
益蝽5龄若虫对不同龄期粘虫幼虫的捕食作用
2022-03-05 02:03申修贤田太安刘健锋于晓飞董祥立李治模杨茂发
中国农学通报 2022年3期
申修贤,田太安,刘健锋,于晓飞,董祥立,李治模,杨茂发,
(1贵州大学昆虫研究所,贵州省山地农业病虫害重点实验室,贵阳 550025;2贵州大学烟草学院,贵阳 550025;3贵州省烟草公司遵义市公司凤冈县分公司,贵州 遵义 564200)
0 引言
粘虫Mythimna seperata(Walker)又称行军虫,属鳞翅目Lepidoptera 夜蛾科Noctuidae,无滞育现象,是一种典型远距离迁飞世界性害虫。粘虫在中国分布广泛,除新疆外都有报道其为害,严重时暴发成灾[1]。粘虫为害涉及作物种类多、爆发性明显、为害程度大和为害历史长等特点,对中国粮食生产造成严重损失。粘虫的为害报道始于公元482 年,在1950—1989 年间就有17 年暴发成灾,21 世纪以来,粘虫的局部为害一直未曾间断,在2012 年全国范围内大面积发生危害,其主要为害玉米、小麦、水稻等粮食作物,取食其叶片,严重时将整株取食光,造成减产甚至绝产[2-3]。至今化学农药仍是生产上对粘虫的主要防治手段,虽然粘虫的迁飞习性导致不同种群间基因交流频繁,对常用化学农药抗药性不明显,但大面积长期使用对生态环境影响较大,如对天敌昆虫、传粉昆虫等益虫造成损害,对物种多样性造成影响[4]。随着人们对食品安全和生态环境的重视,迫切需要安全、环保的方法对害虫进行防控。
天敌昆虫的应用作为生物防治重要组成部分。具有持续性、成本低和对环境友好等优点,是当前害虫的综合防治技术重要策略和发展趋势[5]。益蝽Picromerus lewisi(Scott)属半翅目Hemiptera,蝽科Pentatomidae,益蝽亚科Asopinae,是中国常见的一种天敌昆虫,在俄罗斯、朝鲜和日本等区域也广泛分布[6]。据报道,益蝽能捕食斜纹夜蛾Spodoptera litura、草地贪夜蛾Spodoptera frugiperda、小菜蛾Plutella xylostella和烟青虫Helicoverpa armigera等多种重要农业害虫[7]。现阶段对益蝽已能实现规模化人工饲养[8],其具有较大的生防潜能。唐艺婷等[9]研究了不同龄期益蝽对3 龄粘虫捕食能力,但对天敌自身种内干扰、分摊竞争强度以及不同龄期粘虫的捕食能力鲜见报道。因此,本研究选取益蝽5 龄若虫对不同龄期粘虫幼虫进行捕食能力研究,评估益蝽对粘虫的综合控害能力,为应用益蝽防治粘虫提供理论基础。
1 材料与方法
1.1 供试虫源
益蝽和粘虫由贵州省烟草公司遵义市公司天敌繁育中心提供,粘虫使用人工饲料进行饲养,以5龄粘虫幼虫作为益蝽的食物。两者均饲养于大型人工气候室内,环境条件为温度(25±2)℃、光周期14 L:10 D、相对湿度75%±5%。
1.2 实验条件
益蝽对粘虫的捕食能力实验在直径15 cm、高2 cm的培养皿内完成。培养皿置于人工气候室内,环境条件为温度(25±2)℃、光周期14 L:10 D、相对湿度75%±5%。
1.3 实验方法
1.3.1 捕食功能反应和搜寻效应实验 实验前将益蝽5龄若虫单头置于有4 cm2湿润脱脂棉15 cm×2 cm的培养皿中饥饿24 h 处理,24 h 后放入不同密度的3 龄、4龄、5 龄和6 龄粘虫供其捕食,3 龄粘虫密度梯度为5、10、15、20、25、30 头/皿;4 龄粘虫密度梯度为4、8、12、16、20、24 头/皿;5 龄粘虫密度梯度为3、6、9、12、15、18 头/皿;6 龄粘虫密度梯度为2、4、6、8、10、12 头/皿。每个密度处理均设置7 次重复,24 h 后观察每个培养皿中存活的粘虫数量。
1.3.2 自身密度干扰实验 与以上捕食功能反应实验相同方法。首先对益蝽5 龄若虫进行饥饿24 h 处理,再向装有饥饿处理益蝽5 龄若虫的15 cm×2 cm 培养皿中放入60 头3 龄粘虫幼虫,益蝽5 龄若虫密度设置为1、2、3、4、5、6头/皿,每个密度处理重复7次,24 h后观察粘虫的存活情况。
1.4 数据统计与分析
使用Excel 2010和Origin 2017对数据进行拟合与作图,利用HollingⅡ反应模型进行拟合。圆盘方程[10]、拟合搜寻效应方程[11]和Hassell 的干扰模型[12]见公式(1)、(2)、(3)。

其中Na为捕食粘虫的数量,a为益蝽对粘虫的瞬时攻击率,N为粘虫不同龄期幼虫的密度,Tr是实验的总时间为1 天,Th为处理时间(益蝽捕食1 头粘虫所需时间);A为每头益蝽捕食粘虫幼虫的数量,P为益蝽密度,b为竞争参数。
2 结果与分析
2.1 益蝽5龄若虫对不同龄期粘虫幼虫的捕食功能反应
5龄益蝽若虫捕食不同龄期粘虫幼虫的功能反应使用HollingⅡ进行拟合(R2≥0.949)(表1)。通过拟合方程参数可以看出,益蝽捕食3 龄粘虫的瞬时攻击率(1.303)、日最大捕食量(24.390)和控害效能(31.780)均较捕食其他龄期要高,且相差较大,而对4龄、5龄和6龄捕食功能反应参数之间相差不大。5龄益蝽若虫对粘虫幼虫的处理时间为6龄(0.271)>5龄(0.165)>4龄(0.142)>3龄(0.041);随着不同龄期粘虫幼虫密度的增加,益蝽5 龄若虫对粘虫的平均捕食量逐渐趋于平缓(图1)。

图1 益蝽5龄若虫对不同龄期粘虫的捕食功能反应
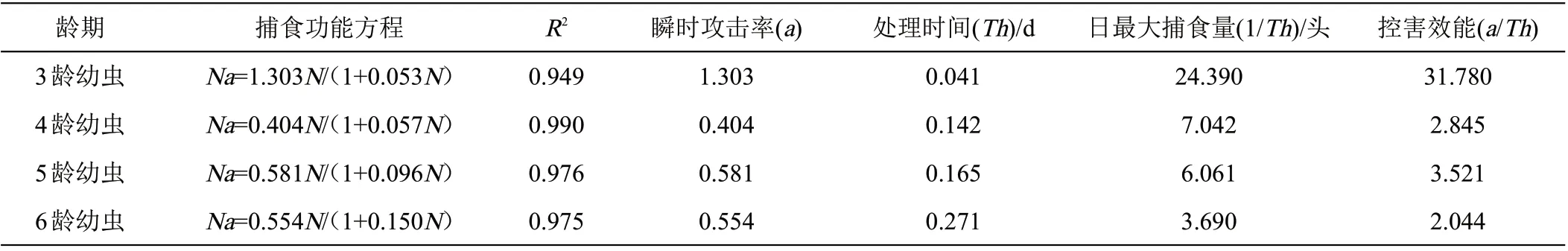
表1 益蝽5龄若虫捕对粘虫的捕食功能反应参数
2.2 益蝽5龄若虫对不同龄期粘虫幼虫的搜寻效应
根据S=aTr/(1+aThN)公式计算出益蝽5龄若虫对不同龄期粘虫幼虫的搜寻效应(图2)。益蝽5 龄若虫的搜寻效应随不同龄期粘虫的密度增加而降低。由搜寻效应曲线可以看出,5龄益蝽对3龄粘虫搜寻效率最高,对4龄粘虫搜寻效率最低。

图2 益蝽5龄若虫对不同龄期粘虫的搜寻效应
2.3 益蝽5 龄若虫对粘虫3 龄幼虫捕食的种内干扰反应
益蝽5龄若虫捕食粘虫3龄幼虫存在自身密度干扰作用(图3)。通过Hassell的干扰模型拟合的方程为A=15.985P(-0.720)(R2=0.970),最大捕食量15.985 头,种内干扰系数m为0.720。随着益蝽密度的增加,单头益蝽对3龄粘虫的平均捕食量逐渐降低。表2可知,其益蝽5龄若虫种内分摊竞争强度随着自身密度的对数值的增长而增长,分摊竞争效应方程I=0.7071logP-0.6679(R2=0.9876)。

表2 益蝽5龄若虫捕食3龄粘虫的种内干扰作用

图3 益蝽5龄若虫对粘虫3龄幼虫的实际捕食量
3 讨论与结论
天敌的捕食功能反应可用于评价天敌对害虫控害效能,其中天敌对害虫的瞬间攻击率a与处理时间Th之比可作为衡量参数之一,比值越大,表明天敌的实际捕食能力越强,其对害虫的控制效果越好[13],本研究利用益蝽对粘虫的捕食作用,结果表明,益蝽5龄若虫能捕食粘虫3 龄、4 龄、5 龄和6 龄幼虫,且与躅蝽Arma chinensis[14]、叉角厉蝽Eocanthecona furcellata[15]、斑腹刺益蝽Podisus maculiventris[16]等捕食性蝽捕食鳞翅目昆虫幼虫一样,捕食关系符合HollingⅡ反应模型。与捕食其他龄期的粘虫幼虫相比,益蝽5龄若虫对粘虫3龄幼虫捕食作用最强,其瞬时攻击率、最大捕食量和控害效能都优于其他3 个相对较大的龄期,这与益蝽对不同龄期草地贪夜蛾捕食作用结果相似[17]。
研究捕食者—猎物的关系模型是天敌昆虫应用研究的重要组成部分,对优化生防策略具有重要意义,捕食者对猎物的攻击过程中存在搜寻效应,一般情况下,捕食者的搜寻效应会随猎物的密度增加而降低[18]。本研究结果表明益蝽5龄若虫的搜寻效应随粘虫幼虫密度的增加而降低。除猎物密度会影响捕食作用外,在一定空间范围下,捕食者自身密度对其捕食作用也有较大的影响[19]。天敌的种内竞争分为争夺竞争和分摊竞争2种形式,而益蝽属于分摊竞争,所有益蝽对粘虫都有相同的机会去接近粘虫。当粘虫数量一定时,益蝽的分摊竞争强度与种内干扰作用均均随着自身密度的增加而增加。这与众多捕食性昆虫的干扰反应一致[20-22]。
因此,通过室内试验研究结果表明,在粘虫3龄幼虫高发期前释放益蝽5 龄若虫,能够在一定程度上防治粘虫的为害,达到很好的防治效果,为田间利用益蝽防治粘虫提供了一定理论依据。本研究未进行田间试验,在后续中将继续进行田间试验,充分结合田间实际环境条件、猎物丰富度、种间竞争等生物与非生物因素等,这样能更全面地评价益蝽对田间粘虫的防治效果。
猜你喜欢
今日农业(2022年13期)2022-09-15
农业工程学报(2022年10期)2022-08-22
天津农业科学(2022年5期)2022-05-31
南方农业学报(2021年11期)2021-03-05
科学之谜(2020年1期)2020-05-18
江苏农业科学(2017年12期)2017-09-16
现代农业研究(2017年4期)2017-05-23
天津农业科学(2015年7期)2015-09-07
天津农业科学(2015年9期)2015-09-02
建筑科学与工程学报(2014年4期)2015-02-03
