
牦牛酥油鞘磷脂对小鼠脂质代谢紊乱和肝脏组织炎症的调节作用
2022-03-03 08:33孙万成罗毅皓
食品科学 2022年3期
罗 鑫,孙万成,罗毅皓*
(青海大学农牧学院,青海 西宁 810016)
牦牛酥油是似黄油的一种乳制品,是从牦牛奶中提炼出的脂肪。产于夏秋季的牦牛酥油色泽鲜黄、味道香甜、口感极佳,产于冬季的牦牛酥油则呈淡黄色。酥油含多种维生素,与普通奶油相比营养价值颇高,其硬脂酸含量更高、棕榈酸含量更低,反式油酸和共轭亚油酸分别是普通奶油的近8 倍和2 倍[1]。
鞘磷脂是含鞘氨醇或二氢鞘氨醇的磷脂,其分子不含甘油,是一分子脂肪酸以酰胺键与鞘氨醇的氨基相连形成的,是细胞膜的重要组成成分[2],也是一种膳食成分,在鸡蛋、牛奶、肉类和鱼类等食物中最为丰富[3]。近年来有研究表明,鞘磷脂与多种疾病的发生和治疗有密切关系[4],鞘磷脂主要位于细胞膜、脂蛋白和其他富含脂类的组织结构上,对维持细胞膜结构尤其是细胞膜的微控功能十分重要,还与人体内的脂肪酸、胆固醇所引起的一些疾病有着密切的关系[5]。膳食中的鞘磷脂可通过调节结肠癌发生早期表达的蛋白质,抑制结肠癌、肝癌细胞增殖,使得鞘磷脂在结肠癌和肝癌的发生过程中可能成为一种潜在的治疗候选药物[6-8];膳食鞘磷脂可显著上调缺镁饮食导致的特异性皮炎模型表皮中神经酰胺合成酶的表达,抑制缺镁饮食无毛小鼠皮肤炎症的发生[9-10];研究表明,与对照组相比,饲喂含鞘磷脂饲料的大鼠血浆中胆固醇含量减少了30%[11-13]。目前,高脂饮食习惯正逐渐影响着人类的健康,长期高脂饮食、体内营养过剩易引起高脂血症,患者长期处于炎症的状态,机体的脂代谢平衡也会发生改变。鞘磷脂具有一定的抗炎症功能[14]和调节脂质代谢[15]的作用。Norris等[11]研究发现牛奶鞘磷脂能降低高脂肪饮食诱导的C57BL/6J小鼠的血清血脂和肝脏甘油三酯。Chung等[12]发现当将鸡蛋鞘磷脂添加到高脂肪的饮食中时,其会以剂量依赖地方式有效降低肝脏胆固醇和甘油三酯的水平,并且对饮食诱导的肝脏脂肪组织炎症具有潜在的预防作用。依帕尔古丽·阿不拉等[13]研究了新疆拜城油鸡中的鞘磷脂提取物对高脂血症小鼠血脂及肝脏的影响,发现拜城油鸡鞘磷脂提取物可显著降低高脂血症小鼠的血脂水平,保护肝细胞、改善肝细胞脂肪变性。
不同食物中鞘磷脂的含量不同,某些水果和蔬菜中鞘磷脂含量不到100 μmol/kg,在鸡蛋、奶制品和部分植物种子中可达2 000 μmol/kg 以上[16],诸多学者通过对市售婴儿配方乳粉中磷脂的含量进行分析,发现磷脂在婴儿配方奶粉中的含量为15~607 mg/100 g,其中鞘磷脂为12~125 mg/100 g,磷脂在牛乳中的质量浓度为4.60~49.10 mg/100 mL,磷脂中鞘磷脂相对含量为17%~41.10%,磷脂在母乳中的质量浓度为13.30~40.60 mg/100 mL,鞘磷脂占磷脂的24.0%~45.9%[17],而牦牛酥油中鞘磷脂含量可达1 918 mg/100 g,通过分析牦牛酥油中鞘磷脂的脂肪酸组成及含量,发现其饱和脂肪酸占脂肪酸总量的73.73%~75.89%,不饱和脂肪酸占24.12%~26.27%,与牦牛酥油和牦牛酥油磷脂中超长链脂肪酸含量相比,牦牛酥油鞘磷脂中超长链脂肪酸(占23.15%)的含量具有极显著性差异[18-19]。因此,本实验分析牦牛酥油鞘磷脂对高脂饮食引起的小鼠脂代谢紊乱及肝脏组织炎症的作用,初步探讨鞘磷脂调节脂代谢及改善组织炎症的机制,为更好地开发和利用鞘磷脂提供理论基础。
1 材料与方法
1.1 动物、材料与试剂
C57BL/6J小鼠,SPF级,雄性,4 周龄,体质量约13.00 g,由兰州兽医研究所提供,实验动物生产许可证号为SCXK(甘)2015-0001。
牦牛酥油鞘磷脂 本实验室自制;总胆固醇(total cholesterol,TC)、甘油三酯(triglycerides,TG)、高密度脂蛋白胆固醇(high density lipoprotein cholesterol,HDL-C)、低密度脂蛋白胆固醇(low density lipoprotein cholesterol,LDL-C)、空腹血糖、丙氨酸转氨酶(alanine aminotransferase,ALT)和天冬氨酸转氨酶(aspartate transaminase,AST)测定试剂盒 南京建成生物工程研究所;脂肪酸合成酶(fatty acid synthase,FAS)、硬脂酰辅酶A去饱和酶1(stearoyl-CoA desaturase 1,SCD1)、肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素(interleukin,IL)-6等基因 上海生物有限公司合成;Total RNA试剂盒美国Omega Bio-Tek公司;Evo M-MLV反转录试剂盒、SYBR Green Pro Taq HS预混型qPCR试剂盒 艾科瑞生物技术有限公司。小鼠低脂饲料(TP23302,脂肪质量分数为10%)和高脂饲料(TP23300,脂肪质量分数为60%) 江苏南通特洛菲饲料科技有限公司。
1.2 仪器与设备
MμLtiskan FC酶标仪 美国Thermo Fisher公司;YM立式压力蒸汽灭菌器 上海三申医疗器械有限公司;CFX96荧光定量仪 美国 BIO-RAD公司;2-16KL冷冻离心机 德国Sigma公司;NanoDrop 2000紫外-可见分光光度计 美国赛默飞世尔公司。
1.3 方法
1.3.1 牦牛酥油鞘磷脂的制备
牦牛酥油鞘磷脂参考本课题组已发表的方法制得[18]。参照GB/T 35867—2018《粮油检验 卵磷脂中磷脂含量的测定 高效液相色谱蒸发光散射检测法》测得牦牛酥油中鞘磷脂的含量为1 918 mg/100 g,提取的鞘磷脂纯度为78%。鞘磷脂标准溶液和牦牛酥油鞘磷脂样品的高效液相色谱图分别见图1、2。
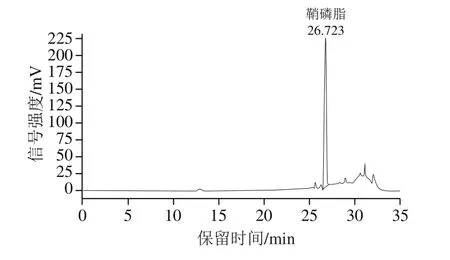
图1 鞘磷脂标准溶液高效液相色谱图Fig.1 High performance liquid chromatogram of sphingomyelin standard solution
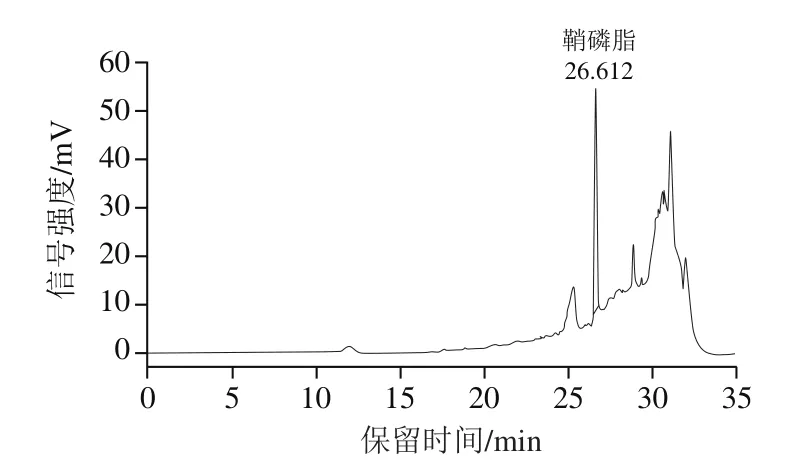
图2 牦牛酥油鞘磷脂样品溶液高效液相色谱图Fig.2 High performance liquid chromatogram of yak ghee sphingomyelin solution
1.3.2 动物分组及饲喂
将30 只雄性C57BL/6J小鼠按体质量随机分为5 组,每组6 只,并适应性饲喂7 d后进行实验,小鼠分组如下:低脂组(LF组)、高脂组(HF组)、鞘磷脂高剂量组(HF+HSM组,鞘磷脂添加量为1.20 g/100 g)、鞘磷脂中剂量组(HF+MSM组,鞘磷脂添加量为0.60 g/100 g)和鞘磷脂低剂量组(HF+LSM组,鞘磷脂添加量为0.30 g/100 g),根据Norris等[21]的方法确定鞘磷脂添加量,按照不同剂量直接将鞘磷脂拌入到饲料中,观察确保每次饲喂的含鞘磷脂饲料均能被小鼠食用完。除低脂组小鼠饲喂低脂饲料外,其他组小鼠均给予高脂饲料或高脂饲料+鞘磷脂进行饲喂,所有小鼠均被安置在恒定的温度、湿度条件和12 h的光暗循环下。自由摄食和饮水,饲料每日更新并测定摄食量,按公式(1)计算食物利用率。每周测量体质量。

鞘磷脂饲喂4 周后,小鼠禁食10 h,麻醉摘眼球取血后处死,处死前进行称质量,麻醉取血后打开小鼠腹腔和胸腔,立即分离肝脏、心脏、脾、肾、腹部脂肪和肾周脂肪,测定质量,然后取腹部脂肪和100 mg肝脏组织进行组织病理学观察,剩余组织于-80 ℃贮存待用。分别按公式(2)、(3)计算脏器指数和脏器脂肪指数。

1.3.3 血清和肝脏生化指标检测
用1.5 mL的无菌EP管收集小鼠眼球血,室温静置1 h后,于4 ℃、3 000 r/min 离心10 min,吸取上层血清,分装保存于-80 ℃,参照试剂盒说明书测定血清中TC、TG、HDL-C、LDL-C和空腹血糖的浓度。再根据试剂盒的方法测定肝脏组织中ALT、AST的活力,结果以蛋白质量计。
1.3.4 肝脏和脂肪组织病理学分析
收集肝脏和腹部脂肪组织,经体积分数4%多聚甲醛固定后,石蜡切片用苏木精-伊红(hematoxylin-eosin,HE)染色,在光学显微镜下观察组织病理学变化,并进行图像采集。
1.3.5 实时定量聚合酶链式反应分析
使用Total RNA试剂盒按照说明对5 个组小鼠的肝脏组织RNA进行提取,并利用NanoDrop 2000紫外-可见分光光度计对提取的RNA浓度和纯度进行测定,OD260nm/280nm值在1.80~2.10之间。根据Evo M-MLV反转录试剂盒的方法合成cDNA第一条链。根据SYBR Green Pro Taq HS预混型qPCR试剂盒的方法对18 个基因进行实时定量聚合酶链式反应分析,脂质代谢和炎症因子的引物序列见表1。所有基因均以GAPDH为参考基因,采用比较阈值法(2-ΔΔCt)进行相对定量,计算小鼠肝脏组织脂代谢和炎症因子mRNA相对表达量。注:ACC-α.乙酰辅酶A羧化酶α(acetyl-CoA carboxylase alpha);FADS1.脂肪酸去饱和酶1(fatty acid desaturase 1);FADS2.脂肪酸去饱和酶2(fatty acid desaturase 2);GPAT4.甘油-3-磷酸酰基转移酶4(glycerol-3-phosphate acyltransferases 4);GPAM.3-磷酸甘油酰基转移酶(glycerol-3-phosphate acyltransferase);THRSP.甲状腺激素应答蛋白(thyroid hormone responsive protein);HMGCR.3-羟基-3-甲基戊二酸辅酶A还原酶(3-hydroxy-3-methylglutaric acid coenzyme A reductase);CPT1.肉碱棕榈酰转移酶1(carnitine palmitoyl transferase 1);FABP1.肝型脂肪酸结合蛋白(hepatic fatty acid binding protein);A-FABP.脂肪细胞脂肪酸结合蛋白(adipocyte fatty acid-binding protein);IL-1Ra.白介素1受体拮抗剂(interleukin-1 receptor antagonist);IFN-γ.干扰素γ(interferon-γ)。

表1 引物序列信息Table 1 Primers sequences

续表1
1.4 数据处理与分析
所有数据以平均值±标准差表示,采用SPSS 16.0软件进行单因素方差分析,采用邓肯检验进行显著性分析,P<0.05表示差异显著。
2 结果与分析
2.1 鞘磷脂对高脂饮食小鼠体质量和总摄食量的影响
由表2和图3可知,与高脂组相比,牦牛酥油鞘磷脂组小鼠(除鞘磷脂低剂量组外)的最终体质量显著降低(P<0.05)。相对于低脂组,高脂组小鼠的摄食量显著降低(P<0.05),而其体质量增加量显著升高(P<0.05),说明高脂饮食能够诱导小鼠肥胖的发生;与高脂组相比,鞘磷脂组小鼠的总摄食量与之相近,但体质量增加量有所降低,尤其是高剂量组具有显著差异(P<0.05)。相对于高脂组,低脂组的食物利用率显著降低(P<0.05),高剂量鞘磷脂组虽有较低的食物利用率,但差异不显著(P>0.05)。上述数据表明牦牛酥油鞘磷脂具有降低由肥胖饮食引发体质量增加的作用。
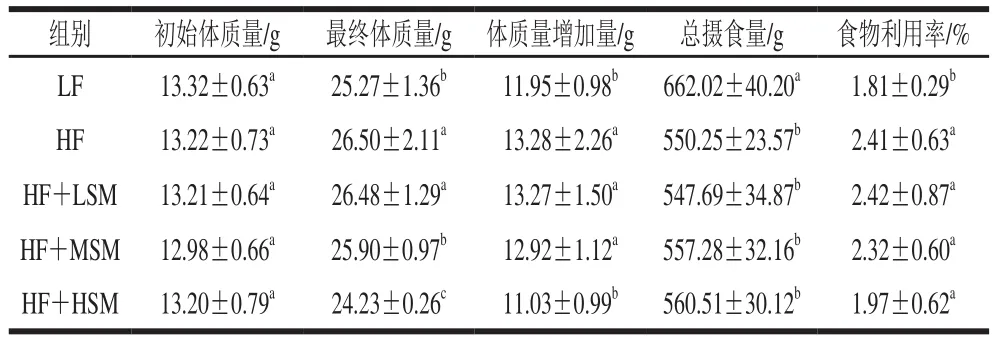
表2 各组小鼠的体质量增加量、总摄食量和食物利用率(n=6)Table 2 Body mass gain, total feed intake, and utilization rate of mice in each group (n = 6)
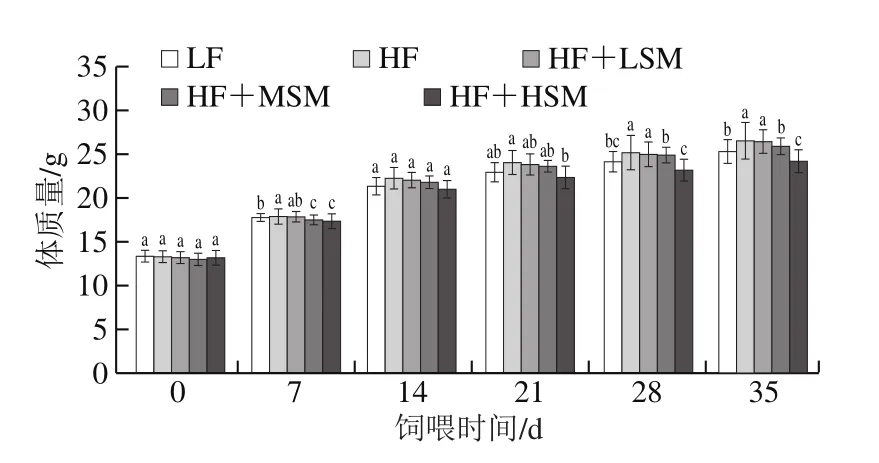
图3 牦牛酥油鞘磷脂对小鼠体质量的影响Fig.3 Effect of yak ghee sphingomyelin on body mass of mice
2.2 鞘磷脂对高脂饮食小鼠脏器指数、腹部和肾周脂肪指数的影响
由表3可知,与低脂组相比,高脂组小鼠心脏、肾脏和脾脏指数均有所升高;肝脏指数显著增高(P<0.05),从39.90 mg/g增加至44.15 mg/g。与高脂组相比,鞘磷脂各组小鼠的脾脏、心脏和肾脏指数较低,但无显著差异;高剂量鞘磷脂组的肝脏指数显著下降了8.54%(P<0.05)。此外,与低脂组相比,高脂组小鼠腹部脂肪和肾周脂肪指数显著升高(P<0.05),鞘磷脂低、中、高剂量组中小鼠腹部脂肪指数较高脂组分别降低了28.54%、29.83%和34.37%,肾周围脂肪指数也显著降低(P<0.05)。以上研究表明牦牛酥油鞘磷脂对小鼠的心脏、肾脏和脾脏无明显影响,且能够改善高脂饮食引起的小鼠肝脏指数变化和缓解腹部脂肪及肾周脂肪的积聚。
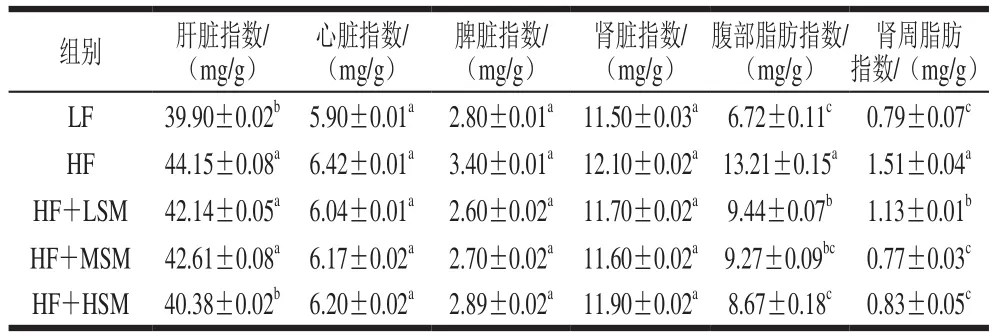
表3 各组小鼠的脏器指数、腹部脂肪指数和肾周脂肪指数(n=6)Table 3 Visceral organ indexes, abdominal fat index, and perirenal fat index of mice in each group (n = 6)
2.3 鞘磷脂对高脂饮食小鼠血清血脂水平的影响
高脂饮食引起的血液中TC、TG和LDL-C水平升高会导致动脉粥样硬化等疾病的发生,本研究对小鼠血清血脂水平的分析评估了牦牛酥油鞘磷脂的脂代谢作用。由图4可知,与低脂组相比,高脂组小鼠TC、TG和LDL-C和血糖水平显著升高,HDL-C水平显著下降(P<0.05);相对于高脂组,鞘磷脂组中TC、TG、LDL-C和血糖水平显著下降(P<0.05),高剂量鞘磷脂组TC、TG和LDL-C和血糖水平分别下降了26.26%、24.70%、38.96%和42.16%。Zhang Ping[20]、Norris[21]等发现牛奶和鸡蛋鞘磷脂可通过抑制内源性胆固醇的吸收降低血液中胆固醇的含量,从而改善高脂饮食引起的血脂水平增加。牦牛酥油鞘磷脂对体内血脂具有代谢作用,其降血脂的潜在机理表现为可以通过抑制胆固醇的吸收防止胆固醇沉积,有效地维持血液中胆固醇水平的平衡;此外,牦牛酥油鞘磷脂中不饱和脂肪酸含量较高,不饱和脂肪酸具有调节血脂、降低血液中胆固醇含量的作用。
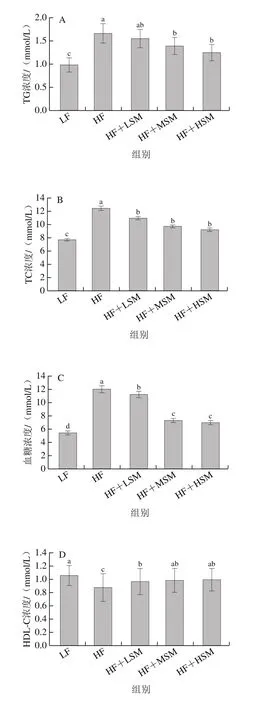
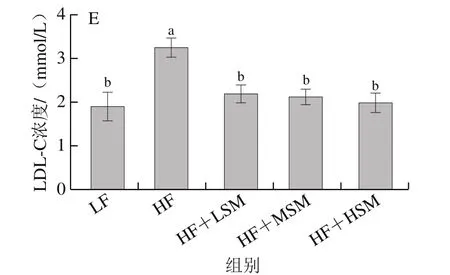
图4 牦牛酥油鞘磷脂对高脂饮食小鼠血清血脂水平的影响Fig.4 Effect of yak ghee sphingomyelin on serum lipid levels in HFD-fed mice
2.4 鞘磷脂对高脂饮食小鼠肝脏AST和ALT活力的影响
ALT和AST活力为评价肝功能的重要指标,肝脏组织受损时,细胞膜的通透性增加,肝细胞浆内ALT和AST释放到血液中,会引起肝脏和血清中转氨酶水平升高[22]。从图5可以看出,与低脂组相比,高脂饮食引起了肝脏ALT和AST活力的显著升高(P<0.05),低脂组中ALT和AST的活力分别为11.16 U/g和11.76 U/g;高脂组中ALT和AST的活力分别为24.37 U/g和15.76 U/g;与高脂组相比,鞘磷脂高、中、低剂量组小鼠肝脏ALT和AST活力均显著降低(P<0.05),ALT的活力分别为15.99、18.63、21.58 U/g,AST的活力分别为12.91、14.24、14.32 U/g。Norris[21]等发现与低脂饮食相比,高脂饮食能够使血清中ALT、AST活力分别显著增加43%和30%(P<0.05),鸡蛋鞘磷脂和牛奶鞘磷脂均能降低ALT(分别降低了8.30%和8%)和AST(分别降低了5%和10%)的活力。
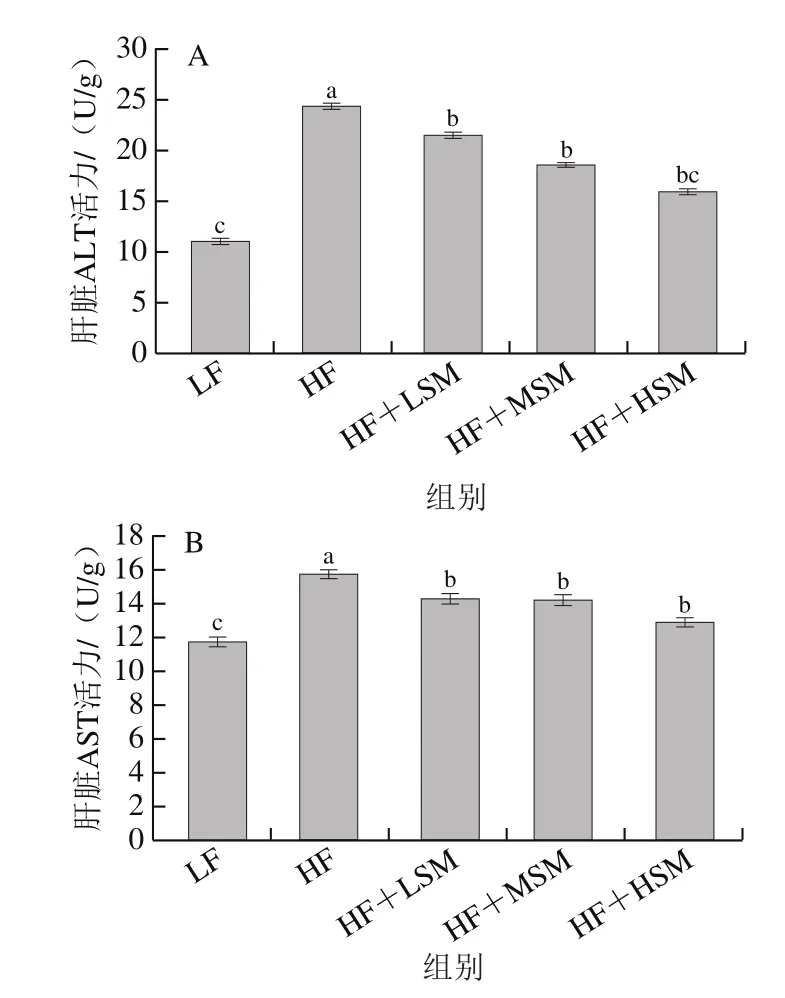
图5 牦牛酥油鞘磷脂对高脂饮食小鼠肝脏ALT(A)、AST(B)活力的影响Fig.5 Effect of yak ghee sphingomyelin on liver alanine aminotransferase (A)and aspartate transaminase (B) activity in HFD-fed mice
2.5 鞘磷脂对高脂饮食小鼠肝脏和腹部脂肪组织病理学的影响
为了研究饲喂鞘磷脂后对小鼠肝脏及腹部脂肪组织病理变化的影响,进一步对肝脏和腹部脂肪组织进行HE染色。由图6、7可知,低脂组和高脂组中,小鼠肝脏组织结构正常完整,肝细胞沿中央静脉呈放射状排列,未见变性、坏死,高脂组偶见轻微肝细胞空泡变;腹部脂肪组织的脂肪细胞形态结构正常,间质未见充血、水肿和炎细胞浸润。鞘磷脂组中小鼠肝脏组织结构正常完整,肝细胞沿中央静脉呈放射状排列,未见变性、坏死,肝窦未见扩张,间质未见充血、水肿和炎细胞浸润;腹部脂肪组织的脂肪细胞形态结构正常,间质未见充血、水肿和炎细胞浸润。染色结果表明,饲喂鞘磷脂后,可以减轻由高脂饮食引起的小鼠肝脏细胞空泡变,从而对肝脏起到一定的保护作用。
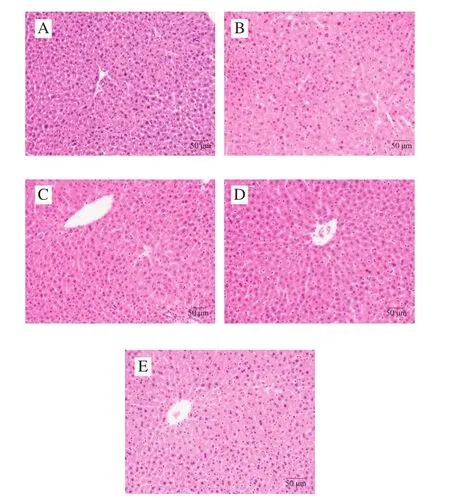
图6 牦牛酥油鞘磷脂对高脂饮食小鼠肝脏组织病理学的影响Fig.6 Effect of yak ghee sphingomyelin on liver histopathology in HFD-fed mice
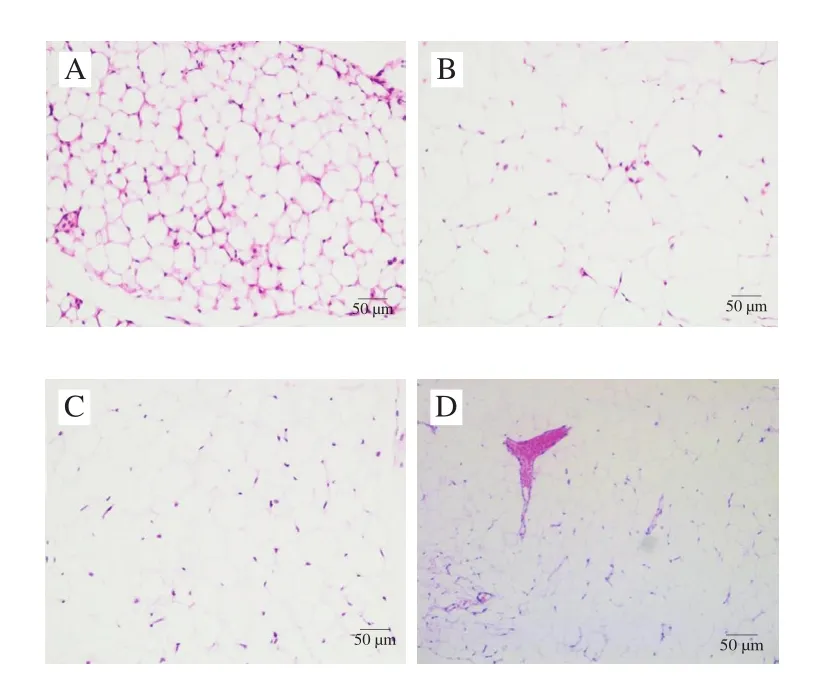
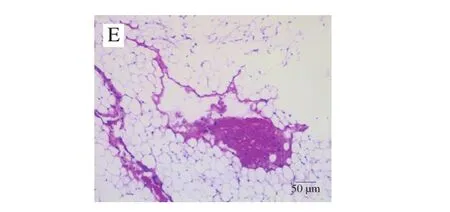
图7 牦牛酥油鞘磷脂对高脂饮食小鼠腹部脂肪组织病理学的影响(×200)Fig.7 Effect of yak ghee sphingomyelin on the histopathology of abdominal adipose tissues in HFD-fed mice (× 200)
2.6 鞘磷脂对高脂饮食小鼠肝脏脂代谢和炎症因子表达量的影响
2.6.1 对脂质代谢基因表达的影响
如图8所示,与高脂组相比,饲喂鞘磷脂后,小鼠肝脏组织中FAS、A-FABP、ACC-α、FADS2、THRSP、FADS1、GPAM、CPT1和GPAT4基因的表达受到抑制,其表达量显著降低(P<0.05);FABP1、HMGCR和SCD1基因的表达显著上调。
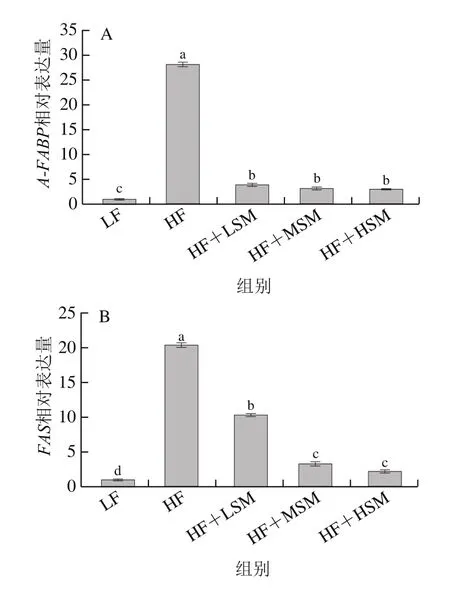
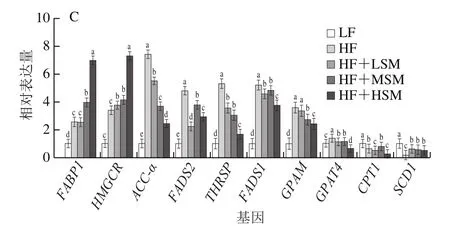
图8 牦牛酥油鞘磷脂对高脂饮食小鼠肝脏脂代谢基因表达量的影响Fig.8 Effect of yak ghee sphingomyelin on lipid metabolism gene expression in liver tissues of HFD-fed mice
FADS1和FADS2基因的表达在多不饱脂肪酸的合成中起关键作用,ACC-α是脂肪酸生物合成第一步的限速酶[23],GPAM定位于线粒体,GPAT4定位于内质网[24],是甘油三酯合成第一步反应的关键酶。THRSP基因参与甲状腺激素和碳水化合物诱导的脂肪生成过程[25],FAS的表达量随着THRSP表达增强而提高[26]。SCD1基因是单不饱和脂肪酸合成过程中的限速酶[27]。因此,牦牛酥油鞘磷脂调节脂质代谢的作用机制表现为:通过抑制磷酸甘油途径合成甘油三酯、减少甲状腺激素和碳水化合物诱导脂肪的合成、调节肝脏中脂类的转运和代谢从而减少肝脏和血清中脂质的积累。牦牛酥油鞘磷脂的降血脂机理与肝脏组织中脂质代谢相关酶的表达密切相关,鞘磷脂通过影响相关酶的表达量调节机体中的饱和及不饱和脂肪酸的含量,进而对血脂起调节作用。
2.6.2 对炎症因子基因表达的影响
促炎因子和抗炎因子在多种炎症反应中起重要作用[28]。高脂饮食不仅会引起血脂代谢紊乱,还会使机体长期处于慢性炎症状态[29]。高脂饲料饲喂小鼠会显著上调机体促炎症因子的表达[30],TNF-α和IL-6等促炎因子的释放,能够增强炎症反应,引起肝损伤[31]。因此,本实验利用荧光定量聚合酶链式反应检测炎症因子在肝脏中的表达水平,由图9可知,高脂组小鼠肝脏中促炎症因子IL-6、TNF-α、IL-1β和IFN-γ的表达量显著高于低脂组(P<0.05),鞘磷脂组小鼠肝脏中促炎症因子的表达量均显著低于高脂组(P<0.05)。与高脂组相比,鞘磷脂中、高剂量组中抗炎症因子IL-10的表达量显著升高(P<0.05),IL-1Ra和IL-4表达量随着鞘磷脂添加量的增加呈上升趋势,因此,牦牛酥油鞘磷脂改善炎症反应的作用机制可能与抑制TNF-α和IL-6等促炎因子的生成相关,但本研究中鞘磷脂对体内炎症因子的表达量影响无明显剂量对应关系,可能与本实验的设计相关,需要通过建立相应的炎症模型或增加饲喂时间及样本量进行验证。
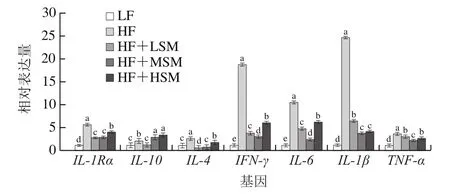
图9 牦牛酥油鞘磷脂对高脂饮食小鼠肝脏炎症因子基因表达量的影响Fig.9 Effect of yak ghee sphingomyelin on inflammatory gene expression in liver tissues of HFD-fed mice
3 结 论
本实验初探了牦牛酥油鞘磷脂对高脂饮食诱导的小鼠脂质代谢及肝脏组织炎症的作用。研究发现,牦牛酥油鞘磷脂能够减轻高脂饮食小鼠的体质量和预防高脂饮食引起的高脂血症。通过对肝脏组织中炎症因子及脂质代谢相关基因表达量的分析发现,牦牛酥油鞘磷脂饲喂小鼠后显著下调了组织中促炎症因子的表达,对组织炎症具有很好的预防作用,但鞘磷脂对抗炎症因子的表达量无明显的剂量对应关系,可能是由于饲喂时间较短或者样本较少,存在误差,后续实验还需通过建立炎症模型,对鞘磷脂的抗炎症剂量进行详细的研究;对脂质代谢相关基因的表达分析发现,鞘磷脂能上调FABP1、HMGCR和SCD1基因的表达,下调THRSP、ACC-α等基因的表达,这些与脂肪合成代谢和分解代谢相关基因表达量的变化能够对高脂饮食引起的小鼠血清血脂水平进行调节。综上,本研究可为今后以牦牛酥油鞘磷脂为原料的功能性食品的开发提供基础数据。
猜你喜欢
农产品加工(2022年11期)2022-07-21
青海湖(2022年3期)2022-06-09
滇池(2022年5期)2022-04-30
散文诗(2021年22期)2022-01-12
小哥白尼(野生动物)(2020年9期)2021-01-18
中华养生保健(2020年5期)2020-11-16
西藏文学(2020年4期)2020-08-04
中国生殖健康(2018年1期)2018-11-06
分忧(2015年2期)2015-09-10
