
西北太平洋灯光围网主要渔获物组成及渔场变动研究
2022-03-02 10:10赵国庆石永闯崔雪森唐峰华
南方水产科学 2022年1期
赵国庆,石永闯,樊 伟,崔雪森,唐峰华
1.上海海洋大学 海洋科学学院,上海 201306
2.中国水产科学研究院东海水产研究所/农业农村部远洋与极地渔业创新重点实验室,上海 200090
随着全球近海渔业资源的衰退,加之公海巨大的开发潜力 (占据全部海洋的60%)[1],公海渔业的发展得到世界各国的重视。西北太平洋是我国目前主要的公海作业区域,对我国的远洋渔业发展具有重要的战略意义[2]。西北太平洋隶属于联合国粮农组织 (Food and Agriculture Organization of the United Nations, FAO) 61渔区,该海域受黑潮暖流和亲潮寒流两大水系的共同影响,海洋生物饵料丰富,是鱼类索饵的重要场所[3-4],从而形成了世界上高渔获量的渔场[5]。据FAO统计,近年来该海域年渔获产量稳定在2 ×107t以上[6],渔获物主要有巴特柔鱼 (Ommastrephes bartrami)、秋刀鱼 (Cololabis saira)、远东拟沙丁鱼 (Sardinops sagax)、金枪鱼和日本鲭 (Scomber japonicus)等[7],而灯光围网中上层渔业是2013年左右出现的新兴渔业,渔获物以日本鲭、远东拟沙丁鱼、巴特柔鱼、秋刀鱼为主,其中日本鲭在数量与质量上均占绝对优势[8]。
我国对西北太平洋中上层围网渔业的系统研究起步较晚,在该海域的捕捞技术和生产效率与日、韩等国有一定差距[9]。因此,掌握渔获种类组成、各渔获物种类的季节变动及时空分布规律、渔场重心变动等关键问题,对提高捕捞效率、资源养护和科学管理等尤为重要[10]。虽有诸多研究涉及西北太平洋渔业资源的分布、丰度、渔场变动等内容,但是多集中在柔鱼[11-13]、秋刀鱼[14-16]等单一物种上,尚未见基于长时间序列的实际生产数据对灯光围网的渔获组成、渔场分布变化的相关研究。本研究利用2014—2019年渔业生产资料,采用重心估算与聚类分析方法等,对我国在西北太平洋围网渔业的生产状况、渔获种类组成和渔场重心变动情况进行了分析,以期为西北太平洋渔场的渔情预报、渔业生产与管理提供参考。
1 材料与方法
1.1 数据来源及渔场区域
本文所用数据来源于中国远洋渔业协会公海围拖网渔业技术组所收集的我国西北太平洋公海灯光围网生产统计资料,该资料共由国内14家远洋渔业公司提供 (图1),主要包括作业时间 (年、月、日)、作业位置 (经度和纬度)、渔获种类、投网次数、渔获量等信息。本文所用数据的时间跨度为2014—2019年,主要作业时间为每年的4—11月。数据的空间分辨率为0.5°×0.5°,作业区域分布在145°E—160°E、35°N—44°N (图2)。

图1 2014—2019年渔业统计资料Fig.1 Fishery statistics in 2014–2019

图2 西北太平洋公海灯光围网作业海域Fig.2 Light purse seine fishing area in Northwest Pacific high seas
1.2 数据处理
1.2.1 渔获数据处理
将渔获物中登记的渔获物确定到种;对渔获量数据进行处理,利用单位捕捞努力量渔获量 [Catch per unit effort, CPUE, t·(d·船)−1]来表示资源丰度[17],计算公式为:
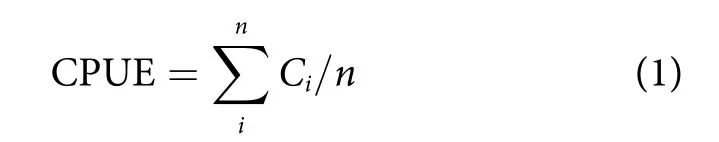
式中:Ci为每艘渔船第i天的渔获量 (t);n为总天数 (d)。
为检验渔船功率与渔获产量是否具有相关性,利用SPSS 19.0软件对渔船功率和产量进行双变量相关分析,以P<0.05作为差异显著水平。在进行相关分析之前,先利用K-S检验验证数据是否符合正态分布。
1.2.2 渔场重心估算
渔场重心的时空变化用渔获量重心法来计算,公式为[18-19]:
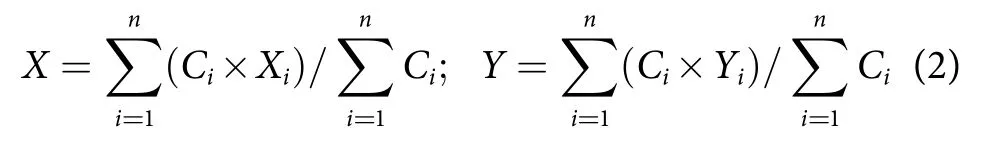
式中:X、Y分别表示某时间段渔场重心的经度和纬度;Ci为第i网次的渔获产量;Xi为第i网次的经度;Yi为第i网次的纬度;n为统计期间的总网次。
1.2.3 渔场重心聚类分析
利用欧式距离来计算年产量重心的变化情况,比较渔场的重心变化,公式为[20]:

式中:Dab为a年和b年产量重心间的距离;Xa、Ya分别为第a年的经度、纬度;Xb、Yb分别为第b年的经度、纬度。根据计算后得到的欧式距离,对2014—2019年各年、各月的产量重心按照最小距离法进行聚类,分析比较年份和月份间的差异性[21]。
数据分析和作图使用Excel 2019、SPSS 19.0、Origin 2018和Matlab 2013等软件。
2 结果
2.1 历年产量与CPUE分析
2014—2019年间,由于投入生产的船只数量有较大的变化 (图1),产量的年间差异较大;统计年间,产量的最大值依次出现在当年的10、8、6、10、9和7月,集中在6—10月,2014、2015、2018和2019年均呈先增后降的变化趋势;统计年间的CPUE波动较为明显,除2016年外,CPUE均呈先增后降趋势,且在10月达到最大值;从月份来看,CPUE 值均超过 10 t·(d·船)−1,介于10.43~30.41 t·(d·船)−1,最大值和最小值分别发生在2014年10月和2018年5月 (图3)。将统计年间的同一月份相结合来分析,产量呈现先增后降的规律,CPUE持续增长到10月,11月与10月持平(图4)。从年份来看,年间CPUE均值除2018年外呈现逐年递减趋势,介于 15.04~23.93 t·(d·船)−1(图5)。对渔获产量进行K-S检验表明,渐进显著性为0.000 (<0.05),渔获产量不符合正态分布,因此使用Spearman和Kendall等级相关系数进行双侧相关性分析。功率与产量的双变量相关性分析结果显示,Spearman和Kendall相关性系数分别为0.000和−0.04,相伴概率分别为0.958和0.428(>0.05),表示功率与产量不存在显著相关关系。

图3 2014—2019年西北太平洋公海灯光围网渔获量和单位捕捞努力量渔获量Fig.3 Catch and catch per unit effort of light purse seine in high seas of Northwest Pacific from 2014 to 2019

图4 2014—2019年西北太平洋公海灯光围网分月份渔获量和单位捕捞努力量渔获量Fig.4 Monthly catch and catch per unit effort of light purse seine in high seas of Northwest Pacific from 2014 to 2019

图5 2014—2019年西北太平洋公海灯光围网单位捕捞努力量渔获量均值Fig.5 Catch per unit effort of light purse seine in high seas of Northwest Pacific from 2014 to 2019
2.2 渔获物组成与季节变化
2014—2019年,渔业统计资料共记录8种主要渔获种类,分别为日本鲭、远东拟沙丁鱼、巴特柔鱼、秋刀鱼、菱鳍乌贼 (Thysanoteuthis rhombus)、细鳞乌鲂 (Brama raji)、日本乌鲂 (B.japonica) 和扁舵鲣 (Auxis thazard),各年度渔获的种类分别为3、6、4、7、5和4种,其中日本鲭、远东拟沙丁鱼和巴特柔鱼在统计年份里均有记录,秋刀鱼除2014年外在其他年份均存在,菱鳍乌贼仅在2017年出现。从月间变化来看,在每个月份均有记载的只有日本鲭;远东拟沙丁鱼、巴特柔鱼和秋刀鱼的出现频率相对较高,依次为87.5%、73%和56.25%;菱鳍乌贼仅出现于2015年的6—10月,细鳞乌鲂、日本乌鲂仅出现于2017年的9和10月,扁舵鲣在2015年的4月、2017年4—6月和2018年10、11月均有出现 (图6)。

图6 2014—2019年各月份渔获种类组成Fig.6 Species composition of catch in each month from 2014 to 2019
渔获产量按月份分析,日本鲭在4月产量占比较低,5—11月产量相差较小;远东拟沙丁鱼和巴特柔鱼在4月产量占比低,5—7月产量占比较高,8—11月产量分布均匀;秋刀鱼在4—6月产量占比低,8—11月产量占比逐月增加,11月达到最大值;菱鳍乌贼、细鳞乌鲂和日本乌鲂出现的月份较少,渔汛旺期发生在8—10月;扁舵鲣4—6月的产量占比达到93.94%,其他月份占比低 (图7)。

图7 每种渔获各月份占比Fig.7 Proportion of each catch in each month
渔获产量年际分析发现,日本鲭的比重占有绝对优势,2014—2016年的产量占比均在95%以上,但是统计年份内日本鲭的占比呈现持续降低的趋势,2019年达到最低值 (58.87%),CPUE的变动与产量占比变化趋势一致,介于9.2~22.16 t·(d·船)−1(图8-a);远东拟沙丁鱼的比重逐年升高,在2019年达到最大值40.52%,CPUE波动增长,介于4.84~7.08 t·(d·船)−1(图8-b);巴特柔鱼的比重在 2016 年达最大值 (1.99%),2018年达最低值(0.0087%),CPUE 介于 0.443~3.13 t·(d·船)−1(图8-c);秋刀鱼仅出现于2015—2019年,比重为0.06%~0.6%,CPUE 介于 1.38~4.17 t·(d·船)−1(图8-d);菱鳍乌贼、细鳞乌鲂、日本乌鲂和扁舵鲣仅在个别年份出现,比重极低且总和不超过0.1% (图8-e—8-h)。统计结果显示,西北太平洋灯光围网的捕捞对象主要以日本鲭为主,近年来远东拟沙丁鱼的产量占比逐年增加,2019年的比重达到40.52%。

图8 各渔获种类的年间单位捕捞努力量渔获量和占总渔获量的比重Fig.8 CPUE of catch species and its proportion in total catch
2.3 渔场重心变化分析
2.3.1 渔场重心季节变化
渔场重心呈现逆时针变化规律,从4月开始逐渐向东北方向移动,8、9或10月到达最东北端,然后向西南方向折返,且统计年间渔场重心的变动趋势一致,可见渔场重心有明显的季节变化(图9—10)。渔场重心经、纬度的变化介于145°E—155°E、35°N—43°N;统计年间,在经、纬度上变动范围最大的均为2014年,变动范围最小的均为2016年 (图9)。从年间变化来看,西北太平洋灯光围网的渔场重心呈现逐渐向东北移动的趋势,从2014年至2019年重心在经度上东移0.92°,在纬度上北移 1.57°。

图9 2014—2019年西北太平洋渔场重心季节变化Fig.9 Seasonal variation in fishing ground gravity center in Northwest Pacific from 2014 to 2019

图10 2014—2019年西北太平洋渔场重心年度变化Fig.10 Interannual variation in fishing ground gravity center in Northwest Pacific from 2014 to 2019
2.3.2 渔场重心聚类分析
2014—2019年,将相同月份的产量重心进行综合统计分析可得,不同月份间产量重心分布存在较大差异。由欧式距离可得,月间产量重心的空间距离介于0.363 (8月/9月)~5.917 (4月/9月)。4月与其他月份间的欧氏距离均大于1,两个相邻月份间的欧式距离分布介于0.363 (8月/9月)、2.885(10月/11月,表1)。经聚类分析可得,若以空间距离2为阈值,则可将月产量重心分为4类,分别为7、8、9和10月,4月,5、11月,6月 (图11)。

表1 西北太平洋灯光围网各月产量重心的空间距离Table 1 Spatial distance of center of gravity of monthly yiled of light purse seine in Northwest Pacific

图11 月产量重心聚类结果Fig.11 Clustering results of monthly yield barycenter
以年间产量重心计算可得,不同年份间的产量重心分布相对较为集中。年间欧式距离的产量重心的空间距离为0.197 (2016年/2017年)~1.800(2014年/2019年)。2014年产量重心与其他年份产量重心的距离除2015年外均大于1;除2015和2019年外,其他年份两两间的产量中心距离均小于1。两相邻年份间的欧式距离介于0.197 (2016年/2017年)~0.794 (2015年/2016年,表2)。经聚类分析可得,若以空间距离1为阈值,则可将年产量重心分为4类,分别为2016、2017、2018年,2019年,2014 年,2015 年 (图12)。

图12 年产量重心聚类结果Fig.12 Clustering results of annual yield barycenter
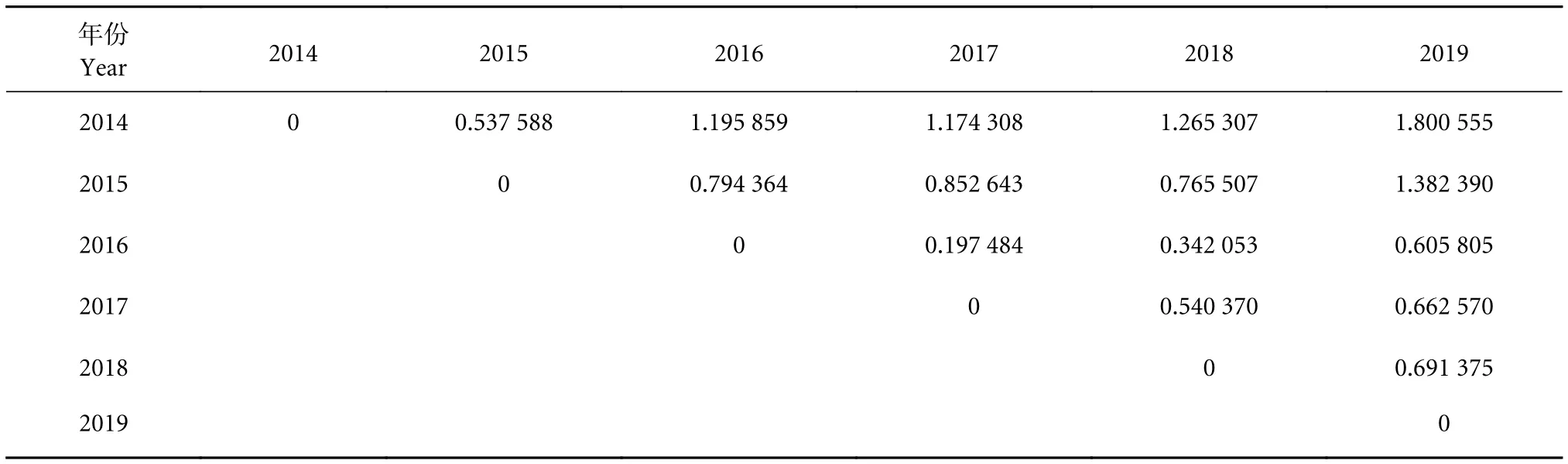
表2 西北太平洋灯光围网各年产量重心的空间距离Table 2 Spatial distance of center of gravity of annual yiled of light purse seine in Northwest Pacific
3 讨论
3.1 CPUE分析
CPUE能否作为渔业资源丰度指数来表征资源密度的变化一直是渔业资源相关研究中争议的焦点[22-23],但不可否认的是,通常假定CPUE即为资源丰度指数,与资源量、资源密度有呈正比的线性关系,并被广泛应用于渔业资源研究和渔业管理中[24-26]。不同情况下CPUE的计算方法有所区别,一般分为3类[27]:1) 直接用渔获量之和除以总捕捞努力量;2) 首先划定好空间分辨率 (本文为0.5°×0.5°),用小区域内的总渔获量除以区域内的总捕捞努力量;3) 先求出每个点要素对应的CPUE值,然后划分好空间分辨率,对区域内的所有CPUE求均值。目前在西北太平洋灯光围网相关研究中,时常将时间、船的数量等作为捕捞努力量来计算CPUE[8,28-31],并不以拖网渔船中常用到的渔船功率作为捕捞努力量,这是由灯光围网的作业方式所决定的,对其产量影响较大的是围网所围住的海域面积、灯光的诱集作用、鱼群的密度等,CPUE受年份、月份、纬度、船队、年份与纬度和年份与船队间的交互效应影响[32],因此本文将作业时间和渔船数量作为捕捞努力量计算CPUE。
日本鲭是西北太平洋灯光围网的主要捕捞对象[8,29],西北太平洋灯光围网的捕捞情况通常受日本鲭支配。据报道,西北太平洋渔业资源的产量和CPUE均具有明显的季节变化[28-30],本研究显示渔获产量和CPUE 每年的变化规律基本一致,4—11月基本上呈现先增后降的趋势,本文得出的产量和CPUE月间变化规律与戴澍蔚等[29]对2014—2015年日本鲭的研究结果类似。渔船自4月出航并进行渔场的找寻,因此开始捕捞月份的渔获产量通常较低[29]。庄之栋等[33]研究指出,8—10月西北太平洋日本鲭处于索饵洄游期,相对集中;9—10月处于生物学较大值,产量和CPUE均较大;11月较大的风浪对捕捞产生了一定影响,因此渔获产量和CPUE均受影响[29]。本研究还显示,CPUE在统计年间出现了逐年递减的趋势,而对此并没有可靠的解释,管理措施的实施、渔船数量的变化、一天内捕捞作业的时长等均可能对其产生影响,后续需对此作更深入的研究。
3.2 渔获量及渔获物组成分析
远洋渔业与近海渔业最大的区别在于海域与作业方式,远洋渔业的资源种类较为单一。本研究统计到8种渔获物,以日本鲭、远东拟沙丁鱼、巴特柔鱼、秋刀鱼为主,而日本鲭在产量比重和时空分布上均占绝对优势,这与徐博等[28]的研究结果一致。董秀强等[8]在西北太平洋的灯光围网调查发现,主要渔获物为日本鲭、柔鱼和远东拟沙丁鱼,其中日本鲭占绝对优势,产量占比为81.77%,丰富度指数为0.84。本研究发现,远东拟沙丁鱼的产量比重逐年增加,2019年增至40.52%,正逐渐成为西北太平洋灯光围网的主要捕捞鱼种之一,下一步可加强对远东拟沙丁鱼的相关研究。渔获种类数量在年间和月间均存在一定差异,各渔获种类的产量和CPUE在年间和月间的差异较大,菱鳍乌贼、细鳞乌鲂、日本乌鲂和扁舵鲣仅在个别年份的个别月出现,原因可能是捕捞随日本鲭而发生了移动,而捕捞位置的海洋环境并不是这些鱼种最适的生存条件,也有可能是灯光围网作业方式达到的水层较浅或相应的光照强度、光色等不适合捕捞这些鱼种。相关研究指出,不同种类的鱼类趋光性不同,具有正向趋光性的多为中上层鱼类,如鲐鲹类、沙丁鱼、秋刀鱼、竹䇲鱼等[34],而具有趋光性的不同鱼种的感光范围有差异,如日本鲭的感光介于0.01~14 lx[35],秘鲁外海茎柔鱼 (Dosidicus gigas) 的适宜照度为0.2~1.0 lx[36]。因此,集鱼灯的类型可能对渔获种类造成一定影响。
3.3 渔场变动及其影响因素
西北太平洋渔场介于 145°E—155°E、35°N—43°N,渔场重心具有明显的季节变化。在统计年份中,每年渔场重心的变化趋势一致,从4月开始逐渐向东北方向移动,8—10月到达最北端,然后向西南方向折返。因西北太平洋灯光围网主要捕捞对象为日本鲭,故渔场重心的变化与日本鲭渔场重心的变化以及日本鲭的洄游路线存在一定的相似性[28-30]。有研究指出,捕捞渔船的作业路线是渔获量重要的影响因素[37]。Sukyung等[38]通过对1970年以来西北太平洋日本鲭的捕捞量、补充量和生物量等捕捞数据,以及温度、盐度和经向风强度等环境数据的研究发现,随着海温的升高,日本鲭的栖息地出现逐渐向北扩展的现象,本文也发现了西北太平洋渔场逐年向东北方向移动的现象,但具体结论还需要更长时间序列和更完善的数据支撑。
渔场变动与环境因素息息相关[38]。水温是影响渔业资源生存状态的重要因素[39]。王良明等[31]通过对西北太平洋进行定点调查得出,春季渔场海表温度 (SST) 介于7~19 ℃,最适SST为11~15 ℃;夏季渔场SST介于8~24 ℃,最适SST为8~12 ℃。Cheunge等[40]指出,日本鲭的栖息水温一般为10~27 ℃,最适栖息水温约为25 ℃。徐博等[28]研究显示日本鲭的最适SST为13~20.18 ℃。Sukyung等[38]研究发现随着海温的升高,日本鲭的栖息地随之发生变化。全球气候变暖导致SST过高,会对日本鲭栖息地造成很大影响[41]。此外,西北太平洋秋刀鱼、柔鱼、金枪鱼等的作业渔场水温与日本鲭有所重合。晏磊等[42]研究认为,秋刀鱼渔场分布于SST 10~17 ℃的海区,最佳捕捞SST为10~13 ℃;唐峰华等[43]研究指出,北太平洋柔鱼最适SST为14~19 ℃;张衡等[44]指出,金枪鱼的渔获量主要分布介于16~23 ℃内。
海流的分布和变化对于渔场的形成和消失有重要影响[45]。本研究发现2014—2019年渔场的重心往东北方向偏移,在经度上东移0.92°,在纬度上北移1.57°。黑潮暖流和亲潮寒流共同影响着西北太平洋的生物地理环境,是渔场空间格局形成和变动的关键因素,尤其是黑潮蛇行的发生对渔场中下层的海水温度有很大影响,而且在黑潮逆流区表层以下观测到小范围不规则水流的存在,这种水流造成了中层水域温度及其他因子的波动,从而对西北太平洋渔业、生态系统以及气候环境等产生重要影响[46-47]。相关研究表明,黑潮暖流和亲潮寒流的强弱变化对于同一海区的SST、温度变化程度影响显著,从而影响渔期爆发的时间及其长短[43]。不同水系、不同水团所形成的锋区和温跃层与具有洄游习性种类的洄游和垂直活动密切相关[48],例如在日本海和太平洋沿岸,太平洋褶柔鱼 (Todarodes pacificus) 幼体的活动路线随黑潮主轴的移动路径而变化[49]。日本鲭渔场主要是流隔型渔场,洄游路线与海流密切相关,同时受海洋环境影响较大[50]。太平洋年际震荡现象、南方涛动和北极涛动等气候事件也对渔业资源的分布有着重要影响[51]。海流可以改变叶绿素a的浓度,而叶绿素a是影响渔业资源分布的重要因素,是估算海洋生产力的基本指标,其含量通常用于表征浮游植物生物量,从食物链的角度来看,与渔场分布存在间接关系[52],因此常被用于预测渔场位置[53-54]。唐峰华等[43]指出,可以利用叶绿素a浓度和分布情况辅助水温来分析暖寒流交汇区域的变化情况。长期的寒暖流交汇作用,导致表层及垂直水温的变化,很有可能是驱动渔场整体向东北移动的一个关键因素,但需要结合更深入和大尺度的遥感GIS追踪研究来剖析。渔场的形成也是气象要素、海洋水文特征、饵料、气候变化等其他多种因素互相作用的结果。
猜你喜欢
中国检察官(2022年20期)2022-11-07
中国水产(2021年12期)2021-12-06
中国水产(2021年6期)2021-07-01
大自然探索(2021年12期)2021-02-07
文萃报·周五版(2020年40期)2020-11-06
环球人物(2020年19期)2020-10-10
农家之友(2020年9期)2020-04-21
当代水产(2019年4期)2019-05-16
浙江海洋大学学报(自然科学版)(2019年5期)2019-03-21
金山(2018年8期)2018-11-09
