
棉蚜ATP合成酶基因AgoATPb的克隆与表达
2022-03-03 13:18:58杨卫军董艳蕾吴秋芳张美玲韩丽滨张元臣
浙江农业学报 2022年2期
杨卫军,董艳蕾,吴秋芳,张美玲,韩丽滨,张元臣,*
(1.安阳工学院 生物与食品工程学院,河南 安阳 455000;2.河南省太行山林业有害生物野外科学观测研究站,河南 林州 456550)
棉蚜[(Glover)]是一种世界性分布的多食性害虫,能够危害300多种寄主植物,主要以葫芦科、锦葵科、茄科和芸香科植物为主。棉蚜以口器刺入植物韧皮部吸食汁液,常造成植物叶片蜷曲和植物矮缩;且可分泌蜜露导致细菌滋生,严重影响植物光合作用。此外,棉蚜是多种植物病毒的传播载体,能够导致植物病毒病的频繁发生。目前防治棉蚜主要以化学防治为主,然而农药对环境污染严重,对人畜有害,同时抗药性也频繁产生,亟需寻找对环境友好的害虫防治新途径,从基因水平上干扰昆虫的生理系统来防治害虫具有较好的发展前景。
ATP合成酶(ATP synthase)广泛存在于生物细胞的细胞器中,通常由嵌入生物膜的疏水端F0亚基和生物膜外的亲水端F1亚基组成,在动物包括昆虫细胞内催化供能物质ATP的合成。ATP合成酶B亚基在不同物种中保守性非常低,主要起连接和固定F0和F1复合体的装配作用。ATP合成酶B亚基对于维持ATP合成酶的空间构型非常重要,一旦缺失,将导致ATP合成酶构象发生改变,影响ATP的合成。ATP合成酶B亚基在蚊子抗性品系中表达量显著高于非抗性品系,表明其与昆虫抗药性有密切关系。在黑腹果蝇()生殖细胞中抑制ATP合酶B亚基的表达后,雄性果蝇生殖细胞不能发育成熟,从而导致生育能力的降低。缺少ATP合酶B亚基的褐飞虱()和果蝇死亡率明显上升,同时该亚基属于豌豆蚜()和柑橘木虱()唾液腺中的效应蛋白。以上研究表明,ATP合成酶在昆虫体内主要参与氧化磷酸化过程中ATP的合成,对昆虫的生长发育、繁殖,以及适应环境起重要作用。若将ATP合成酶作为棉蚜控制的一个靶标,不仅可以开辟害虫分子调控的新领域,而且能够有效提高对棉蚜的防治效果,为棉蚜防治提供新策略。
目前,已鉴定出多种昆虫的ATP合成酶B亚基基因,尚未见有关棉蚜基因克隆与表达分析的研究报道。本研究以棉蚜为研究对象,克隆得到基因的全长序列,利用实时荧光定量PCR(qRT-PCR)技术分析该基因在棉蚜不同组织和日龄,以及棉蚜取食不同寄主植物后的表达水平,为后续深入研究该基因的功能奠定基础。
1 材料与方法
1.1 棉蚜饲养
2017年4月10日在安阳工学院校内木槿上采集棉蚜,带回实验室将棉蚜转接到棉花叶片上饲养,叶片叶柄用沾水的脱脂棉包被起来,放于直径为15 cm的玻璃培养皿中。培养皿用保鲜膜封口,并用昆虫针刺破保鲜膜,保持空气流通。棉蚜饲养的温度、光周期和相对湿度分别设置为(25±1)℃、14 h/10 h(L/D)和60%±5%。
1.2 植物培养
将棉花、黄瓜、西葫芦和甜瓜种子播种于直径为20 cm、高度为15 cm的花盆中,用2 m×2 m尼龙网网罩(200目)将花盆罩起来,放置于室外进行培养,每间隔5 d浇一次水,植株6个叶片(含子叶)后取叶片用于饲养蚜虫。本实验中所用棉花、黄瓜、西葫芦和甜瓜品种分别为中棉所49、早青二号、欧曼和绿宝石,均购买于市场。
1.3 棉蚜AgoATPb基因部分片段克隆
随机收集在棉花叶片上生长的仔蚜(1~5日龄)和母蚜(产仔后1~5 d)共30~50头,立刻放置于TRIzol(Invitrogen)中,然后用TRIzol法提取总RNA。用琼脂糖凝胶电泳和Nano2000测定RNA的完整性和浓度。以700 ng RNA为模板,按照PrimeScriptRT Reagent Kit with gDNA Eraser(Perfect Real Time)试剂盒(TaKaRa)说明书,合成cDNA模板。
根据转录组序列信息,获得棉蚜基因的部分编码区序列。设计引物ATPb-1F和ATPb-1R(表1),以合成的cDNA为模板进行RT-PCR,对该基因的核苷酸序列进行验证。PCR扩增体系(20 μL):10 μL PCR Mix,上下游引物(10 μmol·L)各0.5 μL,3 μL cDNA模板,ddHO 6 μL。PCR扩增条件:95 ℃ 3 min;95 ℃ 20 s,58 ℃ 20 s,72 ℃ 60 s,35个循环;72 ℃ 8 min;10 ℃保存。PCR扩增产物用1.0%~1.5%的琼脂糖凝胶电泳进行检测,切胶回收目的条带。回收产物用DNA回收试剂盒(OMEGA)回收,将回收产物连接到pEASY-T3(全式金)克隆载体上,导入到感受态细胞DH5α中,进行蓝白斑筛选,鉴定阳性克隆质粒,将阳性质粒送往生工生物工程(上海)股份有限公司进行测序。
1.4 棉蚜AgoATPb基因末端快速克隆
利用1.3节提取的RNA,按照宝生物SMARTer RACE 5′/3′ Kit试剂盒(Takara)说明书合成5′和3′端RACE扩增模板。根据1.3节克隆的基因的部分核苷酸序列分别设计5′和3′端GSP引物和NGSP引物(表1)。按照试剂盒要求的PCR扩增体系和条件进行PCR扩增,之后用1.0%~1.5%的琼脂糖凝胶进行检测,回收目的条带,进行连接、转化、蓝白斑筛选和阳性克隆的测序,方法与1.3节相同。
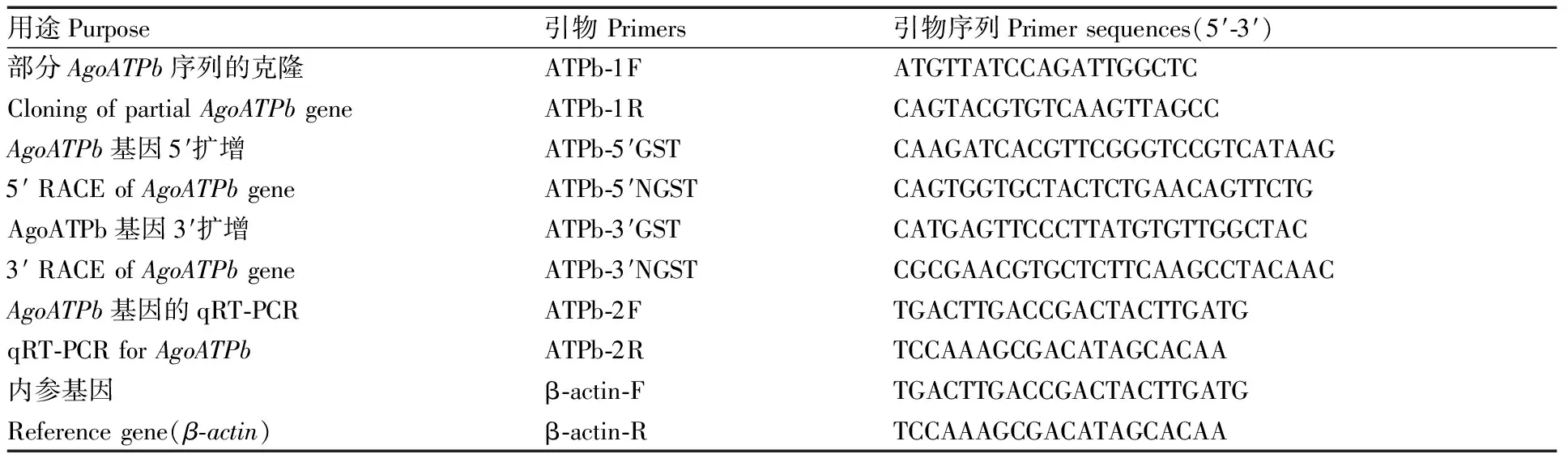
表1 棉蚜AgoATPb基因克隆与qRT-PCR引物
1.5 棉蚜AgoATPb的序列分析与系统发育树构建
用在线工具ORF Finder预测基因的阅读框架;用在线软件ExPASy(https://web.expasy.org/translate/)预测氨基酸的等电点与分子量;利用在线工具http://www.cbs.dtu.dk/services/TMHMM/预测跨膜区;用在线软件Blastp(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行AgoATPb氨基酸序列比对,并下载其他物种同源序列进行系统发育树和多重连配图的构建;利用在线软件SignalP 5.0 Server预测AgoATPb蛋白的信号肽。
1.6 棉蚜AgoATPb基因的时空表达分析
吸取50 μL PBS磷酸缓冲液(pH值8.0)滴在体式显微镜镜头中央,用毛笔挑取4日龄棉蚜置于PBS溶液中,用消毒昆虫针和显微手术镊解剖分离唾液腺、脑、卵巢、中肠和脂肪体等部位。解剖完成后立即放入装有200 μL TRIzol的1.5 mL离心管中,-80 ℃保存备用。收集胚胎期、1日龄、3日龄、5日龄、7日龄、9日龄、13日龄和17日龄的棉蚜,分别提取总RNA,经过琼脂糖电泳和Nano2000确定总RNA质量与浓度后,放置于-80 ℃保存备用。以提取的总RNA为模板,按照PrimeScriptRT Reagent Kit with gDNA Eraser(Perfect Real Time)试剂盒(TaKaRa)说明书反转录获得cDNA,用于qRT-PCR反应,以确定基因的表达水平。每个部位收集50头蚜虫作为一个生物学重复,每个龄期收集30头蚜虫作为一个生物学重复。每个部位和每个日龄3次生物学重复。
1.7 棉蚜AgoATPb基因表达水平
将在棉花上长期饲养的棉蚜,转接到西葫芦叶片上3代后,将1日龄棉蚜分别转接到棉花、黄瓜、西葫芦和甜瓜叶片上,每个叶片转接15头仔蚜,待在4种植物上生长3个世代后,收集10头4日龄的仔蚜,提取总RNA,经过琼脂糖凝胶电泳和Nano2000确定总RNA质量,根据试剂盒要求反转录得到cDNA,进行qRT-PCR反应。每种植物3次生物学重复。
1.8 qRT-PCR
为了避免基因组DNA对qRT-PCR反应的影响,棉蚜700 ng 总RNA用gDNA Eraser去除基因组DNA。之后,根据PrimeScriptRT Reagent Kit with gDNA Eraser(Perfect Real Time)(TaKaRa)试剂盒的说明,以处理过的总RNA为模板合成cDNA。目的基因特异性引物ATPb-2F和ATPb-2R用于确定的表达水平。选取-为内参基因,内参基因的引物为β-actin-F和β-actin-R。引物设计在Primer Premier 5.0中进行,引物序列见表1。qRT-PCR反应在ABI 7500(Carlsbad)上进行,反应体系和反应条件按照说明书进行设置。每个处理设置3个生物学重复和3个技术重复。
1.9 数据分析
基因的表达水平用2法计算。棉蚜不同部位、不同日龄和不同植物寄主棉蚜基因表达差异用单因素方差分析(ANOVA)方法的邓肯氏检验进行比较,差异水平=0.05。所有数据分析都在SPSS软件中完成(Version 14.0)。
2 结果与分析
2.1 棉蚜AgoATPb基因的克隆
以棉蚜cDNA为模板扩增获得了基因的部分核苷酸序列,测序结果表明,该片段长度为697 bp,与预期大小相同。根据获得的部分核苷酸序列设计引物,通过RACE技术,得到了棉蚜基因的全长序列。该基因全长1 247 bp,其中,阅读框822 bp,5′端非编码区128 bp,3′端非编码区297 bp,共编码274个氨基酸残基(图1)。利用生物信息学在线工具(https://web.expasy.org/translate/)对氨基酸序列进行分析,预测棉蚜AgoATPb蛋白分子量为31.40 ku,等电点为8.95。SignalP Server和TMHMMServer预测结果表明,该基因的氨基酸序列无信号肽和跨膜区。

红色代表为起始密码子;*代表终止密码子;黑色横线部分为加尾信号(AATAAA)。
2.2 棉蚜AgoATPb基因的系统发育分析
系统发育树分析结果表明,昆虫的亲缘关系越近,ATP合成酶B亚基氨基酸序列一致性越高,棉蚜、高粱蚜()、豌豆蚜()、大豆蚜()等聚集在一起,与褐飞虱()、茶翅蝽()、二化螟()亲缘关系较远(图2)。多重连配结果表明,AgoATPb与其他昆虫同源氨基酸序列有78个高度保守位点,存在ATP合成酶家族的典型结构域(图3)。将基因的氨基酸序列在NCBI上进行比对发现,其与其他昆虫的一致性为47%~99%。

采用邻接法,自举检验1 000次;图中标尺为遗传距离。

黑色横线部分代表ATP合成酶B亚基的典型结构域。黑色阴影为氨基酸具有100%一致性,灰色表示一致性在50%以上,白色表示一致性在50%以下。AaeATPb,埃及伊蚊;CquATPb,致倦库蚊;GmeATPb,大蜡螟;SfrATPb,草地贪夜蛾;AgoATPb,棉蚜;MpeATPb,桃蚜;ApiATPb,豌豆蚜。
2.3 棉蚜AgoATPb基因时空表达分析
基因在棉蚜唾液腺、脑、卵巢、中肠、脂肪体中都有表达,且在不同部位表达水平存在显著差异(=37.429,<0.001)。在唾液腺和脑中的表达水平虽无显著差异,但是二者表达量均显著高于卵巢和脂肪体,而在中肠表达水平最低;总体上基因在卵巢、中肠和脂肪体中相对表达量较低(图4)。
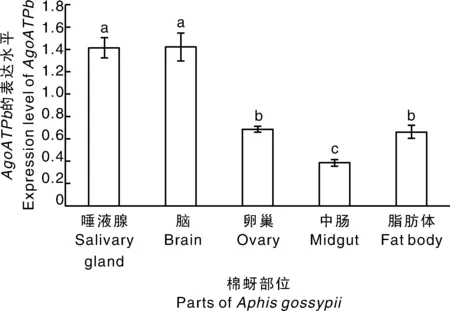
图中的数值用平均值±标准误表示,柱上无相同小写字母表示差异显著(P<0.05)。下同。
基因在不同日龄棉蚜间表达水平存在着显著差异(=23.343,<0.001)。除了在胚胎期不表达外,在其他日龄均有表达,从胚胎期开始表达量上升,3日龄棉蚜中表达水平最高,之后降低,在17日龄表达量最低(图5)。
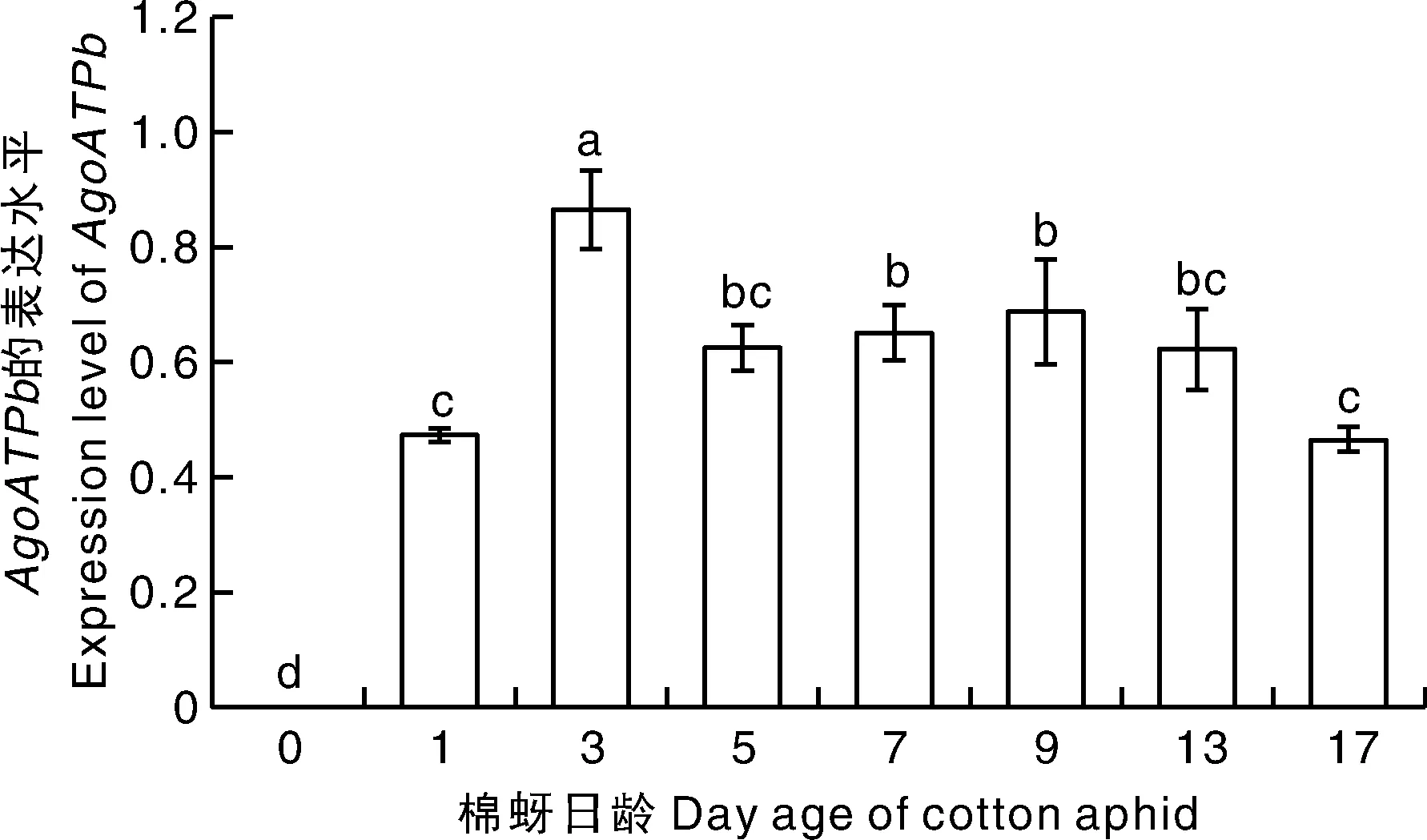
图5 AgoATPb基因在胚胎和不同日龄棉蚜中表达水平
2.4 不同寄主植物上棉蚜AgoATPb基因的表达水平
在棉蚜取食不同植物后,基因表达水平存在显著差异(=64.822,<0.001)。在棉花上棉蚜体内基因表达水平最高,显著高于黄瓜、甜瓜和西葫芦,在黄瓜和西葫芦上表达水平相当,甜瓜上表达水平最低,显著低于在其他植物上(图6)。
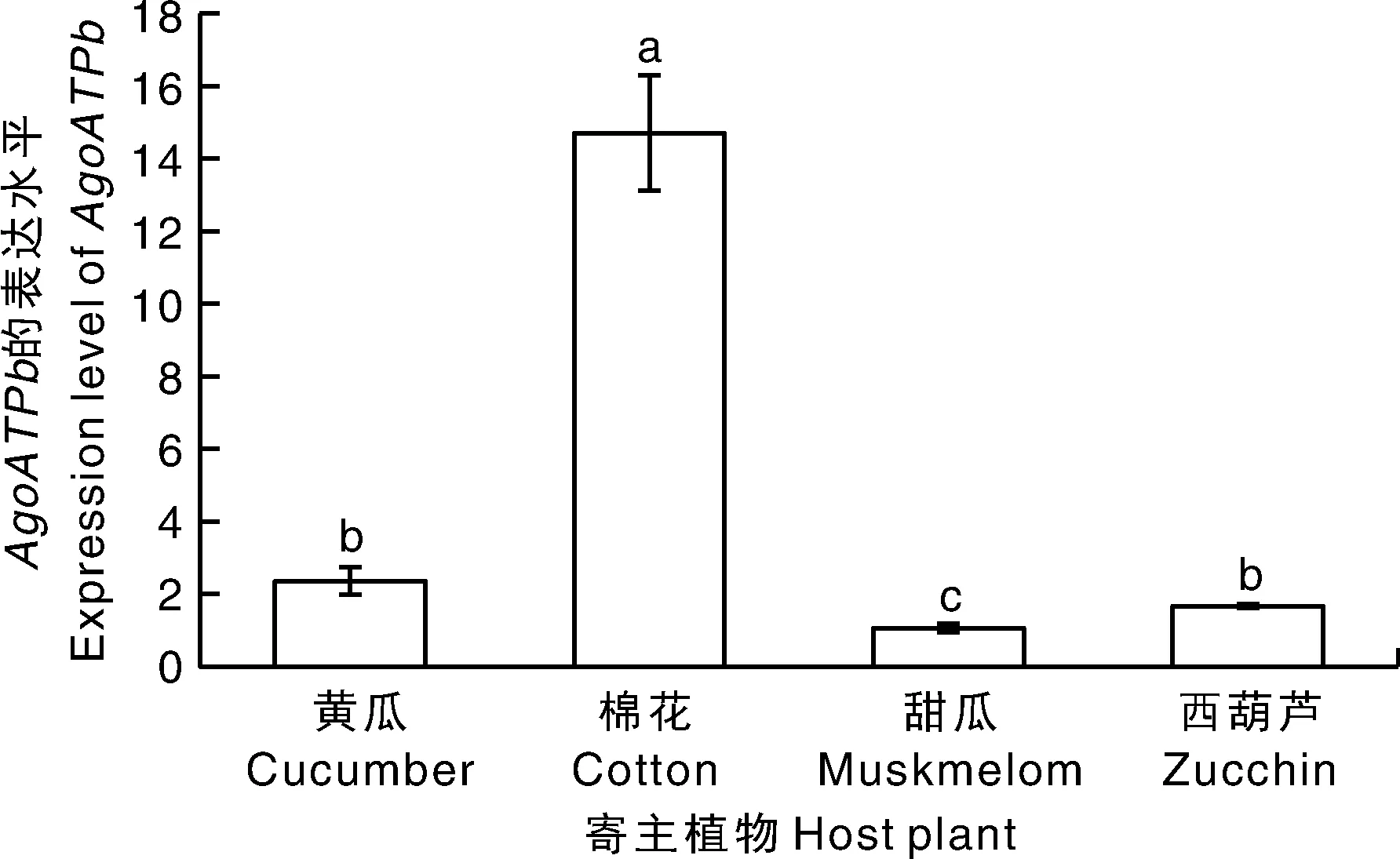
图6 不同寄主植物上4日龄棉蚜体内AgoATPb基因的表达量
3 结论与讨论
ATP合成酶B亚基是昆虫催化供能物质ATP合成的关键基因,但对重要害虫棉蚜ATP合成酶B亚基基因的研究目前还未见有报道。基于此,本研究从棉蚜转录组中筛选出了ATP合成酶B亚基基因的部分核苷酸序列,结合RACE技术获得了棉蚜ATP合成酶B亚基基因的1 247 bp全长序列,其阅读框为822 bp,5′端非编码区128 bp,3′端非编码区297 bp,共编码274个氨基酸残基,其分子量为31.40 ku,等电点为8.95。通过在GenBank数据库中的Blastp进行比对发现,AgoATPb氨基酸序列与其他物种同源氨基酸序列一致性均在47%以上。
根据基因在不同部位表达和龄期的表达,可以推测基因的功能。通常认为ATP合成酶B亚基与能量物质ATP的合成有重要关系,而脂肪体是昆虫能量储存和代谢的主要部位,推测应该在脂肪体具有高水平的表达;然而,本研究发现该基因在脂肪体的表达含量最低,这也说明该基因不仅仅参与能量物质的合成。同时,基因在棉蚜唾液腺中表达水平最高,这可能与棉蚜适应寄主植物有关。因为唾液腺既能够帮助刺吸式昆虫应对植物的防御反应,又能够帮助其顺利取食植物韧皮部的汁液。基因在棉蚜的多个部位均有表达,说明其具有多种生理功能。基因除了在胚胎期不表达外,在棉蚜其他各个日龄均有不同水平的表达,表明其在棉蚜生长发育过程中也起着重要作用。
对豌豆蚜和柑橘木虱唾液腺的研究发现,ATP合成酶B亚基基因存在于昆虫的唾液腺中,推测其与适应寄主植物有关。本研究中,棉蚜取食黄瓜、棉花、西葫芦和南瓜后其体内基因表达水平存在着显著差异。基因表达水平与昆虫适应寄主植物有密切关系,因此昆虫在取食新寄主植物时基因表达水平会发生改变。基因在不同寄主植物上表达水平变化也有可能与棉蚜适应寄主植物有关,基因在棉蚜适应寄主植物中的生理功能有待深入验证。
猜你喜欢
植物保护(2023年1期)2023-02-03 10:22:08
农药学学报(2022年6期)2022-12-27 12:00:28
世界最新医学信息文摘(2020年68期)2020-12-25 11:55:27
新疆农业科学(2020年11期)2020-10-27 09:05:32
现代园艺·综合版(2017年1期)2017-07-14 16:37:59
江苏农业科学(2017年7期)2017-05-23 23:10:25
农业与技术(2016年19期)2016-12-12 02:23:50
现代农业(2016年4期)2016-02-28 18:42:12
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:13
中国医学科学院学报(2013年6期)2013-03-11 20:26:01
