
Environmental variables affecting the gillnet catches and condition of Labiobarbus festivus and Osteochilus hasseltii in northern Malaysia
2022-03-01 03:32:28MustafizurRahmanAhmadFathiMattBroadhurst
Aquaculture and Fisheries 2022年6期
Mustafizur M.Rahman, Ahmad Fathi, Matt K.Broadhurst
aDepartment of Marine Science, International Islamic University Malaysia (IIUM), Jalan Sultan Ahmad Shah, Kuantan, Pahang, 25200, Malaysia
bInstitute of Oceanography and Maritime Studies, IIUM, Kg.Cherok Paloh, Kuantan, Pahang, 26160, Malaysia
cDepartment of Biotechnology, IIUM, Jalan Sultan Ahmad Shah, Kuantan, Pahang, 25200, Malaysia
dNSW Department of Primary Industries, Fisheries Conservation Technology Unit, National Marine Science Centre, Southern Cross University, 2 Bay Drive, Coffs Harbour, 2450, Australia
eMarine and Estuarine Ecology Unit, School of Biological Sciences, University of Queensland, Brisbane, 4072, Australia
Keywords:
Redundancy analysis
PERMANOVA
Growth co-efficient
Water turbidity
Phytoplankton abundance
Benthos
Lake Kenyir
A B S T R A C T
The influences of technical factors on the catching efficiencies of gillnets are well understood, but less is known about the importance of environmental factors and how these might concurrently affect target-species condition.Here we address this deficit for two economically important southeast Asian cyprinids (Labiobarbus festivus and Osteochilus hasseltii) during a one-year study at a key fishing location in Lake Kenyir, Malaysia.Three gillnets(each 200 m long, 2 m deep and comprising either 38-, 51-, or 76-mm mesh) were deployed each month concurrent with sampling of benthic macroinvertebrates and various environmental parameters.Various multivariate analyses (detrended correspondence analysis, redundancy analysis and permutational multivariate analysis of variance) were used to explore potential relationships between the extrinsic variables (mesh size,sampling season, water quality and, based on known prey items, benthic macroinvertebrates) and gillnet catches and the biological condition (growth co-efficient and hepatosomatic index) of the target species.Catches of L.festivus (the most abundant species) were positively influenced by water turbidity without seasonal effects, while their biological condition was positively influenced by benthic macroinvertebrates (mostly Trichoptera) and the concentrations of DO and phosphate, and negatively influenced by water temperature.By comparison, seasonal influences on the catches and biology of O.hasseltii were somewhat size specific with positive and negative effects of the monsoon on small and large fish, respectively.The abundance of phytoplankton also positively affected the catches of O.hasseltii, while their biological condition was positively influenced by water temperature, but negatively influenced by water turbidity.Such species-specific differences were attributed to life histories, and although the more abundant L.festivus might be best sought during any period of turbidity, effort should be focused during the monsoon when individuals have optimal condition (and therefore better flesh quality).In contrast, larger O.hasseltii might be best sought during non-monsoon months concurrent with greater catches and optimal condition.Collecting similar data for other freshwater species both nationally and internationally might facilitate future efforts at fine-tuning gillnet fishing effort.
1.Introduction
Gillnets are widely used throughout the world, and are favoured in artisanal fisheries because they are relatively inexpensive, easily constructed and deployed and quite effective for various species, especially schooling teleosts (Hamley, 1975; Brandt, 2005; Yalei et al., 2018;Grimaldoa et al., 2019).However, like all fishing gears, gillnets are not 100% selective for the target species (Broadhurst, Kennelly, & Gray,2007).Rather, they typically have wide selectivities among similar sizes/shapes of animals, which means at times they also catch unwanted species.To promote sustainable harvesting, ideally gillnets would be configured and fished across appropriate spatio-temporal scales that maximize catches of the permitted species and their sizes.
Various extrinsic and intrinsic factors are known to affect gillnet selectivities and/or efficiencies (reviewed by Hamely, 1975).Key technical parameters can include, but are not limited to: soak time (Minns &Hurley, 1988; Losanes, Matuda, & Fujimori, 1992); and net length(Rudstam, Magnuson, & Tonn, 1984), hanging ratio (defined as the length of stretched meshes divided by the length of rope to which they are attached; Acosta & Appeldoorn, 1995), twine size, material (Jensen,1995; Hovgard, 1996a), headline height (Gray, Broadhurst, Johnson, &Young, 2005), panel configuration (Gilman, Gearhart, Price, Eckert,Milliken, Wang, Swimmer, Shiode & Abe, 2010), and perhaps most importantly, mesh size (Hamley, 1975).
In addition, many environmental and biological parameters can affect species catchability in gillnets.Specifically, beyond short-term diel effects, seasonal influences are often quite important and can directly and indirectly influence the biology or behaviour of many fishes and their subsequent vulnerability to capture (Bobori & Salvarina, 2010;Gray, Johnson, Broadhurst, & Young, 2005).Such broad temporal effects are often linked with the interacting effects of important abiotic(water temperature, dissolved oxygen, turbidity and the concentration of various nutrients) and especially biotic factors such as food availability (Hansson & Rudstam, 1995).Some of these factors probably manifest as quite variable species-specific impacts, although according to Stoner (2004), ambient prey density has the greatest influence on catchability.
Despite recognition of the many environmental and biological factors affecting gillnet catches, most previous studies investigating the importance of key technical factors such as mesh sizes or other gear configurations on relative efficiencies/selectivities have been done over quite short temporal scales (Acosta & Appeldoorn, 1995; Gray, Johnson,et al., 2005; Hovgard, 1996b).Such studies often incorporate random effects in analyses, including different fishing days and locations, which implicitly encompass some variables likely to affect gillnet fishing performances (usually across multiple mesh sizes).But, by default these approaches preclude deciphering the range of potentially important environmental and biological factors listed above, which means data are lacking for many species.
This study sought to contribute towards addressing the above stated shortfall in information by investigating the influence of key extrinsic variables on the gillnet vulnerability (catches) and condition (growth coefficient and hepatosomatic index; HSI) of two economically important Malaysian cyprinid fish (Labiobarbus festivusandOsteochilus hasseltii)across a full annual fishing season.Both species are targeted using gillnets (~51-mm stretched mesh opening {SMO} monofilament)appropriately configured for the desired sizes (~16-18 cm total length-TL) throughout their southeast Asian distributions and mostly for regional consumption, especially in Malaysia.These two species have variable distributions, but previous ecological research suggestsL.festivustypically consumes macroinvertebrates, such as insect larvae along with detritus, whileO.hasseltiiis mostly planktivorous, although larger individuals also consume invertebrates (Nurulnajwa, 2004; Kottelat & Widjanarti, 2005).Variations in the relative availability of these preferred prey items should affect relative abundances and therefore vulnerability to gillnets, but no data are available.
Considering the above, our first aim here was to test the null hypothesis of no effects of a suite of environmental variables on the gillnet catches and biological parameters ofL.festivusandO.hasseltiiin a key Malaysian fishery (at Lake Kenyir which, with a surface area of 38,000 ha, is the largest man-made waterway in Malaysia).Using this information, our second aim was to then identify any conditions (or otherwise) that might be amenable for future regional harvesting, and ultimately to facilitate the broader, effective exploitation of these species.
2.Materials and methods
2.1.Study site and fishing gear
The study was done in Lake Kenyir (5°3′′N; 102°44′′E), northeastern Malaysia between January and December 2016 (Fig.1).At this location,the monsoon and non-monsoon periods are from October to March and April to September, respectively.Three gillnets were constructed for the study: (1) a traditional configuration (51.0 mm SMO) and those with (2)larger (76.2 mm SMO) and (3) smaller (38.1 mm SMO) mesh sizes (to ensure adequate sampling of fish populations).All three gillnets were made from clear polyamide monofilament mesh (0.2-mm diameter-Ø twine) throughout, attached to 200-m floatlines (4.0-mm Ø twisted polypropylene) and suitably weighted footropes (3.0-mm Ø twisted polypropylene) at a hanging ratio of ~50% (Fig.2).Each gillnet orientated to a fishing depth of ~2 m (Fig.2).
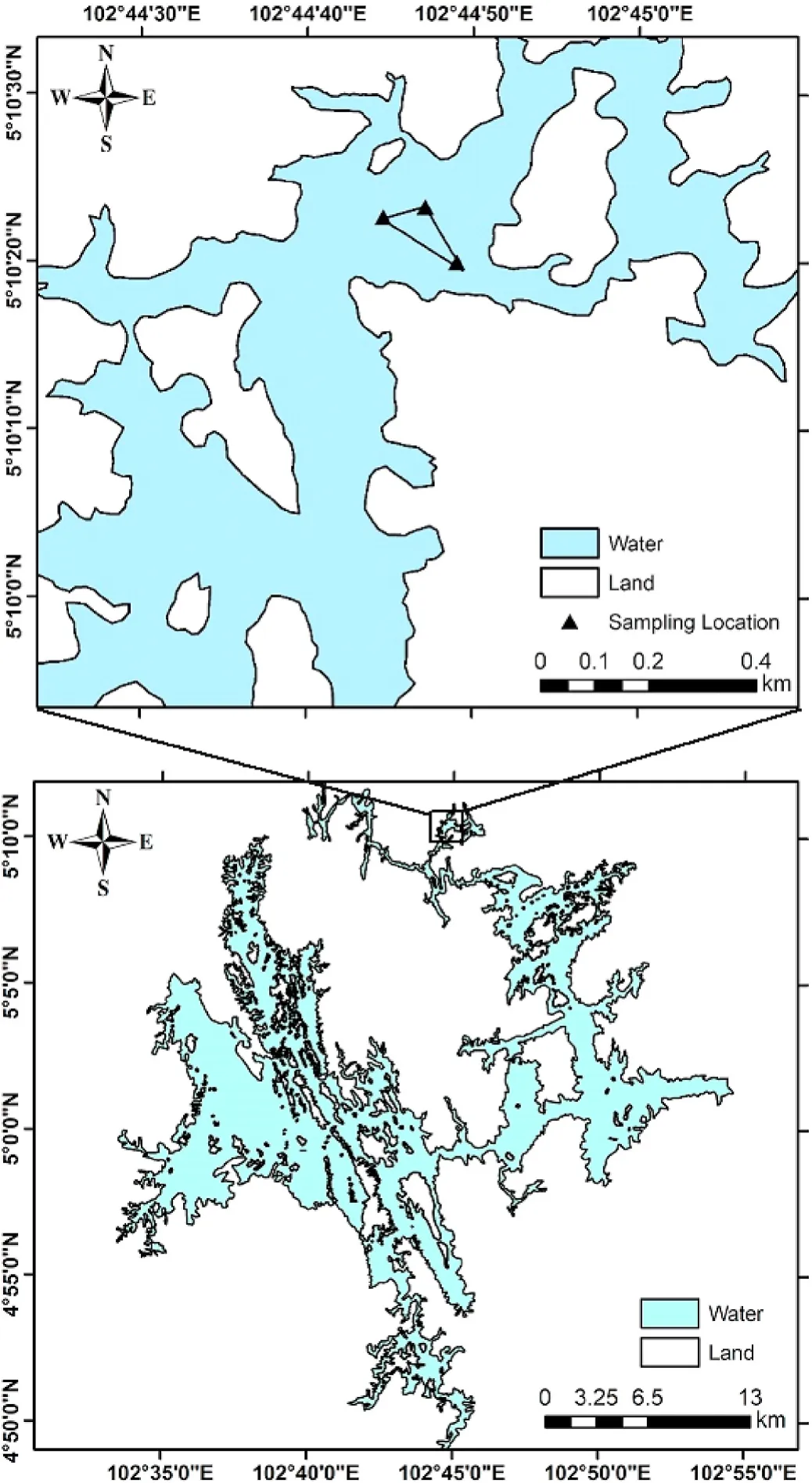
Fig.1.Map of Lake Kenyir, Malaysia and the designated fished area between January and December 2016.
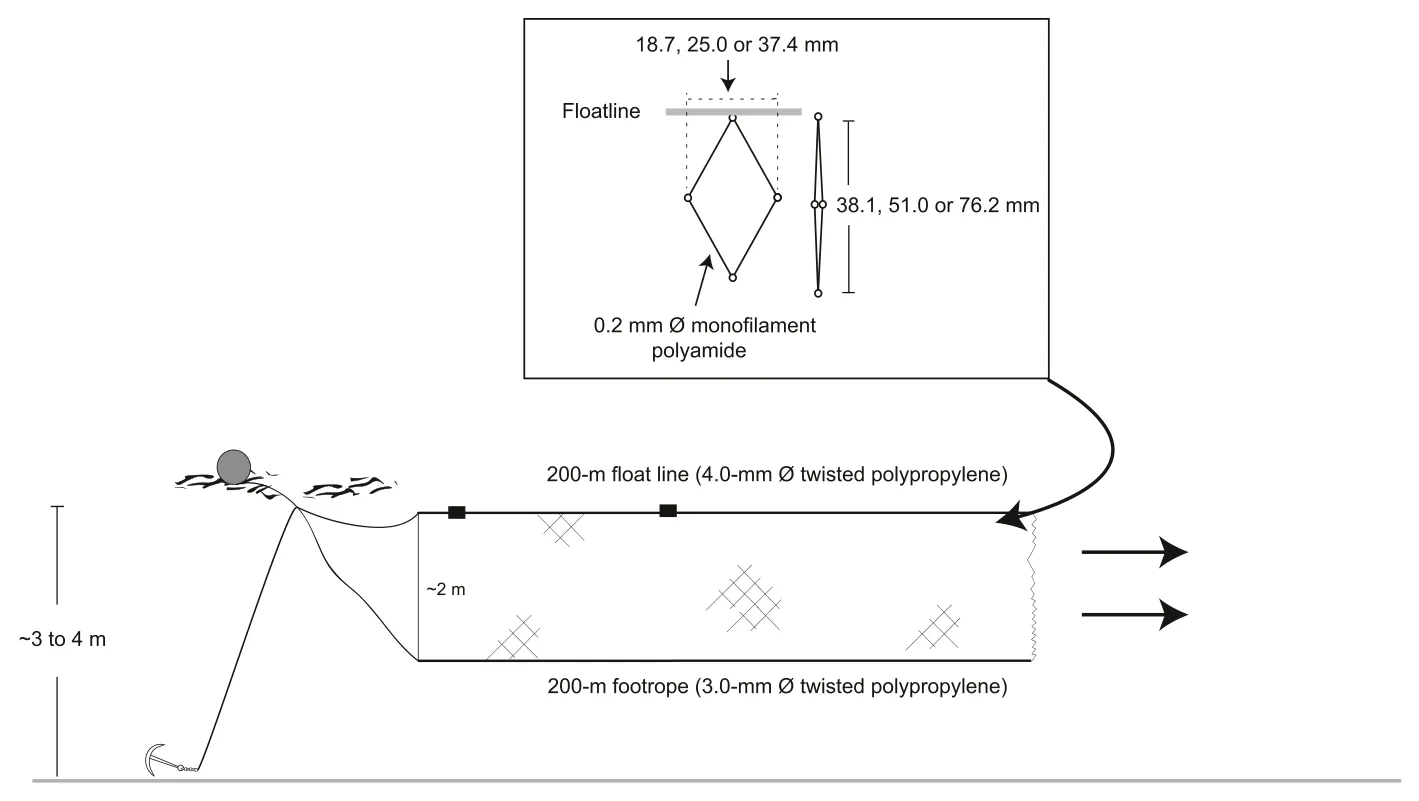
Fig.2.Configurations of three gillnets (comprising 38.1, 51.0 and 76.2 mm stretched mesh opening) monthly deployed in Lake Kenyir, Malaysia between January and December 2016.Ø, diameter.
The gillnets were always fished across the same defined location(depths of 3.1-4.3 m) in the Lake and in an area where conventional fishing effort was known and considered constant (Fig.1).The chosen location was monthly sampled over 12 months (to incorporate a full seasonal cycle).Three replicate-gillnet deployments were completed each month.
2.2.Fish and benthos sampling and associated water quality
During each fishing night, the gillnets were deployed at the surface at 17:00 h and retrieved at 01:00 h.After retrieval, all fish were immediately untangled and counted.Specimens ofL.festivusandO.hasselttiiwere separated, stored in ice and transported to the International Islamic University Malaysia (IIUM) where their total length (TL) (to the nearest 1 mm) and weight (TW to the nearest 0.01 g) were immediately recorded before they were dissected and their livers removed for weighing(LW as above).The growth co-efficient (parameterbin the regressionW=aLb) and the hepatosomatic index (HSI =LW/TW ×100) for each specimen were subsequently calculated following conventional procedures (Pauly, 1993).
Concurrent with gillnet deployments, and within 100 m of each deployment, replicate substrate samples were collected using a Ponar grab (15.2 × 15.2 cm).Samples were washed through a 250-μm mesh sieve and all non-filtered benthos were immediately preserved in plastic vials containing a 10% buffered formalin solution and transferred to IIUM for quantification.Organisms were identified to at least their Class following keys developed by Brinkhurst (1971) and Pinder and Reiss(1983).
Replicates of water temperature (oC), dissolved oxygen (mg l-1),water turbidity (NTU) and pH (-logaH+) were also recorded at the~mid-fishing depths of the gillnets using Hydrolab™ monitors(Hydrolab Minisonde water-quality multiprobes).Additionally, water samples (~1 l) were collected at the surface, mid and bottom depths using a Van Dorn™ water sampler and cumulatively assessed for nitrogenous and phosphorous nutrients, silica (mg l-1) and chlorophylla (μg l-1), with the latter selected because it is indicative of phytoplankton biomass (Jenkerson & Hickman, 2007).
Chlorophyll-a was determined spectrophotometrically after acetone extraction according to Boyd (1979).Total ammonia nitrogen (TAN)and phosphorus (PO4-P) were analysed spectrophotometrically following Stirling (1985).Nitrate (NO3-N; derived via the cadmium reduction method) and nitrite (NO2-N) were determined according to APHA (1998) and along with TAN, were used to calculate total dissolved inorganic nitrogen (TIN).
2.3.Statistical analyses
Data describing the extrinsic variables (e.g.seasons separated into monsoon and non-monsoon periods, water quality measures, pH, TIN,phosphate, chlorophyll-a, silica, and abundant benthic macroinvertebrates) and the intrinsic biology (growth co-efficient and HSI)and gillnet vulnerability (catches) of each species for each mesh size were first explored using the canonical correlation index.The waterquality variables were subsequently designated as explanatory (independent) variables while the biology and gillnet vulnerability of fish were considered responses (dependent) for analyses in multivariate ordinations using CANOCO 4 (ter Braak & Smilauer, 1998).
Initially, a detrended correspondence analysis (DCA) was performed to elucidate prevailing patterns of the response variables relative to the explanatory variable gradient.The subsequent ordination axes were smaller than two standard deviations, implying a linear response, and so redundancy analyses (RDA) was deemed appropriate following ter Braak & Smilauer (1998).The RDAs were run separately for the two species with variables centered and standardized by subtracting the mean and dividing by the standard deviation.The significance of the first-ordination axis and of the first four canonical axes together were evaluated via Monte Carlo-permutation tests with 1000 permutations.
Following the results of the RDA, permutational multivariate analysis of variance (PERMANOVA) based on Euclidian distance was performed to investigate the main categorical effects of season (monsoon vs non-monsoon) and ‘mesh size’ of the gillnets and their interaction on various intrinsic variables for both species.We also used PERMANOVA with the DistLM model to investigate the influence of environmental factors on species biology and gillnet vulnerability.The PERMANOVA analyses were performed using PRIMER v7 (Anderson, 2008), while both DCA and RDA were completed using CANOCO v4 (ter Braak &Smilauer, 1998).
3.Results
In total, 4430 fishes comprising at least 18 species were caught, withL.festivusandO.hasselttiiaccounting for 48 and 13%, respectively of the total catches (Table 1).These catches were mostly taken in the 38.1-mm(total of 1519; and 78% and 22%L.festivusandO.hasselttii) and 51.0-mm gillnets (1162; 79% and 21%), with only 25 fish recorded in the 76.2-mm gillnet.The later gillnet was excluded from any further analyses.The remaining fish species mostly comprised cypriniformes.
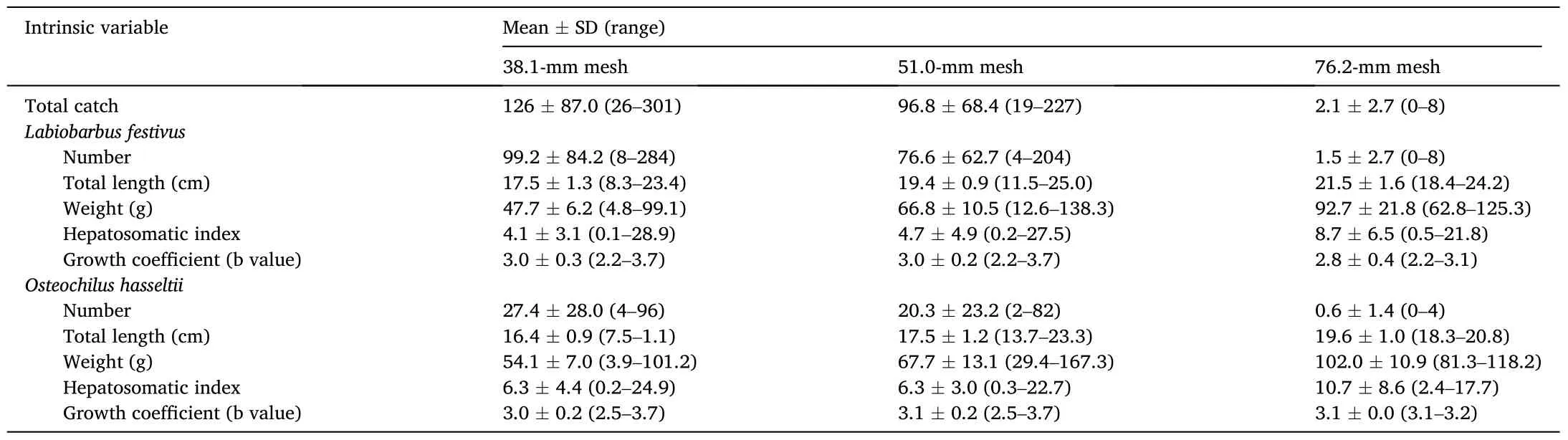
Table 1Summary of means ±SD and ranges of catches and biology of Labiobarbus festivus and Osteochilus hasseltii during the 12-month study fishing gillnets made from three mesh sizes (stretched mesh opening) in Lake Kenyir, Malaysia between January and December 2016.
The sampled benthic organisms (total benthic macroinvertebrates;TBM) belonged to 10 classes, including polychaete, oligochaete, insecta,hirudiena, bivalvia, arachnida, isopoda, copepoda, mysida and gastropoda.However, 62% of organisms were Trichoptera larvae.The collected environmental parameters were all annually variable across the entire fished area, but especially chlorophyll-a and turbidity(Table 2).

Table 2Summary of means ±SD and ranges of various extrinsic variables across the fishing location of gillnets deployed monthly in Lake Kenyir, Malaysia between January and December 2016.
3.1.Labiobarbus festivus
For the RDAs describing relationships between environmental variables and the catches and biology ofL.festivus, the first canonical axis and the first four canonical axes combined were statistically significant(P <0.05; Table 3).The first two canonical axes explained 82.5% of the variance in catches and 93.3% of the catches/environmental relationship (Table 3).Monsoon season scored high on the second RDA axis(Fig.3).For both the 38.1-mm and 51.0-mm mesh gillnets, the second RDA was positively correlated with several environmental variables(excluding turbidity) and fish biology variables, and was negatively correlated with non-monsoon, water temperature, pH, TIN and chlorophyll-a (Fig.3).
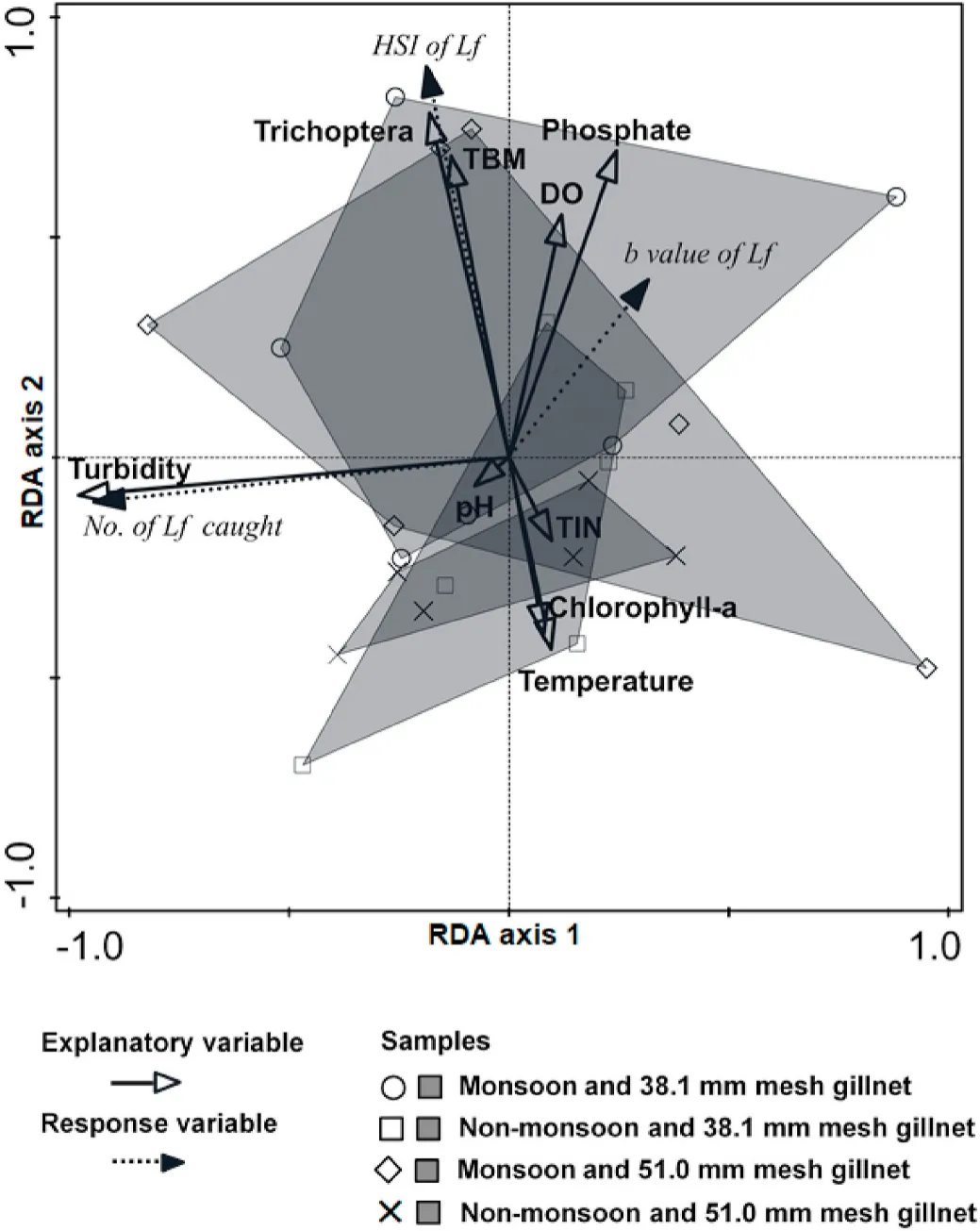
Fig.3.Redundancy analyses plots of environmental variables explaining the gillnet (38.1- and 51.0-mm mesh) catches and biology of Labiobarbus festivus in Lake Kenyir during 12-months’ sampling between January and December 2016.Lf, Labiobarbus festivus; TIN, total dissolved nitrogen; HSI, heptosomatic index;DO, dissolved oxygen; TBM, total benthic macroinvertebrates.
The PERMANOVA assessing the categorical effects of season and mesh size failed to return a significant pseudo-F for either main effect or their interaction on the number ofL.festivuscaught, or their growth coefficient (Table 4).However, there were significant effects of season on HSI (means ± SDs of 7.44 ± 4.94 and 2.61 ± 2.51 for monsoon and nonmonsoon seasons, respectively), and mesh size on the TLs and weights of fish, which manifested as incrementally greater sizes and weights with increasing mesh size (P <0.05; Table 1 and 4).
Irrespective of mesh size, PERMANOVA (with DistLM) revealed that water turbidity strongly positively influenced the number ofL.festivuscaught (P <0.05; Fig.3, Table 5).And the HSI of fish was significantly and positively influenced by benthic macroinvertebrates, DO and phosphate and negatively influenced by water temperature (P <0.05;Fig.3, Table 5).Only phosphate and TIN significantly influenced the growth coefficient ofL.festivus, and with positive and negative effects,respectively (P <0.05; Fig.3, Table 5).
3.2.Osteochilus hasseltii
The first canonical axis and the first four canonical axes combined were also statistically significant forO.hasseltii(P <0.05; Table 3).The first two canonical axes explained 70.1% of the variation in catches and 92.9% of the catches/environmental relationship (Table 3).It was clear that fishing with the 38.1-mm gillnet during the monsoon season, and both gillnets during the non-monsoon scored high on the first RDA axis(Table 3; Fig.4).This axis was strongly positively correlated with the number of fish caught and chlorophyll-a, and negatively correlated withthe phosphate concentration in the water (Fig.4).
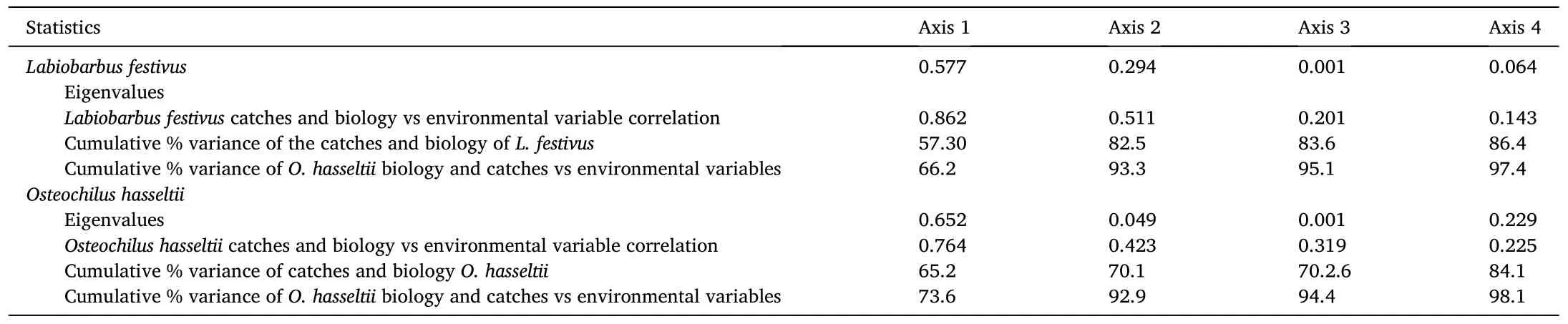
Table 3Redundancy analyses (RDA) of environmental variables explaining the biology and catches of Labiobarbus festivus and Osteochilus hasseltii in gillnets deployed monthly in Lake Kenyir, Malaysia between January and December 2016.
The observed influence of the smaller mesh on catches ofO.hasseltiiduring the monsoon season manifested as a significant interaction between mesh and season in the PERMANOVA (P <0.05; Table 4).However, irrespective of season and like forL.festivus, mean TL and weight were greater in the larger mesh size (P <0.05; Table 1 and 4).Neither the growth co-efficient nor HSI ofO.hasseltiiwere affected by mesh size or season (P >0.05; Table 1 and 4).
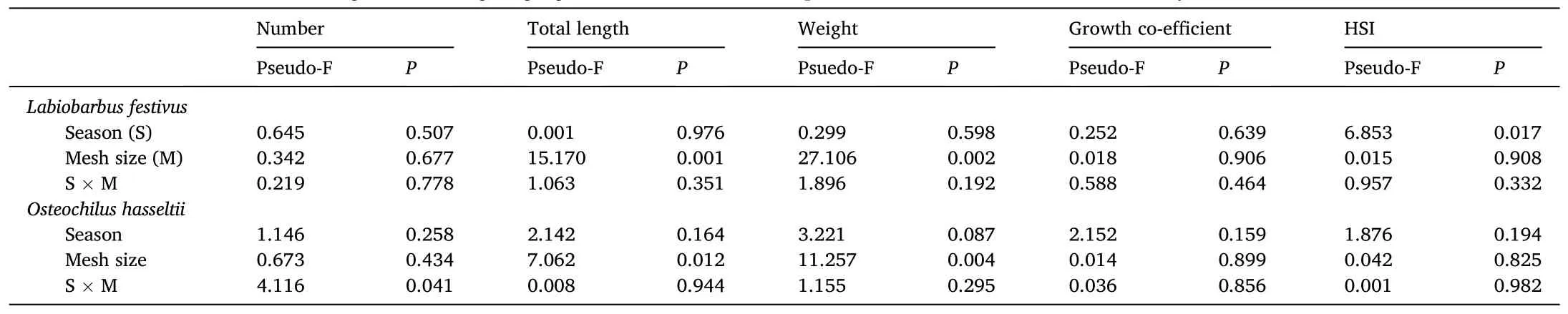
Table 4PERMANOVA pseudo-Fs and significance for influence of season (monsoon vs non-monsoon), mesh size (31 vs 51 mm mesh; see Table 1 for means) and their interaction on the number, total length (cm), weight (g), growth co-efficient and hepatosomatic index (HSI) of Labiobarbus festivus and Osteochilus hasseltii.
PERMANOVA (with DistLM) revealed the number ofO.hasseltiicaught was significantly and positively influenced by the abundance of phytoplankton (chlorophyll-a concentration) (P <0.01), but was not significantly influenced by the phosphate concentration in the water (P>0.05; Fig.4, Table 5).The growth coefficient and HSI were positively influenced by water temperature and negatively influenced by water turbidity (P <0.05; Fig.4, Table 5).While other environmental variables (phytoplankton, Trichoptera and TBM densities, TIN and phosphate concentration and pH) slightly influenced the growth coeffi cient and HSI ofO.hasseltii, their influences were not statistically significant (P >0.05; Fig.4, Table 5).
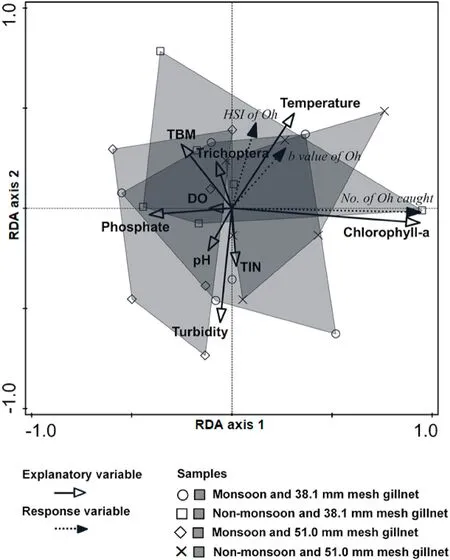
Fig.4.Redundancy analyses plots of environmental variables explaining the gillnet (38.1- and 51.0-mm mesh) catches and biology of Osteochilus hasseltii in Lake Kenyir during 12-months’ sampling between January and December 2016.Oh, Osteochilus hasseltii; TIN, total dissolved nitrogen; HSI, heptosomatic index;DO, dissolved oxygen; TBM, total benthic macroinvertebrates.
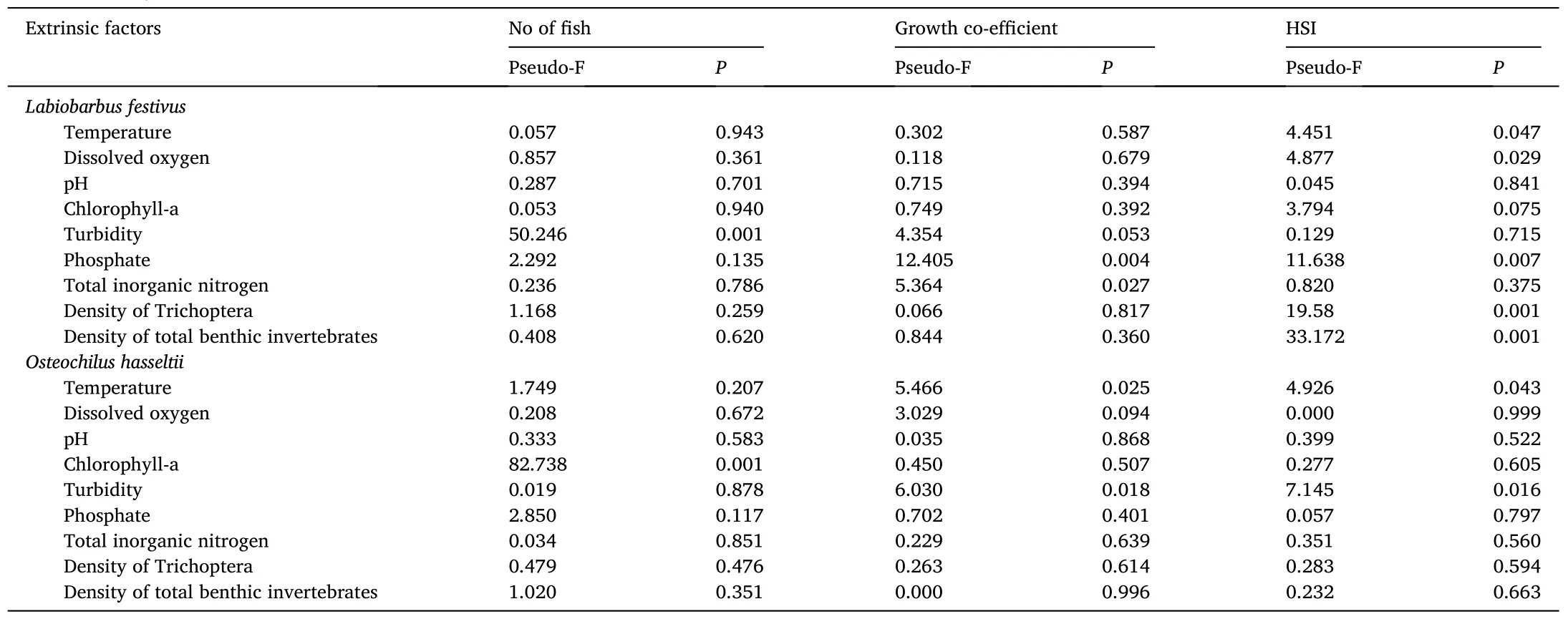
Table 5PERMANOVA (with DistLM) pseudo-Fs and significance for the influence of various extrinsic factors on the catches, growth co-efficient and hepatosomatic index (HSI)of Labiobarbus festivus and Osteochilus hasseltii (irrespective of mesh size and season).
4.Discussion
This study provides a unique contribution towards understating important factors affecting gillnet efficiency/selectivity (e.g.Hamley,1975; Rudstam et al., 1984; Minns & Hurley, 1988; Losanes et al., 1992;Acosta & Appeldoorn, 1995; Jensen, 1995; Hansson & Rudstam, 1995;Hovgard, 1996a,b; Poulsen, Nielsen, Holst, & Sather, 2000; Gray,Johnson, et al., 2005; Gilman et al., 2010).Specifically, by sampling across a full 12-month cycle using three different-meshed gillnets in one fishery and collecting a plethora of environmental data, we have delineated species-specific variability among the relative importance of key variables such as food availability, water turbidity, temperature, DO and nutrients affecting the catches ofL.festivusandO.hasseltiiand also their biology.
We acknowledge that these variables are probably not the limit of causality among observed catches-which could extend to the other factors affecting gillnet vulnerability (e.g.abundances of other fish species or effort by fishers) and should be assessed in future research.Nevertheless, the observed influences provide important information,which can be discussed according to possible life-history requirements for the studied species, and used to not only regulate regional effort, but also contribute towards fisheries management more broadly.
The environmental dataset adequately explained variations among catches and the biology of each species quite well for each of the two mesh sizes with sufficient data, but clearly with divergent speciesspecific importance.Like for several other species, the gillnet catches ofL.festivuswere strongly affected by water turbidity (Balik & Cubuk,2000; Simasiku, Mafwila, & Sitengu, 2017).Such a result might reflect their reliance on visual stimuli considering this species generally resides in clear water and might not be proficient at detecting gillnets as turbidity increases (Kottelat & Widjanarti, 2005).Other factors, such as seasonal variation, did not affect the growth co-efficient and vulnerability ofL.festivusto capture, but did affect their HSI, which was strongly positively influenced during the monsoon, and negatively influenced during the non-monsoon season for both gillnets.Specific variables influencing HSI within these broad seasonal effects included water temperature, DO and phosphate concentrations and the abundance of benthic macroinvertebrates (dominated by Trichopterans).Considering the latter, one hypothesis to explain the positive effect on the HSI ofL.festivusmight be their dietary preferences (Amira et al., 2016; Rahman et al., 2009; Rahman & Balcombe, 2018), which although not studied here are known to be strongly directed towards insect larvae and detritus (Kottelat & Widjanarti, 2005).
The remaining environmental factors (chlorophyll-a, as a measure of phytoplankton density, pH and turbidity) only slightly affected the biology ofL.festivus, and were not significant.This outcome may reflect Liebig’s law of the minimum whereby fish energetics are influenced by the combination of numerous factors, but for many species, are mostly controlled by the scarcest resources such as DO or food availability(Gibson, 1994; Hiddink & Kaiser, 2005).The observed negative relationship between water temperature and the biology ofL.festivusimplies the monsoon season is optimal for the growth of this species.Similar relationships have been recorded for various other teleosts in lakes and reservoirs (e.g.Kocovsky & Carline, 2001; Quist, Guy, Bernot, & Stephen, 2002).
Unlike forL.festivus, the catches ofO.hasseltiiwere greatly influenced by the abundance of phytoplankton in lake water.Catches ofO.hasseltiiwere mostly greater during the non-monsoon season and probably due to changes in habitat selection and increased activity during the former seasons.A similar relationship between gillnet catch rates and the non-monsoon season has been observed for other species,includingPerca fluviatilisin Lake Doirani in Greece (Bobori & Salvarina,2010).
Notwithstanding some effect of the non-monsoon season, there was a clear size-specific influence of the monsoon onO.hasseltii(evidenced via the PERMANOVA interaction between mesh size and season) which manifested as a negative influence on the catch and biology of largerO.hasseltii(as sampled by the 51.0-mm gillnet).While speculative, such a result may have reflected food and habitat preferences.According to Nurulnajwa (2004), smallO.hasseltiiare mostly planktivorous, while larger individuals feed on phytoplankton, zooplankton, aquatic invertebrates and detritus.At the onset of the monsoon, largerO.hasseltiiare known to migrate into inundated areas for feeding (mainly aquatic invertebrates) and spawning (Motomura, Tsukawaki, & Kamiya, 2002).Such size-specific shifting habitat and food preferences could explain the negative influence of monsoon on catches ofO.hasseltiiin the larger-meshed gillnet.
We also observed that water temperature and turbidity positively and negatively influenced the growth coefficient and HSI, respectively ofO.hasseltiiwithout any clear seasonal influence.UnlikeL.festivus,water turbidity may not greatly affect the visionO.hasseltii, but could affect their biology (Table 5).Negative effects of turbidity on capture and biology are commonly observed in many freshwater fishes (Pekcan-Hekim.2007; Pekcan-Hekim & Lappalainen, 2006; Rahman &Verdegem, 2007).Considering the improved condition ofO.hasseltiiduring the non-monsoon period (and with significantly warmer temperatures than during the monsoon period), one hypothesis might be that this correlates better with the preferred temperature range,although further research is required to validate this hypothesis.
5.Conclusions
The data collected here have implications for optimal fine-scale harvesting of both species.In terms of absolute abundances, forL.festivusthe lack of any seasonal effect means the species could be sought during any period of turbid waters throughout the year.But,becauseL.festivusdemonstrated optimum biological condition (which by default might imply better flesh quality and therefore economic return) during the monsoon season, directed effort during this period would be advantageous in terms of maximizing profits for local fishers.In contrast, largeO.hasseltiiwould best be targeted during the nonmonsoon using the conventional gillnet, simply because the species demonstrated optimum biological condition during this period.For other areas of the species’ distributions, the observed relationships between extrinsic environmental factors and gillnet vulnerability could be used to refine targeting, although additional data would be required.
The existing data also have implications for fishery-independent sampling and potential confounding effects.In particular, sporadic episodes of increased turbidity could confound time-series data forL.festivus, but notO.hasseltii.Such factors require consideration for effective stock assessments and to decipher trends during anomalous annual climatic events.Similar characteristics could affect the vulnerability of other freshwater species to gillnets elsewhere and warrant ongoing assessments.In this regard, our data support broader, more holistic evaluations of extrinsic factors affecting the gillnet vulnerability of targeted fish.
CRediT authorship contribution statement
Mustafizur M.Rahman: Funding acquisition, Methodology, Supervision, Data curation, Software, Writing - original draft, Formal analysis, Visualization, Writing - review & editing.Ahmad Fathi: Data curation.Matt K.Broadhurst: Review and editing.
Declaration of competing interest
No author has any conflict of interest with the contents of the manuscript.
Acknowledgments
We are thankful to the International Islamic University Malaysia,Malaysia for providing financial support through P-RIGS18-032-0032 project.We also thank the Malaysian Agricultural Research and Development Institute (MARDI) for logistical support during the study, and the NSW Department of Primary Industries for the involvement of Matt Broadhurst.
 Aquaculture and Fisheries2022年6期
Aquaculture and Fisheries2022年6期
- Aquaculture and Fisheries的其它文章
- Impact of anthropogenic activities on changes of ichthyofauna in the middle and lower Xiang River
- Comparison for ecological economic performance of Chinese sea perch(Lateolabrax Maculatus) under different aquaculture systems
- Genetic diversity and relatedness inferred from microsatellite loci as a tool for broodstock management of fine flounder Paralichthys adspersus
- Retention of fin clips and fin and operculum punch marks in rainbow trout
- Preliminary data of life history traits of Mormyridae (Actinopterygii:Teleostei) in the Upper Sanaga River, Central Region of Cameroon
- Hybrids production as a potential method to control prolific breeding in tilapia and adaptation to aquaculture climate-induced drought