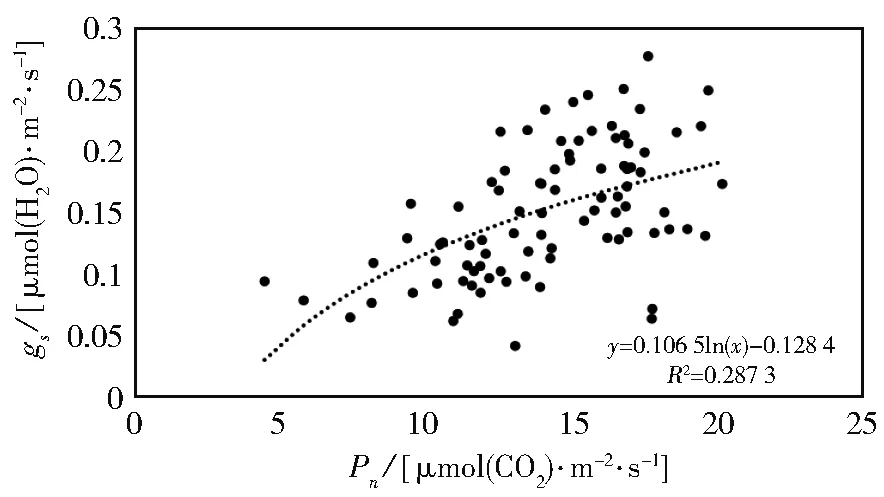
图4 苹果树叶片气孔导度(gs)与净光合速率(Pn)的关系Fig.4 Relationship between stomatal conductance(gs)and net photosynthesis(Pn)of apple tree leaves
2.4 不同阶段光合作用的气孔限制与非气孔限制
由图5可以看出,在果实膨大期和成熟期内,Ls从早间开始逐渐增大,在 14:00-17:00达到峰值,之后虽呈下降趋势,但Ls在下午仍较大,这表明气孔限制是影响全天光合作用最重要的因子。Ci/gs的变化趋势与Ls正好相反,从早晨开始变化一直不大,在正午时段稍微有所上升,在Ls达到峰值的同时,Ci/gs下降到低谷,随后持续升高,增长幅度较大。

图5 苹果树叶片气孔限制值(Ls)和非气孔限制值(Ci/gs)的日变化Fig.5 Diurnal variations of stomatal(Ls)and non-stomatal limitations(Ci/gs)to photosynthesis of apple tree leaves
3 讨论
3.1 主要生理因子及“光合午休”现象
Ci作为判别叶片光合作用气孔或者非气孔限制的主要因素,在试验期间呈“V”字型变化规律,此结论与其他作物的研究结果一致[13-15]。外界CO2进入叶片叶绿体内包括两个阶段:一是Ca通过气孔向Ci扩散,二是Ci通过叶肉细胞向叶绿体内扩散 CO2(Cc),因此 Ci值的大小与 gs和叶肉导度(gm)有关[13]。当 gs减小,Pn降低,Ci也呈下降趋势时,表明气孔限制是影响叶片光合作用的主要因子;如果Ci升高,而gs与Pn仍降低,则表明造成此时光合速率下降的主要原因是叶肉细胞光合能力的降低,即非气孔限制因素的影响[16]。有研究表明,gm是非气孔限制因子中的不可或缺的因素[17],空气越干燥,VPD 越大,gm就越小[18-19]。上午VPD较下午小,因此gm较大,外界CO2通过气孔进入Ci,通过叶肉细胞进入叶绿体内,对CO2消耗加大,因此Ci降低,下午VPD虽有降低但一直居于较高水平。因此gm较小,阻碍CO2向细胞内部扩散,非气孔限制增强,Ci值增大。
两个生长期Pn在13:00左右都出现了短暂的“光合午休”现象,同时Ci呈下降趋势,值较小,此时受Ta增高和VPD过大的影响,气孔关闭,gs减小。因此可以推断,苹果树叶片出现“光合午休”现象主要是受气孔限制因素的影响[13]。虽然gs的日变化呈现周期性波动,是黄土高原半干旱地区苹果树自身适应干旱的自行生理调节机制[20],但强烈的气孔限制仍旧影响着光合产物的转化,“光合午休”现象无法从根本上消除,因此需要通过改善栽培结构,利用不同农作物的生理特征,发展间作和套种模式,加强光合作用,改善局地小气候,适当补给水分,减缓气孔限制的影响,提高果实品质,增加农业效益[21-22]。
3.2 光合作用的气孔限制和非气孔限制
Ls在两个生长期内的8:00-15:00,都呈上升趋势,这是由于早间Pn逐渐增大,此阶段gs和 gm都较大,导致 Ca向 Ci、Ci向 Cc的扩散速度加快,叶片的光合作用暂未受到限制。因此,Ls值升高反映了Ci的过度消耗;午间由于太阳辐射增强、VPD增大,气孔逐渐关闭,gs减小,Pn随gs的降低而逐渐减小,出现“光合午休”现象,但 gm相对较大,因此 Ci仍向 Cc扩散,Ci不断降低,表明这两个时间段内光合作用的降低以气孔限制为主导。下午,Ls逐渐降低,但其值仍较大,表明下午时段的气孔限制也不容忽视。Ci/gs在 8:00-14:00稍有上升,但整体趋势较缓,表明该时间内非气孔限制较小;在15:00左右逐渐增大,增长速度较快,表明该时段内非气孔限制占据主导,这是由于下午VPD仍较大,gs与gm都很小,限制了CO2向叶绿体内的扩散,Ci值逐渐回升。
本研究对果实膨大期和成熟期中午均出现的非气孔限制的原因以及造成非气孔限制的其他因素如叶绿体活性[23]、Rubisco活性、RuBP再生能力[2,23]以及植物含水量、叶绿素含量等植物光合生理指标的数据并未测定,希望以后进行研究。
由于试验条件的限制,本文仅探讨了2019年7月-10月自然条件下苹果树叶片光合生理特性的日变化规律及对环境因子的响应,所得到的结果可以为了解苹果树生长期的光合作用限制因素提供一定参考,但数据研究的时间跨度较短,仅能表示苹果树两个生长期的光合特性变化规律。在后续研究中将对苹果树结果前期的叶片光合特性以及其他光合生理因子如植物含水量和叶绿素含量等因子对环境的响应规律进行研究,来进一步完善苹果树整个生长期的生理特性变化数据和环境响应规律,以期获得更加普遍和更具有代表性的规律。
3.3 苹果树生长和管理措施
3.3.1 覆草、生草
黄土高原地区是我国优质苹果的产地之一,但该地区气候干旱,降水不均,土壤疏松,水土流失严重,使得裸露土层底部含水量降低,土壤干燥化加重,难以调蓄苹果园地下的水分,通过覆秸秆和生草可以增加苹果园土壤孔隙状况及贮水库容,建立果园覆盖技术体系,改善果园土壤水分[24],此外,地面覆盖还可以提高土壤肥力[25],影响苹果树的生长和生理状况[26]。面对黄土高原地区特殊的气候,应因地制宜制定不同覆盖方式来保持水土,调节土壤水分含量,减轻果实膨大期和成熟期内因空气温度升高湿度降低而导致的“光合午休”带来的一系列影响,增加空气湿度。
3.3.2 修剪
对果树进行整形修剪,不仅能够影响光合产物的积累量,又能合理调配养分在营养器官和生殖器官之间的分配[27]。若没有合适的整形修剪,苹果树的生长(光合作用、养分分配)必然会受到明显的抑制,最终导致果实品质降低,苹果树发育缓慢。在苹果树管理方面,应根据苹果树生长状况及时对其进行修剪,合理调整树冠结构,进而改善土壤水分,保证枝条能充分汲取营养,提高苹果树果实膨大期和成熟期叶片的光合速率,提高果实坐果率,确保苹果的质量和产量。
3.3.3 间作
研究表明,农林复合生态系统在提高土地生产力、解决土地矛盾和改善生态环境等方面有重要的促进作用[28]。果树下间作其他作物,能够有效改善果园小气候,提高小系统内的空气湿度,并且间作作物的气温日变化幅度小[22],在高温期(7月-10月果实膨大期和成熟期)能够有效防止高温对苹果的不利影响。从表2可以看出,果实膨大期和成熟期内午间时段净光合速率和蒸腾速率都降低,会影响到苹果糖分等光合产物的积累,因此,采取不同的间作方式,可以改善苹果树周围的小气候,使周围空气温度降低湿度增加,蒸腾速率加快,减弱“光合午休”现象,积累更多的光合产物,提高果实质量。另外,间作其他作物,也能增加相应的经济效益,增加收入。
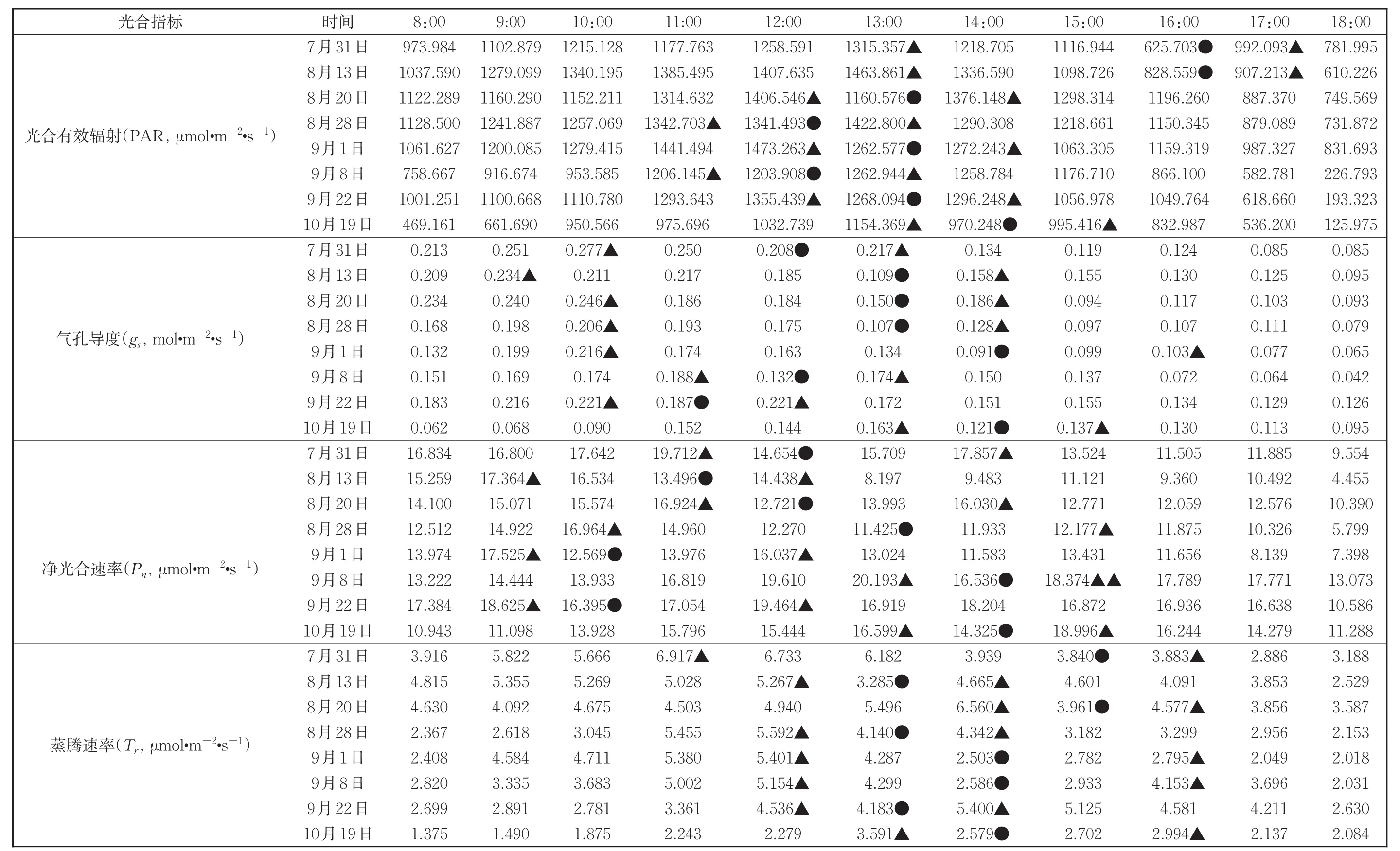
表2 不同时期各个光合生理指标的差异性Table 2 The differences of photosynthetic physiological indexes in different periods
3.3.4 合理选择灌溉方式
研究表明,苹果树在不同生长期对水分的需求也不同,在生长中期(果实膨大期)需水量最大,其次为果实成熟期[10,29-30],且果实膨大期内土壤水分的供应量直接影响果实的最终大小和产量[10]。在果园内部完善灌溉基础设施,合理选择灌溉方式,能够调配果树正常发育对水分的需求量。因此,因地制宜合理选择灌溉方式既可以达到节水的效果,又能够通过补充土壤水分来提高果实品质,具有很大的推广意义。
4 结论
(1)苹果树叶片光合作用的限制因子在两个生长期内出现的时间不同。Ls呈先增大后减小的趋势,上午逐渐增大,午后达到最高值后逐渐下降,但值仍较大,表明光合作用在上午及午后主要受气孔限制影响,且受气孔限制影响的时间也较长,果实膨大期中午时段气孔限制最强烈,而果实成熟期主要是在下午最强。Ci/gs在上午较小,变化幅度不大,下午逐渐升高且斜率较大,表明光合作用的限制形式由气孔限制转为非气孔限制为主。
(2)在两个生长期内都出现了“光合午休”现象,主要是受气孔限制的影响,由于过大的PAR和VPD,导致气孔逐渐关闭,但此时非气孔限制因素的影响也不容忽视。面对“午休”现象,管理者可以通过增加覆草、生草面积提高土壤中水分的含量,保证苹果树在水胁迫条件下对水的需求量,并且可以在早期对苹果树进行更新修剪,积极实行间作种植模式,合理设置灌溉方式,提高果实品质,实现规模效益。
猜你喜欢
航空材料学报(2022年4期)2022-08-04
贵州农业科学(2022年4期)2022-05-06
干旱地区农业研究(2022年1期)2022-01-28
房地产导刊(2020年9期)2020-10-28
当代工人(2020年11期)2020-07-06
汉语世界(The World of Chinese)(2019年5期)2019-11-11
科技创新与品牌(2019年12期)2019-02-06
中国计算机报(2017年47期)2018-01-12
电影文学(2016年23期)2017-02-13
作文大王·中高年级(2016年11期)2016-12-09
