
中国温带典型森林植物比叶面积的空间格局及其影响因素
2022-03-07 13:15刘可佳何念鹏侯继华
生态学报 2022年3期
刘可佳,何念鹏,侯继华,*
1 北京林业大学生态与自然保护学院,北京 100083 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101
植物功能性状(Plant Functional Trait,PFT)是能够揭示植物响应变化环境的生态策略且易于测量的植物特征[1—2]。叶片是植物直接接触大气环境的器官,是植物进行光合作用和气体交换的基本功能单位,是维持生态系统运转的枢纽[3]。而叶功能性状作为主要的植物功能性状之一,与植物的生长、存活和对光的需求密切相关[4],能够反映植物对现有资源的利用分配能力和对环境变化的适应对策[5—6]。
比叶面积(Specific Leaf Area, SLA)是关键的叶功能性状指标之一,被定义为叶面积与其自身干重之比[7],不仅可以表征植物对生长前期捕获光、养分等资源的预期回报,而且能够反映植物在不同资源环境下的碳获取策略、生长策略和适应策略的权衡[8]。生态学者针对SLA这一指标从不同角度开展了大量的研究工作,包括SLA的空间变异[9—11]、种内种间变异[12]、SLA与其他植物性状的关联[11, 13]和环境变化对SLA的影响[14—15]等多个方面。研究结果表明,在全球尺度上,随着空间的变化,不同区域植物SLA存在显著差异[9];SLA随纬度的增加逐渐增加[11],而随着海拔的升高,SLA显著下降[10]。不同树种SLA也存在显著差异,落叶树种的SLA显著大于常绿树种,阔叶树种的SLA显著大于针叶树种[11—12],各功能性状间也存在明显联系,SLA大的植物,往往具有较低的叶干物质含量和叶氮含量,它们之间相互协作以适应环境[13, 16]。环境因子是影响植物SLA变异的极为重要因素,通常情况下,SLA随着温度的升高而增加;生长在湿润和低辐射条件下的植物具有较大的SLA[14—15]。这些研究极大地提升了人们对植物生理活动、养分利用策略及植物对全球变化的响应与适应策略的理解。需要关注的是,当前已有的大多数植物功能性状研究仍是以少数优势物种为主要研究对象[17—18],并未考虑天然森林群落物种组成的复杂性和区域差异性。仅仅关注优势种会弱化对生态系统整体性的认识,而生态系统的功能和服务最终取决于组成生态群落的物种的特征,因此对植物群落所有物种SLA进行系统性的调查对探究SLA的空间变异及其影响因素至关重要。
中国东北部的温带森林是中国森林资源的重要组成部分,也是东北大平原农业发展的重要屏障, 对区域气候环境的维护和调节起着重要的生态作用[19—20]。这一地区典型的地带性植被为寒温带兴安落叶松林、温带红松针阔混交林和暖温带落叶阔叶栎林。本研究以我国东北地区三种典型的森林作为研究对象,设置覆盖我国东北地区约1200 km的野外调查样带,并向南扩展到北京东灵山,共设置12个调查样点,通过详细的群落调查及群落内所有物种比叶面积的系统性调查与测定,重点探究如下科学问题:温带森林SLA在空间格局上的变化规律,东北地区3种典型森林类型植物SLA的差异以及SLA空间变异的影响因素。以期为未来深入研究植物对环境变化的适应对策及机制提供理论支撑。
1 研究地区及其概况
1.1 研究区概况
研究地区位于经度115°—129°E,纬度39°—54°N,跨暖温带、中温带、寒温带3个温度带,海拔范围在316—972 m,年平均温为-5.47—6.55℃,年均降水范围在383.53—836.23 mm。根据三个温度带的典型地带性森林的空间分布特征和代表性,调查主要选择在远离村落、人为干扰少的国家级自然保护区或林场内进行,主要选择保存较好、区域代表性强的成熟林。共布设12个采样点(图1)。具体包括:北京市门头沟区东灵山,河北省兴隆县雾灵山自然保护区、辽宁省清原满族自治县、吉林省安图县长白山自然保护区、吉林省辉南县龙湾国家森林公园、吉林省蛟河市、黑龙江省尚志市帽儿山国家森林公园、黑龙江伊春市凉水自然保护区、内蒙古自治区根河市、内蒙古自治区莫尔道嘎国家森林公园、黑龙江省呼玛县呼中自然保护区、黑龙江省漠河市。各研究区基本信息详情见表1。
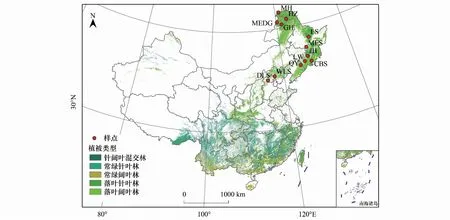
图1 典型温带森林采样点分布图Fig.1 Distribution of sampling sites in typical temperate forestsDLS:东灵山;WLS:雾灵山;QY:清原;CBS:长白山;LW:龙湾;JH:蛟河;MES:帽儿山;LS:凉水;GH:根河;MEDG:莫尔道嘎;HZ:呼中;MH:漠河
1.2 研究方法
1.2.1野外采集
本研究于2019年植物生长旺盛的7—8月进行野外调查与取样。按照纬度从南到北的顺序依次对不同样点进行野外调查与取样。在每个取样点设置4个30 m×40 m调查样地,进一步划分为次一级样方。为了便于区分,将原样地称为一级样方(30 m×40 m),用于乔木层的调查; 在每个一级样方内设置2个5 m×5 m的二级样方,用于灌木层的调查;设置4个1 m×1 m的三级样方,用于草本层的调查。群落结构调查方法参见《中国生态系统研究网络(CERN)监测规范》。调查后采集样地中出现的所有植物的叶片样品。乔木叶片样品采集:每个树种选择样地内胸径、树高最大、长势良好的个体3—4株。每株个体根据不同层次和方位,用高枝剪或人工爬树的方法采集树冠中上部东南西北4个不同方向的小枝4个,手工摘取无病虫害的、完全展开的健康成熟叶片,并且把不同方位的叶片样品按比例均匀混合,获取叶片的混合样品。红松和油松仅对多年生叶进行取样。灌木叶片样品采集:选取样地中每个灌木物种冠幅、株高最大的个体3—4株。每株个体采集灌木中部偏上、无病虫害、完全展开的健康成熟叶片。草本植物叶片样品采集:选取样地中常见物种基径、冠幅最大的5—10株个体。每个个体采集无病虫害、完全展开的健康成熟叶片。野外采集叶片后,立刻将采集到的叶片放入内置冰块的保温箱(或冷藏箱)(0—4℃)中保存。共采集物种数1086种,其中乔木174种,灌木269种,草本643种。具体采样方法参照何念鹏等[21]。
同时,在每个样方中收集一份土壤样品。取样时,先除去表面的凋落物,然后用土钻法钻取0—10 cm表层土壤,手动挑除其中可见的根系和凋落物。为减少土壤异质性,将每个调查点4个样方中钻取的土壤样品混合得到一个混合土样。之后将混合土样自然风干并通过直径2 mm的土壤筛,用于土壤理化性质的测定。
1.2.2室内测定
阔叶树种用扫描仪(Cano Scan LIDE 220, Japan)扫描获得叶片图像,利用Image J软件(National Institutes of Health,Bethesda,USA)计算叶片的面积,然后放入75℃的烘箱中烘干至恒重,用万分之一天平测量其干重。落叶松、云杉等可近似看作长方形,随机挑选30片叶子测量其长和宽,计算总叶面积;红松单针横截面可近似看作三角形,测量每根针叶长和横截面外边长,得到单叶面积,共取30片针叶计算其总面积,烘干后得到总干重。根据公式SLA=叶面积/叶干重,得到比叶面积。用元素分析仪(Vario EL Ш, Elementar, Germany)测定土壤碳、氮的含量,用钼锑抗比色法测定土壤磷含量,用FP 6410火焰光度计(上海仪电分析仪器有限公司)测定土壤钾含量,土壤酸碱度用pH计(Myron L. Company, Carlsbad, CA, USA)测定。
1.2.3环境要素的获取
12个调查点的气象数据来自1961—2010年中国气象站数据插值结果,包括年均温度(MAT)和年均降水量(MAP);通过 Science Data Bank网站(http://www.dx.doi.org/10.11922/sciencedb.332)提取连续15年的辐射数据,包括年均紫外辐射(UV)和光合有效辐射(PAR);土壤理化性质数据由实验室测得。
1.2.4数据分析方法
根据生活型将森林植物划分为乔木、灌木和草本植物,根据叶片形态特征和脱落特征将树种分别分为阔叶树种和针叶树种、常绿树种和落叶树种[22—23]。首先探讨温带森林不同生活型植物SLA的分布规律,之后采用回归分析探讨不同生活型的植物SLA随空间格局的变化规律,采用单因素方差分析(one-way ANOVA)和多重比较(Duncan法)探讨植物SLA在不同森林类型间的差异,采用回归分析探讨环境要素对植物SLA的影响,并进一步利用结构方程模型评估气候、辐射、土壤因子对植物SLA空间变异的解释程度。由于结构方程模型要求解释变量与被解释变量之间满足线性相关关系,因此对具有二次项相关关系的变量进行开方处理,以满足模型要求。此外,为避免环境因子之间因相关性高而在模型建立时产生共线性问题,对其进行降维处理或选择其中一个因子进入主效应。最终,通过逐步回归选择具有显著作用的因子作为最终模型的参数。数据统计分析采用SPSS 25.0完成,图的绘制采用R 4.0.0、Origin 2018和AMOS 24软件进行。显著性水平为P=0.05。
2 结果
2.1 温带典型森林植物 SLA的整体分布特征
中国温带典型森林的植物SLA总体上呈现正态分布趋势(图2)。比叶面积整体变化范围为2.02—99.65 m2/kg,平均值及其标准差为(34.18±16.03) m2/kg,峰值为1.07 m2/kg,变异系数为0.47。乔木、灌木和草本SLA的分布特征与温带森林所有物种SLA的分布趋势一致。其中,乔木SLA变化范围为2.02—58.74 m2/kg,均值及其标准差为(21.32±10.24)m2/kg;灌木SLA变化范围为2.89—99.65 m2/kg,均值及其标准差为(31.60±14.21)m2/kg;草本植物SLA变化范围为4.66—98.53 m2/kg,均值及其标准差为(38.75±15.91)m2/kg。不同生活型的植物SLA表现为草本>灌木>乔木,这说明乔木比灌木和草本更能适应贫瘠环境。SLA的变异系数在不同生活型中表现为乔木(0.48)>灌木(0.45)>草本(0.41),说明乔木比灌木和草本植物对区域间的环境变化更加敏感,而草本植物比乔木和灌木具有更好的生态适应性。
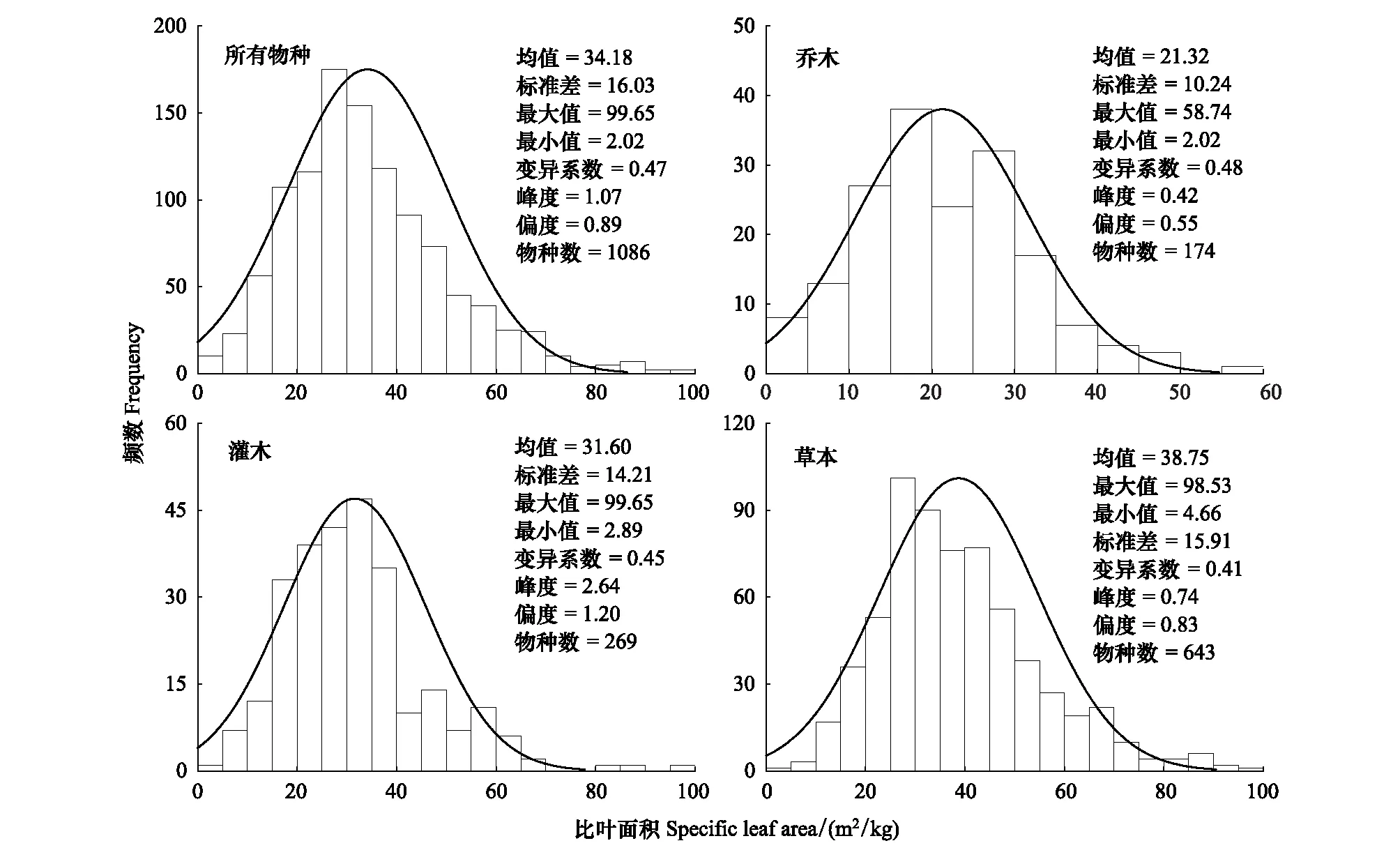
图2 温带森林植物比叶面积频数分析Fig.2 Frequency distribution of plant specific leaf area in temperate forests
2.2 温带典型森林的植物 SLA的纬度格局
中国温带森林的植物SLA表现出显著的纬度变化格局(图3)。温带森林所有物种的SLA随纬度升高呈现显著的二次项回归关系,即表现为先升高后降低的变化趋势,其中,龙湾、蛟河地区植物SLA最大。乔木SLA、灌木SLA和草本植物SLA随纬度变化趋势与所有物种SLA表现出一致的规律性,且与纬度格局均具有显著相关性。乔木SLA在龙湾、清原地区较大,在蛟河地区较低;灌木和草本SLA均在龙湾、蛟河具有最大值。
2.3 三种典型森林的植物SLA差异分析
三种典型森林的植物SLA具有明显的差异(图4),表现为温带红松针阔混交林>暖温带落叶阔叶栎林>寒温带兴安落叶松林。对于阔叶树种和针叶树种、常绿树种和落叶树种而言,其SLA也存在显著差异,但在不同森林类型中均一致表现为阔叶树种SLA>针叶树种SLA;落叶树种SLA>常绿树种SLA。阔叶树种与落叶树种SLA在不同森林类型间具有一致的变化趋势,即温带红松针阔混交林>暖温带落叶阔叶栎林>寒温带兴安落叶松林。不同森林的针叶树种SLA表现为寒温带兴安落叶松林>温带红松针阔混交林>暖温带落叶阔叶栎林;常绿树种SLA表现为温带红松针阔混交林>寒温带兴安落叶松林>暖温带落叶阔叶栎林,但两个树种的SLA在不同森林中均差异不明显。
2.4 环境因子与植物SLA的关系
温带森林植物SLA与年均降水量(MAP)呈极显著正相关关系(图5),但随年平均温度(MAT)、紫外辐射(UV)和光合有效辐射(PAR)的增加,均表现为先增加后降低的变化趋势,且植物SLA与MAT、UV和PAR之间均具有显著相关性。温带森林植物SLA也受土壤理化性质的影响(图5),植物SLA随着土壤碳、氮含量的增加呈线性增加趋势,随土壤碳氮比的升高表现为线性减少趋势,但与土壤钾、磷元素含量和土壤酸碱度无显著相关关系。说明土壤碳含量、土壤氮含量是影响植物SLA的主要土壤因子。
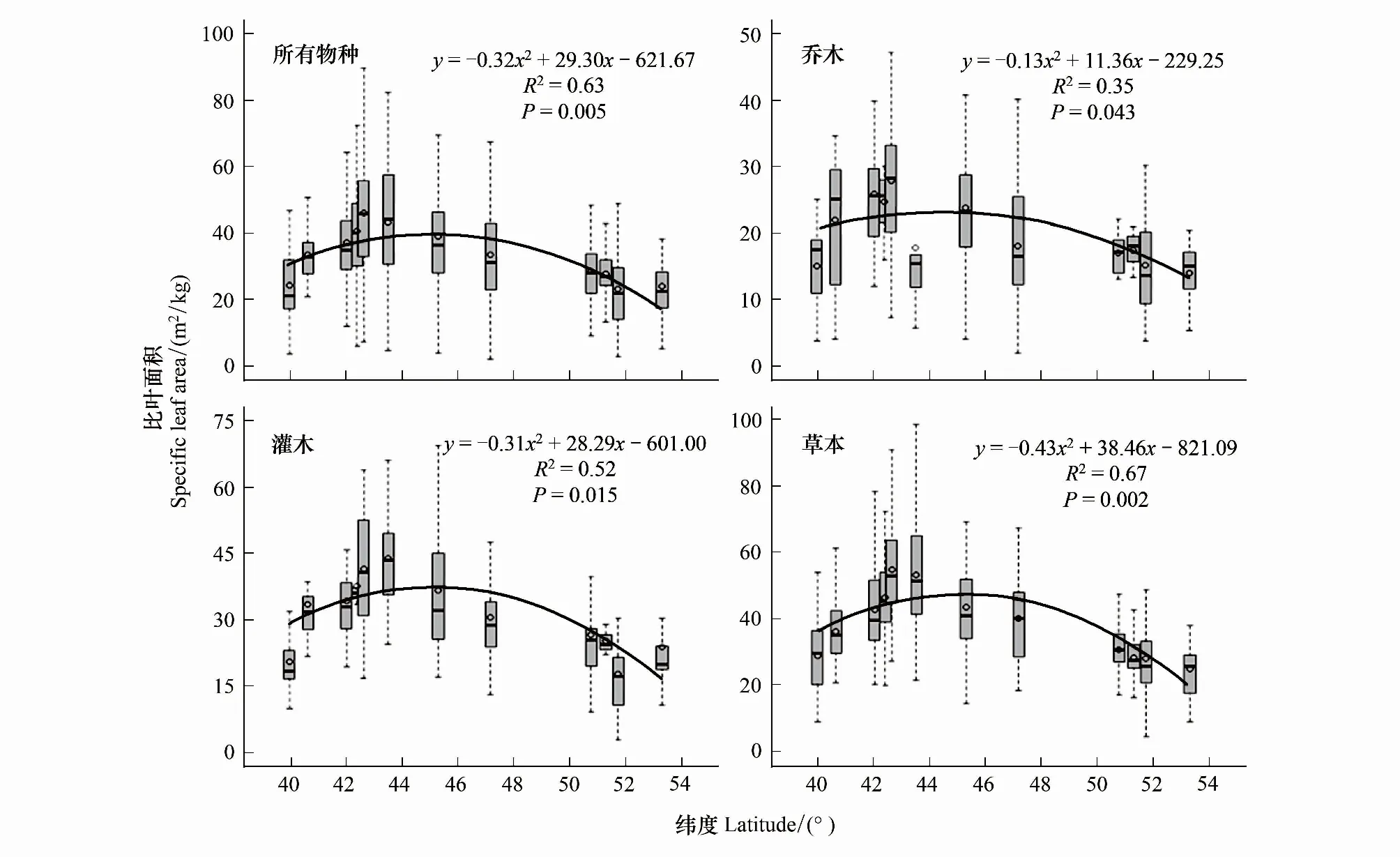
图3 植物比叶面积纬度空间格局Fig.3 Latitudinal patterns of plant specific leaf area图中箱型所代表的地区按纬度从低到高依次为东灵山、雾灵山、清原、长白山、龙湾、蛟河、帽儿山、凉水、根河、莫尔道嘎、呼中、漠河。箱线图中黑色长方形代表各分组比叶面积中值,黑色空心圆代表各分组比叶面积平均值;黑色曲线为比叶面积与纬度格局的拟合曲线
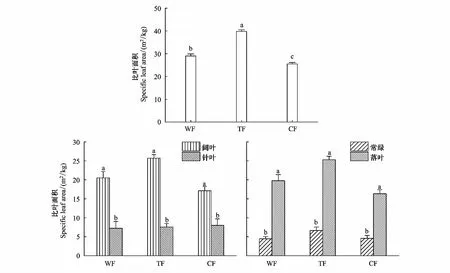
图4 不同森林类型植物比叶面积比较Fig.4 Comparison of specific leaf area among different forest typesWF:暖温带落叶阔叶栎林Warm-temperate deciduous broad-leaved Oak forest;TF:温带红松针阔叶混交林Temperate broad-leaved Korean pine mixed forest;CF:寒温带兴安落叶松林Cold-temperate Larix gmelinii forest;不同小写字母表示组内差异性显著(P<0.05)

图5 环境要素与植物比叶面积的关系Fig.5 The relationship between environmental factors and plant specific leaf area黑色线条为温带森林比叶面积与环境要素的拟合曲线,无显著相关性则不在图中显示
2.5 环境因子对植物SLA的整体影响
植物比叶面积与气候和土壤理化性质间存在显著相关性,结构方程模型结果表明气候因素(MAT和MAP)是影响植物SLA的最重要因素,能够解释植物比叶面积78%的变异(图6)。辐射、气候和土壤养分均可以直接影响植物SLA。在整个构建的模型中,气候对SLA的总效应最大(0.77),辐射次之(-0.52),土壤养分最小(0.51),三者对SLA的解释率为0.81。

图6 环境因素对植物比叶面积的影响结构模型和效应Fig.6 Structure equation modeling exploring the influence of environmental factors on specific leaf area (SLA) of plantMAP:年均降水量 Mean annual precipitation;MAT:年均温度 Mean annual temperature;TC:土壤碳含量 Soil total C content;TN:土壤氮含量 Soil total N content;PAR:光合有效辐射 Photosynthetically active radiation;UV:紫外辐射 Ultraviolet radiation;深灰色实线和浅灰色虚线分别表示正负效应;线体粗细表示标准化通径系数的大小;模型拟合参数为χ2=1.96,Df=1,P = 0.16,NFI=0.95,IFI=0.97,AIC=27.96;*和**分别表示变量间关系在P = 0.05和P=0.01水平上显著
3 讨论
3.1 温带森林植物SLA的空间变异
本研究通过对中国东北部12个典型温带森林的植物SLA进行分析研究,发现中国东北部区域尺度下植物SLA变化范围为2.02—99.65 m2/kg,平均值为34.18 m2/kg,SLA总体上呈正态分布。我们对温带森林SLA变异范围的研究结果与Wang等[24]对中国东部南北样带847个物种叶片SLA的调查结果(1.89—94.99 m2/kg)相似,与全球区域植物SLA数值的变化范围(2.3—22.8 m2/kg)也一致[25]。这说明我们的调查具有系统性,真实的反映了温带地区植物SLA的变异特征。植物SLA在不同生活型间表现为草本>灌木>乔木。草本植物和灌木处于整个群落的中下层,受上部叶片的遮光影响[26],为适应这种弱光环境,增加对光能、CO2的吸收,植物增大自身SLA[27]。而处于顶层的乔木,受到强光的辐射,水分蒸发变强,为了减少光合辐射的伤害,往往采取减小SLA的方法来降低光合速率,而把更多的物质投入到增强叶片细胞壁和维管组织等结构建设上来[28—30],使自身得到保护,延长叶寿命[31]。
温带森林中所有物种、乔木、灌木和草本植物SLA沿纬度梯度的变化格局一致表现为:随纬度的升高,SLA呈现二项式的变化趋势。这与已有的物种水平上关于叶片SLA纬度格局的研究结论有所差异[24, 32]。这可能是因为本研究对研究区域内的物种进行了系统性的调查,所得结果更能准确表征该生态系统内植物SLA的特征,对植物SLA的纬度格局有更高的解释度。同时说明了少数优势种或常见种个体水平的性状值并不能代表该性状在整个森林内植物SLA的真实值。随纬度梯度升高,由暖温带到中温带,大气温度逐渐降低、降水量和土壤碳、氮元素含量明显增加(表1),良好的环境条件促使中高纬度地区倾向于选择较高SLA的植物。随纬度继续升高,由中温带向寒温带过渡,针叶林逐渐成为当地的优势树种,而针叶树种通常叶面积较小,SLA变小。大气温度和降水量明显降低,土壤元素含量下降,植物被迫减小叶片面积以降低体内水分的散失,同时通过自身调节将更多的物质分配到叶片中,增加叶片厚度,抵御寒冷,同时植物细胞体积变小,SLA相对减小[33]。
3.2 不同森林类型生活型植物SLA的差异
对于中国温带典型森林,不同森林类型间植物SLA差异显著(图4)。本研究结果显示,温带森林植物SLA在不同森林类型间表现为温带红松针阔混交林>暖温带落叶阔叶栎林>寒温带兴安落叶松林,这与纬度格局变化所伴随的环境、资源的改变密切相关。暖温带落叶阔叶栎林地区,生物多样性丰富,物种间对于资源的竞争激烈,物种都偏向相对保守的生存策略,因此SLA较小。温带红松针阔叶混交林区域,种内竞争激烈,淘汰率高,使其林下树种隔离程度高,生态位重叠低,林内资源得到充分利用,竞争压力小,SLA相对较高[34]。位于高纬度地区的寒温带兴安落叶松林,其所处环境资源贫瘠,温湿度低,植物在恶劣环境下,往往将叶片内大部分物质用于储存和防御或增加叶肉细胞密度,常形成厚度大而面积小的叶片,因而SLA较小。
不同森林类型间不同树种的植物SLA表现为落叶>常绿、阔叶>针叶。这与前人的研究结论基本一致,例如Wang等[24]对中国东部南北样带的研究结果表明,不同类型的植物SLA值表现为:阔叶>针叶;落叶>常绿,Wright等[13]的结论也与本研究结论一致。常绿树种往往比落叶树种叶厚度大,这是因为叶肉细胞密度大的缘故[35],进而影响CO2在叶肉细胞中的扩散,导致CO2同化速率降低,光合作用变弱[36],产生较小的SLA。在物质分配方面,常绿树种通常将物质一部分用于根、茎、枝等器官以及保卫细胞的构建,另一部分用于增加叶片硬度和厚度,以抵御寒冷干燥的环境和防御虫食,而落叶树种通常将干物质更多地投入到植物本身和叶面积的增长,因此落叶树种SLA较常绿树种高[12]。针叶树种生长在纬度较高的高寒地区,需要通过降低SLA来抵御不良环境,其自身叶面积也远小于阔叶树种,因而SLA很低。
3.3 不同环境条件植物SLA的适应对策
降水、温度、辐射和土壤养分等环境要素都会对植物SLA的大小产生影响(图6),Poorter等[15]和Wright等[13]都证明了这一点。本研究显示植物SLA随着降水的增加呈线性增加的趋势,可能是因为在降水较少时,植物受到水分胁迫,为减少水分的散失,它们会通过关闭气孔、减小SLA、增加表皮细胞特别是栅栏组织的厚度使细胞变得紧凑,以减少水分向外扩散[37—38]。而降雨量充沛的地区,生长条件适宜,环境的选择倾向于具有较高的SLA的植物出现。SLA随温度升高先增加后降低,在低温条件下,由于物种自身的适应性差异会导致适应能力弱的植物体内碳循环失调[39],使得环境不再适宜其生存;而适应能力强的植物通过自身调节增加体内可溶性糖含量,提高抗冻能力以适应高寒环境,同时由于需要碳水化合物维持其基本生命活动[40—41],植物将碳更多的用于提升防御和生存能力,叶干物质量增加,SLA降低。在高温强辐射条件下,植物会通过关闭气孔、减小SLA来降低光合速率和水分的蒸散[29]。在低光能条件下,植物可能会为了避免食草动物的取食,增加叶片表皮毛密度、叶肉细胞密度和细胞壁的厚度,以达到增强叶片硬度的目的[42],叶干重增加,SLA降低。温度、辐射适宜的条件,适合植物生长,SLA相对较大。因此,植物SLA随光照强度同样表现为先增加后降低。土壤养分高的地方,植物SLA较高[25, 43],而养分限制会影响植物叶面积,造成SLA过小[15]。研究表明,SLA与土壤总氮含量呈正相关关系[43],这与本研究的结论一致。土壤养分会直接影响植物体内养分含量,叶碳含量和叶氮含量的提高促使植物的物质生产和光合作用加强[44—45],SLA变大。以往研究表明,SLA与土壤磷元素含量呈正相关关系[46],与土壤酸碱性之间表现为负相关。这在本研究中并未体现。这可能是因为本研究中研究地点所处地区土壤磷含量和土壤酸碱性差异并不是很明显,所以并未显示出很强的相关性。
4 结论
本研究探讨了温带森林植物SLA的特点。经分析发现,植物SLA随空间格局的变化呈二项式变化规律,植物SLA在三种典型森林间具有明显的差异,表现为温带红松针阔混交林>暖温带落叶阔叶栎林>寒温带兴安落叶松林。对于SLA的影响因素而言,气候是调控植物SLA的主要因素,特别是随着年均温度和年均降水量的变化,植物SLA会产生极显著变化。本研究揭示了中国温带森林SLA的空间变异格局及其主要影响因素,可以为将来全球尺度下SLA的研究提供数据支撑,为预测全球气候变化背景下植物性状变异提供理论基础,但对于SLA在群落水平上的概况、不同植被类型、不同植物功能群中的变异大小以及其与生态系统功能的关系的研究尚有所欠缺,未来应对此方面有所加强。
猜你喜欢
疯狂英语·新悦读(2022年7期)2022-11-23
北京大学学报(自然科学版)(2022年4期)2022-08-18
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
林业科技(2020年3期)2021-01-21
意林·少年版(2020年17期)2020-10-12
发明与创新·中学生(2020年4期)2020-04-17
新疆农业科学(2020年1期)2020-02-14
现代园艺(2018年2期)2018-03-15
中国麻业科学(2015年5期)2015-12-28
安徽农业科学(2015年30期)2015-12-25
