
和厚朴酚对猪热应激卵母细胞体外成熟的影响
2022-03-02 06:23金小虎石学颖孙良振林敬轶郭海涛王婧然王漠然岳顺利周佳勃东北农业大学生命科学学院黑龙江省动物细胞与遗传工程重点实验室黑龙江哈尔滨150030
中国兽医学报 2022年1期
金小虎,石学颖,孙良振,林敬轶,刘 雪,郭海涛,王婧然,王漠然,岳顺利,周佳勃 (东北农业大学生命科学学院 黑龙江省动物细胞与遗传工程重点实验室,黑龙江 哈尔滨 150030)
动物胚胎工程所需卵母细胞多来自于屠宰场废弃卵巢中的卵丘-卵母细胞复合体(COCs)。COCs再经过体外成熟(in vitromaturation,IVM)可得到大量廉价可用卵母细胞。优质的卵母细胞是体外受精、克隆动物、转基因动物生产成功与否的重要保证[1]。因此,改进体外成熟技术,提高卵母细胞体外成熟质量越来越受到人们的重视。动物运输、气候异常导致高温往往使动物遭受热应激。有报道显示,猪卵母细胞在生发泡破裂(GVBD)至4细胞(4-cell)之间的发育阶段对热应激具有高度敏感性[2]。热应激还会诱发氧化应激[3],造成线粒体功能紊乱[4],从而产生过量的活性氧(ROS),而过量的活性氧会进一步导致卵母细胞体外成熟利率以及胚胎发育率的下降。
Sirtuins蛋白家族成员Sirt3定位于线粒体基质中,可以直接通过去乙酰化作用激活超氧化物歧化酶2(SOD2)调节细胞内的氧化应激状态[5],在卵母细胞和植入前胚胎中发挥保护作用[6]。和厚朴酚(3′,5-二-(2-丙烯基)-1,1′-联苯-2,2′-二醇)是一种天然木脂素,目前已被广泛用于治疗各种疾病[7]。和厚朴酚可以提高过氧化物酶体增殖物激活受体γ辅激活因子1α(PGC-1α)水平和活性,提高线粒体的活性,保护线粒体在氧化应激下的功能[8]。而PGC-1α可以调节Sirt3活性,Sirt3介导PGC-1α对细胞内活性氧生成和线粒体生物发生的影响[9]。
因此,本试验以卵巢体外热应激模型为基础,研究和厚朴酚对猪热应激后卵母细胞体外成熟和后续发育中的作用,为Sirt3及其控制的抗氧化防御体系在相关病理及生理机制研究提供理论参考。
1 材料与方法
1.1主要试剂所使用的生化试剂与药品如无特殊说明均购自Sigma-Aldrich公司。
1.2主要液体的配制体外成熟液是TCM199培养液基础中添加3.05 mmol/L葡萄糖、0.91 mmol/L丙酮酸钠、1 mmol/LL-谷氨酰胺、0.57 mmol/LL-半胱氨酸、0.01 IU/L PMSG、0.01 IU/L hCG、75 mg/L青霉素、50 mg/L硫酸链霉素、10%猪卵泡液。根据参考文献[8]的方法,先将和厚朴酚用DMSO配制成10 mmol/L的浓储液,—20℃保存,在试验时根据需要用体外成熟液稀释成10μmol/L的工作液。
1.3卵巢的获取与热应激处理将获取自屠宰场的新鲜猪卵巢放入37℃含青霉素100 mg/L、链霉素50 mg/L生理盐水的保温桶中,1 h之内带回实验室,使用经过预热并添加青霉素与链霉素的生理盐水冲洗。根据参考文献[10]的方法将部分卵巢放入温度为41.5℃的培养箱中进行1 h的热应激处理。
1.4卵母细胞的收集与培养用20 m L注射器抽取直径3~6 mm卵泡中COCs。抽取结束后卵泡液转入离心管中,37℃温箱中静置10 min后弃上清,重复2次后收集沉降的沉淀COCs,在TLHepes-PVA液中挑选胞质均匀的COCs,随后按照30枚COCs一组加入到预先过夜平衡的成熟液微滴(150μL)中,在38.5℃、5%CO2、饱和湿度的培养箱中培养44 h。
1.5活性氧水平检测卵母细胞中的活性氧水平检测是根据参考文献[11]的方法,使用活性氧检测试剂盒(Beyotime,Cat#S0033)检测。取适量的检测液用培养液或DPBS按照1∶1 000的比例稀释使其终浓度达到10μmol/L,将去卵丘卵母细胞加入此工作液中孵育30 min,孵育完毕后用DPBS充分洗净并进行图像采集。
1.6线粒体膜电位检测取适量JC-1(Beyotime,Cat#C2006)使用培养液按照1∶200的比例稀释后制成培养微滴覆盖石蜡油,放入二氧化碳培养箱中进行平衡。平衡结束后将去卵丘卵母细胞加入到含有JC-1的培养微滴中,在培养箱中孵育20 min。孵育结束后立即使用荧光显微镜观察并统计红色和绿色荧光的卵母细胞的比例。
1.7Sirt3、SOD2免疫荧光染色根据参考文献[12]的方法将卵放入4%多聚甲醛中固定30 min,固定完毕后用杜氏磷酸盐缓冲液(DPBS)清洗3次,使用1%Triton-X100对卵母细胞进行通透处理30 min,处理结束后继续用DPBS清洗3遍,使用3%BSA进行封闭处理,封闭时间为1 h,随后将卵母细胞在一抗(Sirt3抗体,货号bs6105R或者SOD2抗体,货号AF4360)中4℃孵育过夜,使用DPBS清洗3次,二抗孵育2 h,DPBS清洗3次后将使用Hoechst33342对染色体进行染色10 min,用DPBS将卵母细胞充分洗净后将其小心加入到已滴加至载玻片上的防荧光淬灭剂液滴中,封片后使用激光共聚焦显微镜进行观察与拍照。
1.8猪卵母细胞孤雌激活将成熟卵母细胞加入到含有5μmol/L离子霉素的PBS操作液中激活5 min,随后将卵母细胞放入用含2 mol/L 6-二甲氨基嘌呤的PZM-3培养液培养5 h。激活完毕后,将胚胎转入经过预先平衡的PZM-3培养液中培养,并统计发育情况。
1.9数据统计与分析试验数据用GraphPad Prism7软件进行分析,显著性统计使用软件中的ANOVA模块进行单因素方差分析。数据用平均数±标准误(x±s x) 表示。所有试验除特殊说明外至少重复3次。
2 结果
2.1和厚朴酚对热应激卵母细胞成熟率的影响在经过44 h的体外成熟后,热应激组卵母细胞体外成熟率要明显低于对照组(P<0.01);在热应激+和厚朴酚组的卵母细胞成熟率显著高于热应激组卵母细胞成熟率(P<0.05)(图1)。结果表明,和厚朴酚可以提高热应激卵母细胞的成熟率。

图1 和厚朴酚对热应激卵母细胞成熟率的影响
2.2和厚朴酚对热应激卵母细胞中Sirt3蛋白的影响通过免疫荧光染色证实在正常卵母细胞中Sirt3分布于GV期卵母细胞的胞质中,但在生发泡周围有较为明显的聚集;在卵母细胞成熟过程中Sirt3水平在细胞质中增加;而经历热应激后,Sirt3荧光强度出现了减弱,并贯穿体外成熟的整个过程;热应激+厚朴酚组的成熟卵母细胞中Sirt3的荧光强度增强并接近于对照组(图2)。
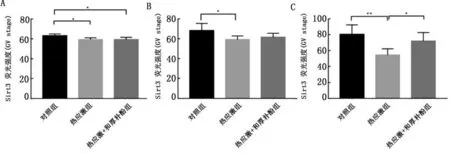
图2 和厚朴酚对热应激卵母细胞中Sirt3的影响
2.3和厚朴酚对热应激卵母细胞中SOD2的乙酰化程度的影响使用SOD2乙酰化抗体检测和厚朴酚对热应激卵母细胞SOD2的乙酰化程度的影响。结果显示,经过热应激后GV期的猪卵母细胞中SOD2的乙酰化程度要高于对照组(P<0.05),热应激组卵母细胞在MⅠ期、MⅡ期的乙酰化程度分别高于对照组(P<0.05,P<0.01);在热应激+和厚朴酚组,卵母细胞SOD2的乙酰化程度呈现降低趋势,与热应激组相比差异显著(P<0.05)(图3)。
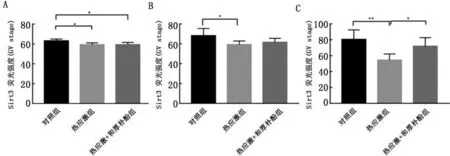
图3 和厚朴酚对热应激卵母细胞中SOD2的影响
2.4和厚朴酚对热应激卵母细胞中的ROS水平的影响对卵母细胞中的ROS水平进行检测后,结果发现MⅡ期,热应激组卵母细胞的平均荧光强度要显著强于对照组(P<0.01),而和厚朴酚的加入使得热应激卵母细胞的ROS水平显著降低(P<0.05)(图4)。
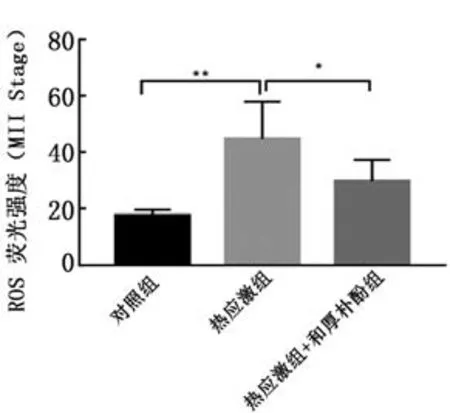
图4 和厚朴酚对热应激卵母细胞(MⅡ期)中ROS水平的影响
2.5和厚朴酚对热应激卵母细胞线粒体膜电位的影响使用JC-1荧光探针对猪卵母细胞的线粒体膜电位进行测定。通过红∶绿荧光JC-1染色法测定的卵母细胞线粒体活性发现,热应激卵母细胞红∶绿比值显著低于对照组(P<0.01);添加和厚朴酚后,卵母细胞线粒体膜电位相较于热应激组显著升高(P<0.05),表明和厚朴酚对于提高热应激卵母细胞线粒体膜电位保护线粒体功能具有积极的作用(图5)。
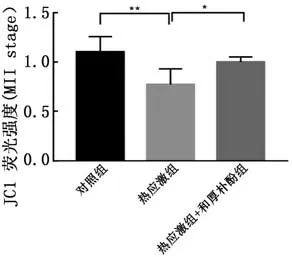
图5 和厚朴酚对热应激卵母细胞(MⅡ期)线粒体膜电位的影响
2.6和厚朴酚对热应激卵母细胞的体外发育能力的影响热应激组卵母细胞经孤雌激活后胚胎发育能力相比对照组降低极显著(P<0.01),热应激+和厚朴酚组胚胎的体外发育能力与热应激组相比显著提高(P<0.05)(图6)。结果表明,和厚朴酚对于热应激卵母细胞的体外发育能力有提高作用。
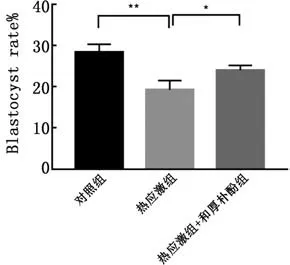
图6 和厚朴酚对热应激卵母细胞孤雌激活胚胎体外发育能力的影响
3 讨论
夏季高温、动物运输往往使动物遭受热应激,会导致动物生殖能力下降。热应激后机体的氧化应激也会提高,从而造成生殖细胞的氧化损伤,最终降低细胞的发育能力[11]。
健康细胞的线粒体会严格地控制细胞呼吸过程中ROS的生成。如果细胞ROS升高会影响到线粒体正常的生理功能[13]。在糖尿病模型小鼠卵母细胞中线粒体的功能性障碍就是由ROS水平升高所导致[14]。有研究发现,在猪的成纤维细胞中ROS含量过高可引起线粒体膜电位的下降[15]。本试验结果表明,热应激后的猪卵母细胞的ROS含量显著升高、线粒体膜电位显著下降。而在成熟液中添加和厚朴酚,热应激卵母细胞的线粒体膜电位与热应激组相比显著升高。这种现象与在中国仓鼠卵巢细胞[16]以及大鼠心肌细胞中[17]的研究中添加和厚朴酚保护线粒体膜功能的结果是相一致。
锰超氧化物歧化酶2(SOD2)属于金属过氧化物歧化酶家族成员。SOD2广泛存在于线粒体。其表达以及活性的降低将导致对线粒体内产生的ROS的清除作用降低,从而导致线粒体DNA的过氧化损伤[18]。SOD2蛋白序列第122位的赖氨酸残基以及该残基周围一段由13个氨基酸组成的特殊的序列,在哺乳动物进化中具有保守性[5],通过检测该残基的乙酰化程度可以判断SOD2的活性高低。Sirt3定位于哺乳动物线粒体基质中,具有NAD+依赖的去乙酰化酶活性[19]。据报道,Sirt3能够特异性的调节SOD2的活性,是调节线粒体ROS稳态的关键分子[14]。
卵母细胞健康发育依赖于氧化剂和抗氧化剂所维持的稳定状态[20]。LIU等[14]发现,过表达Sirt3可以显著降低糖尿病模型小鼠的卵母细胞氧化应激状态。本试验结果表明,热应激后卵母细胞的Sirt3蛋白的表达量下降和SOD2乙酰化水平上升,而添加了和厚朴酚却可以提高Sirt3蛋白的表达量和降低SOD2的乙酰化水平。据报道,和厚朴酚是Sirt3的激活剂具有很强的抗氧化性能[8],它可以逆转小鼠的心肌肥厚[21]。另一项研究结果表明,和厚朴酚可以有效激活t-BHP损伤的肝细胞中的Sirt3,同时降低SOD2的乙酰化程度,进而提高SOD2的活性,使肝细胞免受氧化损伤[8]。在试验结果表明,热应激猪卵母细胞的体外成熟率和发育率显著下降,而添加和厚朴酚可以减轻热应激对卵母细胞造成的损害,表明和厚朴酚可能是通过激活卵母细胞中的Sirt3蛋白,促进SOD2发生去乙酰化,提高抗氧化酶活性,清除过量的ROS,增强了卵母细胞线粒体功能,进而提高热应激卵母细胞的成熟率和囊胚率。
综上所述,向成熟液中添加和厚朴酚可以改善卵细胞体外成熟质量,维持体外成熟卵母细胞的成熟效率并提高体外发育率,这些结果可以为优化细胞体外培养系统提供理论参考。
猜你喜欢
中国畜牧杂志(2022年9期)2022-09-14
中国畜牧杂志(2022年4期)2022-04-15
科学导报(2022年11期)2022-03-03
中国药学药品知识仓库(2021年11期)2021-11-25
中小学德育(2020年11期)2020-03-18
教育界·上旬(2016年12期)2017-05-25
江苏农业科学(2016年8期)2017-02-15
风湿病与关节炎(2016年12期)2017-01-14
环球人文地理(2016年10期)2016-11-24
江苏农业科学(2016年7期)2016-10-20
