
2016—2020年华北地区猪繁殖与呼吸综合征病毒分子流行病学调查
2022-03-02 06:23王丙雷陈少杰顾文源范京惠左玉柱河北农业大学动物医学院河北保定0700河北省兽医生物技术创新中心河北保定0700邢台市邢东新区动物卫生监督所河北邢台05400
中国兽医学报 2022年1期
王丙雷,王 晶,刘 媛,陈少杰,顾文源,郭 禹,2,范京惠,2*,左玉柱,2* (.河北农业大学动物医学院,河北 保定 0700;2.河北省兽医生物技术创新中心,河北 保定 0700;.邢台市邢东新区动物卫生监督所,河北 邢台 05400)
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)也被称为猪神秘病或蓝耳病,该病是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)引起,以各年龄段猪表现呼吸道症状,妊娠母猪流产、死胎以及高病死率为主要特征[1-2]。PRRS最早于1987年发生于美国,并很快流行于欧洲和美洲[3]。直至2006年由变异毒株HP-PRRSV为病原引起的高致病性PRRS在我国南方暴发,引起了人们的关注和重视,随后在国内出现了极易的类NADC30毒株[4]。高致病性毒株在全国范围内传播,给养猪业造成了严重的经济损失,已成为养猪业中危害最为严重的疾病之一[5]。
PRRSV为动脉炎病毒属的成员,单股、正链、不分节段RNA病毒,该病毒基因组长度约为15 kb,编码9个开放阅读框(ORF):1a,1b,2a,2b,3~7[2]。由于依赖RNA的RNA聚合酶缺乏3′至5′核酸外切酶的校对能力,因此RNA病毒的突变率很高。PRRSV核苷酸取代的计算速率为每年(4.7~9.8)×10—2/位点,是报道的RNA病毒中最高的[6]。PRRSV的糖蛋白5(GP5)由开放阅读框5(ORF5)编码,包含多个中和表位,其编码序列通常用于PRRSV分离株的系统发育分析和分类。通过严格的分子时钟模型对分离毒株进行BEAST估计,得出ORF5的平均替代率为7.966×10—3,可信区间的95%为每年(6.943~9.031)×10—3替代/位点[6]。迄今为止,PRRSV已在全球范围内被识别,并分为2种不同类型:PRRSV-1(欧洲血统,原型病毒Lelystad病毒,LV)和PRRSV-2(北美血统,原型病毒,VR-2332)[7],这2个基因型共有50%~80%的氨基酸序列和55%~70%的核苷酸同一性[8]。根据ORF5序列间的遗传关系,Ⅱ型PRRSV分为9个谱系,谱系1~谱系9(L1~L9)[9]。同样,每种病毒亚型的菌株之间也存在广泛的遗传变异。这种变异意义重大,因为实验室和现场研究支持以下论点:遗传上不同的PRRSV变异体可能具有固有的毒力和临床特性。由ORF5基因编码的GP5蛋白作为主要的结构蛋白,其功能最复杂和最易变异,该蛋白在病毒附着和内在化过程中发挥了重要作用[10-11]。因此,检测ORF5基因的遗传变异对PRRSV的流行病学研究具有重要意义。目前,PRRSV已被视为控制猪场疾病的主要问题之一,虽然商品化的疫苗可预防和控制PRRSV,但无法为猪群提供充足且有效的保护[12],且导致疫苗免疫失败的重要因素是PRRSV的遗传多样性[11]。因此,对流行株的遗传多样性分析将有助于制定有效的预防和控制策略,本试验为进一步了解在2016—2020年华北地区PRRSV的流行病学和遗传特征,对测序得到的33条PRRSV ORF5基因开展遗传进化及分子流行病学分析。
1 材料与方法
1.1样品采集2016—2020年,在华北地区的不同猪场中采集到692份临床组织样本,—80℃保存。
1.2引物设计参考GenBank上登录的PRRSV ORF5基因设计1对引物ORF5-F/R,引物由上海生工生物工程有限公司合成。ORF5-F:5′-AGGCTTGACCCTGCCTGCCTTGA-3′,ORF5-R:5′-GAATTCACAAGCAGTGCCGACTG-3′。
1.3病毒核酸的提取根据RNeasy Mini试剂盒说明书提取病毒总RNA,并通过反转录试剂盒PrimeScriptTM1st Strand c DNA Synthesis Kit进行反转录,以获得cDNA,—80℃保存。
1.4PRRSV ORF5基因扩增以1.3中获得的cDNA为模板,ORF5-F/R为引物对PRRSV阳性样品进行PCR扩增,反应体系为20μL:10 mmol/L d NTP 3μL,10×PCR Buffer 2μL,Taq酶0.5μL,2μL cDNA,上、下游引物各1μL,10μL dd H2O。PCR反应程序:94℃5 min;94℃30 s,55℃30 s,72℃1 min,35个循环;72℃10 min。PCR产物于1.5%琼脂糖凝胶中电泳,经鉴定为阳性的PCR产物送至上海生工生物工程有限公司测序。
1.5PRRSV ORF5基因遗传进化分析在MEGA 7.0软件中利用1 000个重复的最大复合似然和bootstrap置信度,通过邻接连接构建系统发育树。从GenBank下载筛选到的PRRSV毒株的参考序列。通过Meg Align对获得的序列和71条ORF5参考序列进行核苷酸和氨基酸的同源性分析,探究核苷酸和氨基酸的变异模式。
1.6PRRSV GP5蛋白分析利用Net Nglyc 1.0 Server网站预测GP5蛋白中潜在的N-糖基化位点。
2 结果
2.1PRRSV ORF5基因的扩增通过RT-PCR检测692份临床样品,结果共获得368份阳性样品,阳性率为53.18%。对2016—2020年在华北地区的368份PRRSV阳性样本进行ORF5基因扩增,共有33份样品扩增出了目的条带,除HB-XT为600 bp外,其余均为603 bp(图1)。测序获得的33条PRRSV ORF5序列已上传至GenBank。
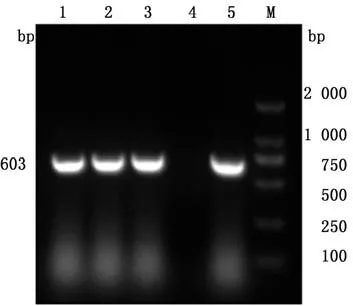
图1 部分PRRSV ORF5基因PCR鉴定结果
2.2ORF5基因遗传进化分析为研究PRRSV的遗传关系,本试验利用MEGA 7.0软件建立了ORF5基因遗传进化树(图2)。根据建好的进化树将104株(其中·代表分离的毒株)分为Ⅰ型和Ⅱ型,Ⅱ型又进一步分为9个谱系。本试验中的9株属于Lineage 8,其中包含高致病性代表性毒株JX-A1株和Hu N4株,以及疫苗株CH1-a和CH1-R,其余24株与美国JL580株和中国类NADC30株(CHsx1401)均属于Lineage 1。然而,北美型VR-2332和活疫苗株MLV RespPRRSV属于Lineage 5.1,野生型重组株GM2和QYYZ属于Lineage 3,本研究中获得的33株PRRSV均分布在Lineage 1和Lineage 8,据此推断近2~3年PRRSV流行毒株以NADC30株Lineage 1为主。核苷酸和氨基酸同源性分析结果显示,33株ORF5基因之间的核苷酸同源性为81.1%~99.8%,氨基酸同源性为80.1%~99.5%。氨基酸同源性分析发现,Lineage 8中9株之间同源性为92%~98.5%,而与CH1a和JXA1同源性分别为90.9%~95.4%和94.2%~99.8%。Lineage 1中的24株之间的氨基酸同源性为85.6%~99.5%,与NADC30毒株的同源性为91.7%~95%,而与CHsx1401株和JL580株的氨基酸同源性略低于NADC30毒株,分别为89.4%~94.7%和90.5%~93.9%。
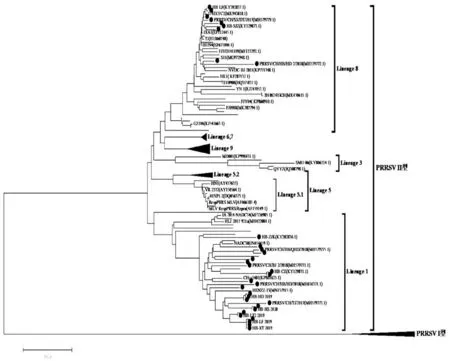
图2 PRRSV ORF5基因遗传进化树分析
2.3GP5蛋白氨基酸序列分析本研究获得的33条GP5蛋白氨基酸序列与参考序列比对(图3),结果显 示,与 参 考 毒 株(VR-2332、JXA1、HUN4、NADC30)相比,在信号肽区域(1~31aa)、跨膜区(TM)发生了不同程度变异。而在13位氨基酸有2种突变,且均发生在亚组3中:R→Q和R→P,其中包含11个Q13R突变和5个P13R突变。作为另一个重要的突变位点,151位氨基酸在Lineage 1中除HB-HS株均有R151K突变,而Lineage 8中无该位点突变。位于B表位(中和表位)中的多个位点均发生了突变,Lineage1中的6株(HB-LF2、HB-XT 2019、HB-SJZ 2020、HB-LF 2019、HB-XT 2018和HB-ZJK)发现了N38H突变,此外,39位氨基酸仅在Lineage 8中发生了与JX-A1株相同的I39L突变。同时在A表位(诱骗表位)中发现Lineage 8中大多数出现了A27V突变;而在28、29和30位氨基酸仅有少数出现了突变,HB-ZJK发现了独特的P28L突变;Lineage 8中HB-TS和HB-HS 2018株分别发现了D29V和A29V突变;HB-XT-2株出现了独特的E30N突变,HB-XT和HB-SJZ-2 2018和HBHS 2020株出现了S30N突变。GP5第137位氨基酸被认为可以区分野毒株与疫苗株的鉴别位点[13]。33株PRRSV在137位氨基酸均为S,不同于VR2332株和MLV Resp PRRS株的突变A137S,说明分离到的毒株均为野毒株。在67~90,107~119和138~150位氨基酸区域相对保守。

图3 PRRSV GP5氨基酸序列分析
2.4GP5蛋白糖基化位点分析通过Net Nglyc 1.0 Server网站预测并分析了GP5蛋白潜在的糖基化位点,结果显示,本研究中33株GP5蛋白中预测到了7个N-糖基化位点,分别为N30,N32,N33,N34,N35,N44和N51。如表1中显示,33株分别含有3~5个N-糖基化位点,且在7个位点中均有N44和N51位点。数量仅次于N44和N51位点的是N34和N30糖基化位点,分别有25个和23个位点。而Lineage 8中仅有1个N32位点,Lineage 1中有2个N32位点。就N33位点而言,Lineage 8和Lineage 1中分别有2个和17个位点。Lineage 1中的N-糖基化位点均无N35位点,Lineage 8中有6个N35位点。

表1 PRRSV GP5蛋白中的N-糖基化位点

续表1
3 讨论
2006年在江苏省暴发了HP-PRRS,并于2007年确认了高致病性变异毒株为引起该病的病原,随后该病在国内多呈现为地方性流,HP-PRRSV严重影响了中国养猪业[14]。先前的研究表明,HPPRRSV可能起源于中国经典PRRSV(类似CH-1a),并逐渐转变为HP-PRRSV毒株,在nsp2中不连续地缺失了30个氨基酸,导致了毁灭性病毒的产生。然而,后来的研究表明,这种独特的缺失与PRRSV的毒力无关[15]。尽管针对HP-PRRSV的商业MLV疫苗(例如,JXA1-P80,TJMF-92,R98和Hu N4-F112)被广泛使用,但是由于PRRSV基因组极易发生突变、重组和缺失等变异,引起抗原性发生改变,导致商品化疫苗的保护力下降,使得当前的控制策略未能提供可持续的疾病控制。同时,广泛使用MLV疫苗可能会增加猪群的免疫选择压力,从而加速PRRSV的变异和进化[16-17]。2013年,类似NADC30的PRRSV毒株在中国出现,这增加了PRRS在现场的复杂性。在随后的1~2年中,中国报道了许多NADC30样和中国HP-PRRSVs/VR-2332之间的重组菌株[16,18-19]。本实验室在2016—2020年收集到692份临床样品通过RTPCR检测,结果共获得368份阳性样品,阳性率为53.18%。PRRSV检出率较高,防控力度需要加大。
本试验共收集到华北地区368株PRRSV阳性样品,并用特异引物扩增出了33条ORF5基因序列,经进化树分析得出33株均为北美型,其中9株分布在Lineage 8(包含HP-PRRSV株-JX-A1,Hu N4株),24株分布在Lineage 1(包含NADC30株和类NADC30株)。同源性分析结果显示,33株北美型之间的氨基酸序列同源性为80.1%~99.5%,与JX-A1株和NADC30株的同源性分别为81.1%~99.5%和85.6%~94.5%。这意味着华北地区流行的毒株为HP-PRRSV株和类NADC30株,且在华北地区以类NADC30毒株流行为主,但分布在Lineage 1的24株核苷酸与氨基酸的同源性差距较大,这提示华北地区类NADC30毒株一直处于进化状态,且更加复杂,与此同时防疫意识仍然需要高度警惕。此外,2018年之前流行的HPPRRSV正逐渐被流行的类NADC30毒株所取代,并发展成为优势毒株,外部环境的不同PRRSV不可控因素可以通过不同途径侵袭猪场内部,使得猪场内部野毒株和变异毒株感染几率的风险加大。
GP5蛋白由ORF5基因编码,是PRRSV变异中最为明显的结构蛋白,它包括3个区域:N端信号肽区域(signal peptide)、诱骗表位(Decoy)、中和表位(PNE)、2个高变区(HVRs)和3个跨膜区(TM)[20],其中抗原决定簇27VLVN30和37SHLQLIYNL45作为主要的诱骗表位和中和表位,具有很高的变异性并且在诱导免疫应答中发挥重要作用[21]。而且,在PNE中H38、I42、Y43和N44的残基被认为是GP5的主要抗原识别位点,39~41位氨基酸可能有助于抗体结合。本试验结果表明,33株中除了HB-XT株在AA33中具有特定的缺失,其他毒株均未发现缺失。GP5蛋白的13、151位氨基酸(NADC30株:13Q和151K,VR2332株:13R和151R,MLV Resp PRRSV株:13Q和151G)被认为是与PRRSV毒力相关的关键残基。本试验中只有Lineage 1中11株有Q13R突变且与本谱系JL580株和NADC30株、Lineage 5.1 MLV Resp PRRSV疫苗株相同,而在Lineage 8中的13位氨基酸与JX-A1和Hu N4株均为R。同时,Lineage 1中在151位氨基酸均发生了R151K,而Lineage 8中未发生突变,这些结果证明了在华北地区发生的PRRS中存在着高致病性毒株和类NADC30毒株的流行。同时,我们还发现在A表位中发生了A27V、P28L、D29V和E30N突变,B表位中的38、39和40位氨基酸均发生了突变,尤其在39位氨基酸发生突变的数量较多且具有与JX-A1和Hu N4相同的突变。因此,我们推测位于中和表位中氨基酸发生突变,导致抗原性改变,使用不同毒株疫苗对华北地区流行的毒株中和效果不一致,这可能是疫苗未能对宿主提供有效保护力的原因。
GP5蛋白通常具有许多潜在的N-糖基化位点,病毒N-糖基化在病毒的毒性和存活中起着至关重要的作用,因为它促进了正确的蛋白质折叠,这被认为与病毒敏感性和宿主的免疫反应有关,起到帮助病毒从疫苗诱导的免疫反应中逃逸的作用[22]。因此,确定具有保守基序6Asn-X-Ser/Thr(N-X-S/T)的潜在糖基化位点。有研究报道北美基因型GP5蛋白共有4个N-糖基化位点(N30、N34、N44和N51),且位点的突变容易降低PRRSV中和抗体的水平进而增强了PRRSV宿主体内的增殖。本研究对PRRSV GP5蛋白氨基酸全序列进行了糖基化位点的预测,分析了位于抗原表位中的N-糖基化位点。结果表明,33株PRRSV预测得到了7个N-糖基化位点,且不同株潜在的位点和数量均不同,分别含有3~5个位点。作为GP5蛋白的保守N-糖基化位点,在本试验中的N44和N51位点没有发生缺失和突变,这与之前的报道N44和N51位的糖基化信号高度保守相一致。然而,其余5个糖基化位点在不同位置均发生了突变,仅HB-XT在N33位点发生了缺失,但不同氨基酸位点的缺失对糖基化位点的预测影响不大。本试验推测出的N-糖基化位点主要位于抗原表位中的糖基化位点,不同数量及位点可能会减弱中和抗体表位的免疫原性。这些结果提示,在华北地区PRRSV中,由于氨基酸的缺失和突变可能导致了抗原性改变,但其生物学功能仍需进一步研究,研究PRRSV GP5蛋白N-糖基化位点对病毒感染能力以及免疫原性具有重要意义。
类NADC30传入我国数年间,因其较强的基因变异能力,加之与本土毒株的基因重组能力也较强,迅速发展成为我国猪场中的优势毒株,而商品化类的疫苗对其猪场的免疫保护能力总是不尽如人意,使得我国猪场对PRRSV防控难度增强。而试验通过RT-PCR对2016—2020年收集的华北地区692份组织进行阳性样品的筛选,并对ORF5基因进行扩增和测序。从遗传进化分析、序列相似性分析、GP5蛋白氨基酸分析3个方面分析了华北地区PRRS遗传特点和流行情况。根据遗传进化和相似性分析表明本研究获得的33株均为Ⅱ型。此外,根据GP5蛋白氨基酸序列比对发现,氨基酸位点突变导致毒力不同,可以得出在华北地区流行的毒株以类NADC30为主,同时还存在少数高致病性毒株的流行。本研究从分子水平分析了华北地区PRRSV的流行特征,这些结论有助于制定具有针对性的防控措施和进行新疫苗的研发。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
中国农业科学(2022年15期)2022-08-09
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
环球时报(2021-07-12)2021-07-12
电脑报(2020年40期)2020-11-06
电脑知识与技术(2018年19期)2018-11-01
山东农业科学(2017年2期)2017-03-15
科技与创新(2016年12期)2016-06-25
建材发展导向(2014年5期)2014-10-20
