
氮素水平及形态对娃娃菜根系特征及生理指标的影响
2022-02-24 05:20:28马玉峰周忠雄李雨桐高雪琴乔亚丽张文斌颉建明胡琳莉郁继华
中国农业科学 2022年2期
马玉峰,周忠雄,李雨桐,高雪琴,乔亚丽,张文斌,颉建明,胡琳莉,2✉,郁继华,2✉
1甘肃农业大学园艺学院,兰州 730070;2甘肃省干旱生境作物学重点实验室,兰州 730070;3湖北凯龙楚兴化工集团有限公司,湖北钟祥 431900
0 引言
【研究意义】氮是植物生长发育的基本元素,植物以复杂反应从生理和形态水平响应氮素供应的变化,植物器官依赖于局部和远距离的信号通路来协调整个植物水平的反应,激素等是这些途径的一部分信号物质。植物生长发育过程中许多代谢活动的信号调节离不开氮素,植物以根系吸收同化利用,根系生长发育对植物养分和水分的吸收有很大的影响作用,并影响植物地上部分的干物质重量[1-2]。因此,研究氮素参与娃娃菜根系形态塑造的作用机理,可以为进一步研究氮素调控根系形态的分子机制奠定基础。【前人研究进展】氮素水平和形态影响植物根系生长发育,低浓度氮素会刺激根伸长和侧根发育,而高浓度氮素会对根生长产生抑制作用[3-4]。局部供铵可以促使维管系统中地上部 IAA积累并影响拟南芥侧根伸长,而根尖部分积累的 IAA决定侧根的发生,局部供硝可以促进一级、二级侧根的分支和伸长。IAA在根的伸长和侧根起始中占主导地位,CTK可以协同IAA调控植物根系生长,而SA、ETH与ABA均表现为拮抗 IAA以达到根系调控的作用,其中乙烯会刺激植物生长素合成,抑制细胞延伸[5-8]。JA抑制植物幼苗叶片及主根伸长,又能通过影响细胞分裂和细胞伸长促进侧根和根毛产生及根毛分叉,其他激素如ETH通过调控JA达到抑制根伸长的目的[9-13]。氮素能引起内源糖的变化[14],控制拟南芥根系的伸长生长,而糖对植物根系生长的影响也是通过与植物激素互作产生的,如葡萄糖或蔗糖通过影响 IAA合成运输的途径影响植物根系伸长生长[15-16]。氮素也能引起内源 NO的形成,在根的生长发育中,硝酸还原酶产生的 NO依赖于硝酸盐的浓度[17],而外源高剂量的NO会使植物初生根长度缩短,促进侧根形成。研究表明,NO与H2O2共同参与IAA介导的万寿菊外植体不定根发育,而过氧化氢调节植物生长并影响各种发育过程,包括根生长以及根的形成和发育[18-21]。生根相关酶中,PPO诱导植物生根,并催化IAA酚酸化合物,IAAO与PPO通过改变IAA水平调控根系生长,POD通过活性变化调控根原基形成过程中IAA的代谢[22-24]。【本研究切入点】大量研究表明铵态氮和硝态氮能够对植物根系形态进行调控,且两种氮素对植物根系的形态调控作用不同[25-26],但多数研究并未明确阐述不同浓度下单一氮素形态如何调控根系形态。【拟解决的关键问题】本研究以娃娃菜为试材,采用低、高浓度的铵态氮和硝态氮营养液对娃娃菜进行水培,观察和测定单一氮素形态及水平对娃娃菜幼苗根系形态的影响,并从生理代谢水平研究两种氮素形态对娃娃菜根系形态的调控机理,为农业生产中娃娃菜根系形态的调控提供理论基础。
1 材料与方法
试验于2018年6月至2019年4月在甘肃农业大学进行。
1.1 供试材料
供试材料为娃娃菜(Brassica pekinensis,‘惠农 金娃娃’),种子购于甘肃省农业科学院种子市场。
1.2 试验方法与设计
选取颗粒均匀、饱满的种子,温汤浸种后置于垫有润湿滤纸的培养皿里,放在25℃黑暗条件下催芽20 h。待子叶长出后,生长条件设置如下:光周期12 h/12 h,温度25℃/18℃(昼夜),湿度75%,光照143 μmol·m-2·s-1,蒸馏水培养至根长达到 4 cm 左右移至自制培养装置(高45 mm、口径120 mm、底径85 mm)中处理,每盆9株,每个处理4盆。4种处理液设置如下:0.1 mmol·L-1NH4+(LA)、1.0 mmol·L-1NH4+(HA)、0.1 mmol·L-1NO3-(LN)和1.0 mmol·L-1NO3-(HN)。营养液分别用KNO3和NH4Cl配制,加入KCl使其溶液浓度与营养液浓度相同,以平衡离子。2 d更换一次营养液,处理8 d后采样进行各项指标测定。
1.3 测定指标及方法
1.3.1 根系形态参数的测定 娃娃菜活体根系从根原基处剪断,根系扫描仪(型号 Epson Expression 11000XL)扫描根系图片,WinRHIZO软件分析得出总根尖数、总根长、总根体积和总根面积。
1.3.2 生理指标的测定 铵态氮测定参照张婧的方法[27];硝态氮的测定采用硝基水杨酸比色法;还原糖测定用3,5-二硝基水杨酸比色法;可溶性糖测定用蒽酮比色法;NO测定用Griess试剂法;H2O2测定用张小莉等[28]的方法;POD活性采用愈创木酚比色法测定;PPO活性检测用邻苯二酚比色法;IAAO活性参考高俊凤[29]的方法。
1.3.3 根系中内源激素含量的测定 植物根系中的细胞分裂素类(CTK)、茉莉酸类(JA)、生长素(IAA)、乙烯前体1-氨基环丙烷羧酸(ACC)、脱落酸(ABA)、水杨酸(SA)采用同位素内标法,对含量进行LC-MS/SM绝对定量分析。样本制备方法、激素标准样品以及LC-MS/MS定量分析方法均由上海中科新生命生物科技有限公司(www.aptbiotech.com)提供。
样本采集以及制备:取新鲜植物根系采用冷生理盐水洗涤去除样品表面泥土或污染物,液氮中淬灭,-80℃保存,低温下(干冰中)运输至技术中心。每个组织样本全部液氮研磨,混匀后称取所需样品,加入同位素内标,800 μL甲醇/水(4∶1,v/v)溶液,漩涡混合30 s,低温超声萃取20 min,-20℃孵育1 h,13 000 r/min,4℃离心20 min,取上清液,固相萃取,萃取液氮气吹干,-80℃低温保存待用。
LC-MS/MS绝对定量分析:采用UPLC反相色谱系统对样本进行分离,AB 5500Q-TRAP质谱采集MRM模式数据一套。
1.4 数据统计分析
采用EXCEL 365和SPSS 26.0软件对植物生理指标、根系内源激素与根系形态参数进行数据统计分析并做相关性分析,数据用平均值±标准误(n=3)表示,使用TBtools作相关性分析图。
2 结果
2.1 氮素形态及浓度对娃娃菜幼苗根系形态的影响
由表1可知,相同氮素形态下,LN较HN分别显著提高总根长、总根表面积、总根尖数43%、24%和50%。LN可增加根系体积,但相比HN差异不显著。与HA相比,LA显著增加总根长、总根体积、总根表面积和总根尖数(P<0.05),分别提高了96%、73%、85%和45%。在同一氮素浓度下,HN与HA相比,能够显著增加总根长、总根表面积和总根体积(P<0.05);而 HA能够增加总根尖数,但差异不显著。LN与LA相比,能够增加总根长、总根表面积;LA能够增加总根体积和总根尖数,但差异均不显著。
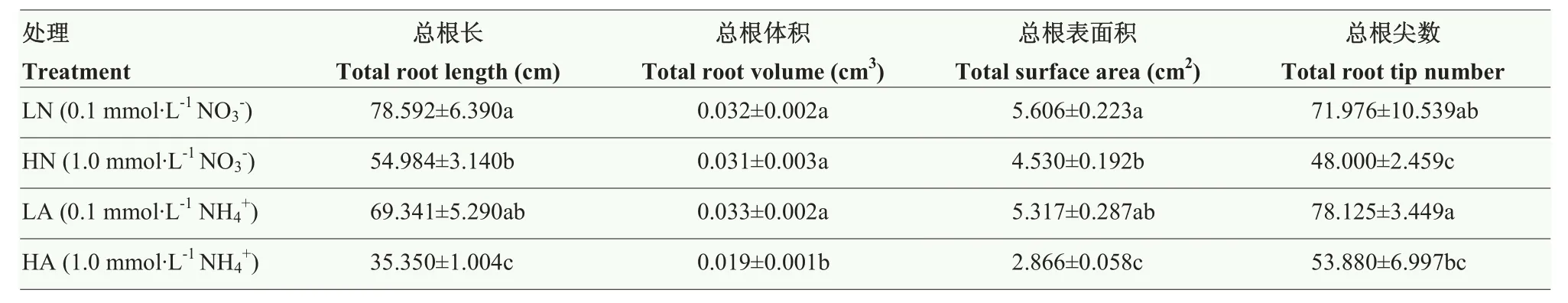
表1 氮素形态及浓度对娃娃菜幼苗根系形态的影响Table 1 Effects of nitrogen form and concentration on root morphology in mini Chinese cabbage seedlings
从图1可直观地看到4种处理下娃娃菜的根系形态,其中主根长表现为LN>HN>LA>HA,一级侧根长度也出现了相同的变化,说明硝态氮对主根和侧根长度具有显著的促进作用,低浓度影响更加显著,而低浓度铵态氮促进根长作用与低浓度硝态氮相比不明显。

图1 不同处理下娃娃菜幼苗的根系形态Fig.1 Root morphology of mini Chinese cabbage seedlings under different treatments
2.2 氮素形态及浓度对娃娃菜幼苗根系铵态氮、硝态氮、还原糖及可溶性糖的影响
从表2可以看出,8 d后不同处理根系组织中硝态氮、铵态氮、还原糖以及可溶性糖含量差异明显。LN处理下硝态氮和还原糖含量均高于其他处理,与 HN处理相比,根系硝态氮含量提高14.99%,但差异不显著。还原糖含量提高55.81%,且在硝态氮的处理下, 娃娃菜根系硝态氮与还原糖水平整体高于铵态氮的处理,如LA处理硝态氮含量低于LN处理67.10%,差异显著(P<0.05);LA处理相比 HA处理显著提高娃娃菜根系还原糖含量 200%(P<0.05)。铵态氮含量在LN处理与LA处理中相近,在HN处理与HA处理中也相近,高浓度氮素处理下娃娃菜根系铵态氮含量显著高于低浓度处理,4种处理下还原糖含量变化与其类似。以上结果说明氮素营养对娃娃菜根系氮、糖含量影响是从其自身浓度高低来产生作用,低浓度氮素可能更利于娃娃菜根系糖类物质的积累。
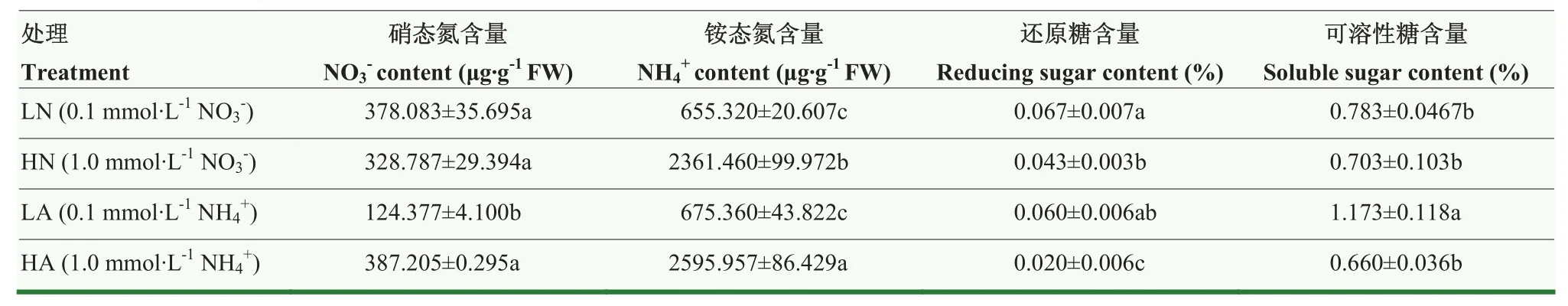
表2 氮素形态及浓度对娃娃菜幼苗根系铵态氮、硝态氮、还原糖和可溶性糖含量的影响Table 2 Effects of nitrogen form and concentration on the content of ammonium nitrogen,nitrate nitrogen,reducing sugar and soluble sugar in root of mini Chinese cabbage seedlings
2.3 氮素形态及浓度对娃娃菜幼苗根系 NO含量及H2O2含量的影响
硝态氮处理下的娃娃菜根系NO和H2O2含量均显著高于铵态氮处理(表3),氮素形态相同时,LN相比HN提高NO含量18.31%, H2O2降低了20.44%。LA相比HA提高NO和H2O2含量74.59%和13.58%,但差异均不显著。氮素浓度相同时,LA处理NO含量比LN处理低73.68%,H2O2含量低40.98%,两者差异显著(P<0.05)。HA处理NO和H2O2比HN处理分别低82.16%和58.66%,差异均显著(P<0.05)。由此可见,硝态氮处理娃娃菜会使其根系NO和H2O2的含量增多。
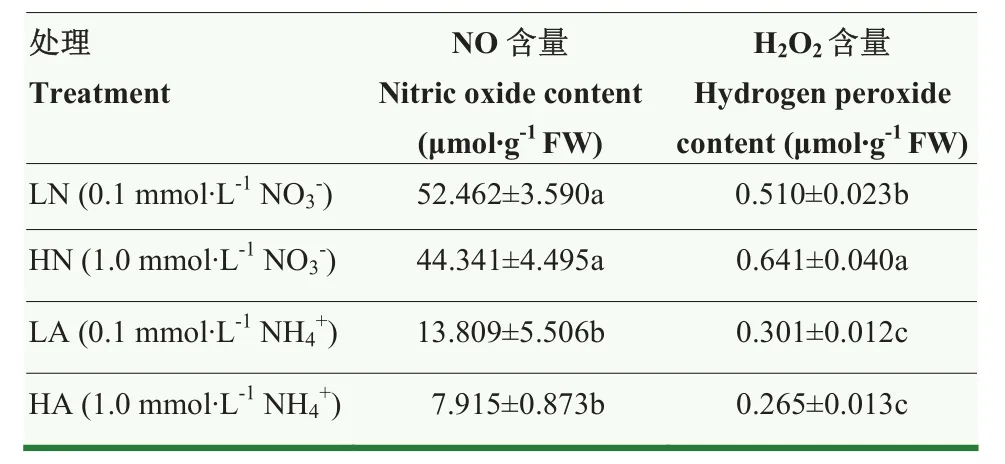
表3 氮素形态及浓度对娃娃菜幼苗根系NO含量及H2O2含量的影响Table 3 Effects of nitrogen form and concentration on NO and H2O2content in root of mini Chinese cabbage seedlings
2.4 氮素形态及浓度对娃娃菜幼苗根系生根相关酶活性的影响
氮素形态和浓度显著影响娃娃菜根系生根相关酶的活性,由表4可以看出,硝态氮处理下的POD活性和IAAO活性低于铵态氮处理下的酶活性。其中,LN处理下POD酶活性低于HN处理55.04%,HA处理下POD酶活性最高。PPO酶活性在LA处理下最低,低于HA处理56.41%,在HN处理时最高,高于LN处理 50.93%,差异均显著(P<0.05),硝态氮处理下的PPO活性高于铵态氮处理。IAAO酶活性在HA处理高于LA处理34.98%,LN处理最低,低于HN处理 16.39%,差异均显著(P<0.05)。说明铵态氮对POD和IAAO酶活性有显著促进作用,而硝态氮更加有利于PPO酶活性。
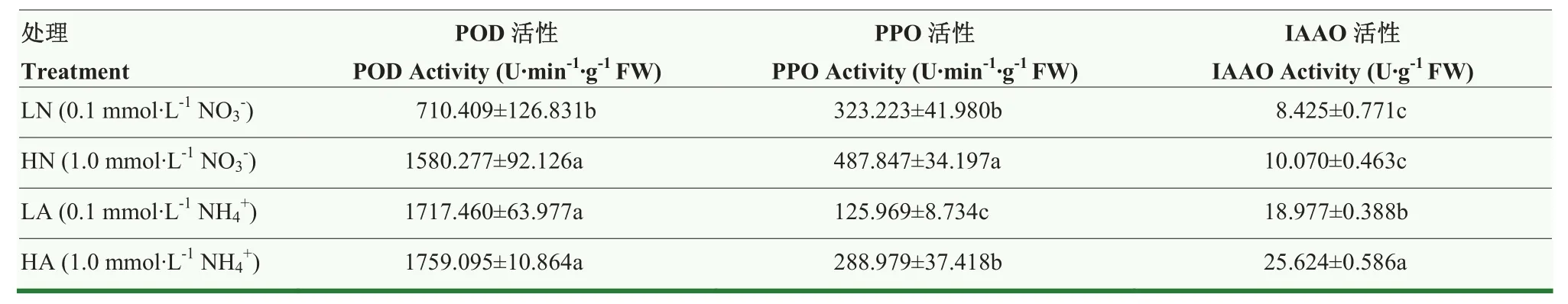
表4 氮素形态及浓度对娃娃菜幼苗根系生根相关酶活性的影响Table 4 Effects of nitrogen form and concentration on the activities of root-related enzymes in root of mini Chinese cabbage seedlings
2.5 氮素形态及浓度对娃娃菜幼苗根系激素含量的影响
4种处理下IPR、tZR、tZ含量出现了显著差异,且铵态氮处理下的整体细胞分裂素含量高于硝态氮处理,低浓度氮素效应更好。LA处理IPR、tZR、tZ含量分别高于HA处理78.03%、37.25%和40.44%。虽然cZR、IP和cZ含量数值略有不同,但均未有显著差异(表5)。
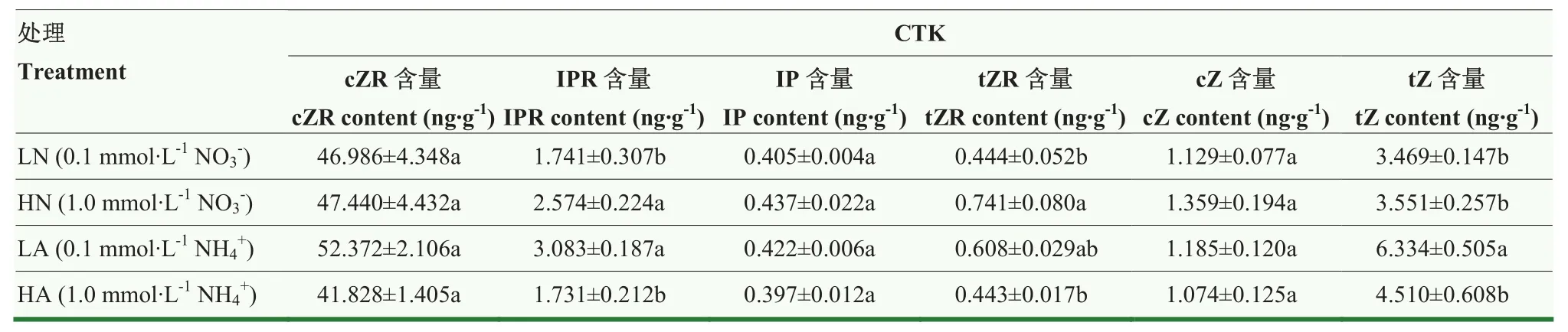
表5 氮素形态及浓度对娃娃菜幼苗根系细胞分裂素含量的影响Table 5 Effects of nitrogen form and concentration on the content of cytokinin in root of mini Chinese cabbage seedlings
LA处理和HA处理的娃娃菜根系茉莉酸类激素含量显著高于LN处理和HN处理,12-OPDA含量以LA处理最大,高于HA处理55.18%,差异显著(P<0.05)。LA与HA相比,JA与JA-ILE均无显著差异,但在铵态氮处理下,低浓度铵态氮对茉莉酸类激素促进效果更佳。IAA、SA、ACC和ABA含量呈现在HN处理和LA处理中较高,且除ABA外,其他3种激素在HN处理与LA处理中的含量显著高于另外两个处理(P<0.05)。IAA含量在HN处理中最大,LA高于HA处理93.79%。HA处理的SA含量低于LA处理54.09%,HN处理高于LN处理26.89%,差异显著(P<0.05)。氮素形态一致时,乙烯前体ACC含量表现为HN比LN处理高51.58%。浓度相同时,HN高于HA处理37.75%,LA高于LN处理22.54%,差异显著(P<0.05)。4种处理ABA含量均无显著差异(表6)。

表6 氮素形态及浓度对娃娃菜幼苗根系JA类激素、IAA、SA、ACC和ABA含量的影响Table 6 Effects of nitrogen form and concentration on JA,IAA,SA,ACC and ABA content in root of mini Chinese cabbage seedlings
2.6 娃娃菜根系各物质与根系形态参数的相关性分析
根据相关性分析结果(图1),总根表面积与总根长、总根体积呈显著正相关(P<0.05)。总根长与还原糖呈极显著正相关(P<0.01),说明在娃娃菜根系生长发育中,还原糖在娃娃菜根系伸长生长的过程中起到了重要作用。12-OPDA为茉莉酸前体,结果显示根尖数与 12-OPDA呈显著正相关(P<0.05),表明在娃娃菜根系侧根生长发育过程中茉莉酸作用显著,根尖数与可溶性糖也呈显著正相关(P<0.05)。其他生理指标与根系形态的相关性如下图所示。
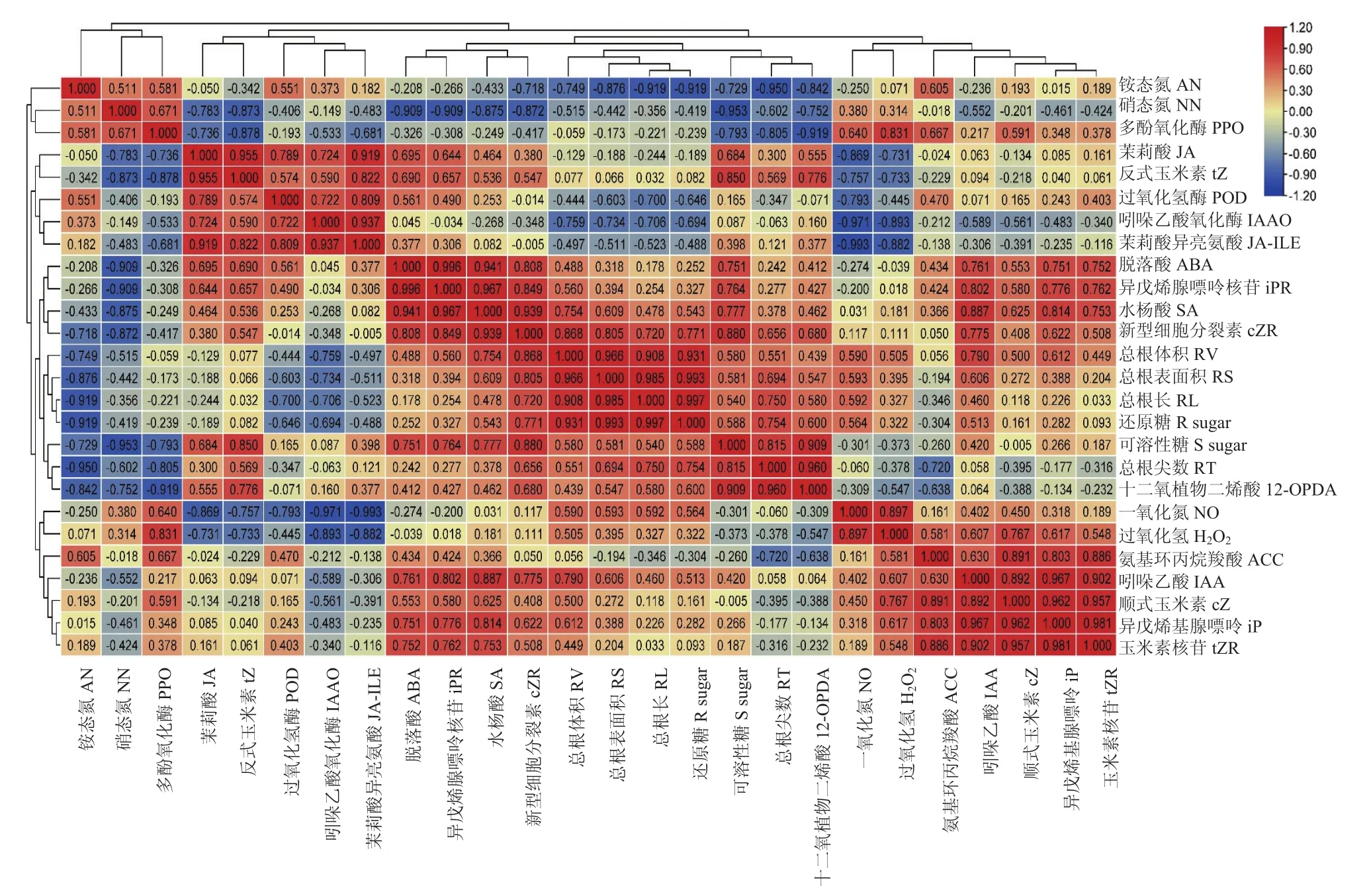
图2 根系生理指标与根系形态参数的相关性关系Fig.2 Correlation between root physiological indexes and root morphological parameters
3 讨论
3.1 植物根系氮吸收及影响氮吸收的因素
根系吸收的主要氮素形态是铵态氮与硝态氮,根系形态的建成与氮素形态相关[30],铵态氮可以直接被植物根系利用而硝态氮多数被运输到茎叶中才被利用[31]。硝态氮在植物根系建成过程中,可以有效促进植物根系的伸长生长,作用显著大于铵态氮,但铵态氮处理下植物含氮量高于硝态氮处理[26],拟南芥中铵态氮供应能引起高度分枝短根的出现[32],也能刺激根系生长素积累[6],这与本研究中娃娃菜根系在不同氮素形态下生长发育的结果相一致。大量研究已表明,植物中的 NO、H2O2、糖、激素等都是信号物质以调节植物体内生理稳态。以植物基本营养之一的氮素为研究目标,在不同形态和水平氮素条件下对这些物质含量变化进行测定发现,单一氮素形态和水平并不是单独在植物根系的生长发育中产生作用,而是与多种信号物质共同作用的结果,其中,糖、NO、H2O2和激素等都参与植物根系形态构建[33-35]。
3.2 植物中糖类物质参与氮素对娃娃菜根系的调控
植物根系吸收水分和营养物质后,通过光合作用生成糖,在植物的生长发育中,糖既能促进细胞的碳和能量代谢,还能像激素一样作为信号物质参与并调控植物开花、衰老以及根系的生长发育,以及控制与植物生长和代谢有关的不同基因的表达,但需要与其他必须营养相结合,如氮、磷等[36-37]。氮素形态影响植物体内糖的变化并参与植物根系形态的建成是植物调控根系形态的一种方式,植物幼苗叶片感受光刺激后子叶光合作用产生的糖经韧皮部运输到根尖,通过抑制根尖分生区的长度调节主根生长[38-39],且糖在调节植物根系生长过程中是通过影响IAA的运输来产生作用的。因此,氮素影响植物根系生长发育过程中,首先是根系吸收氮素运输到地上部分,并引起植物对糖类物质的合成,然后糖又从地上部运输到根系引起IAA运输并以此来调控根系生长。本研究中,低氮条件下的还原糖含量高于高氮下的含量,而且无论何种氮素形态,低氮条件下娃娃菜根系总根长均大于高氮条件下的总根长,说明不同氮素浓度也参与植物根长的调节过程。
3.3 植物激素及NO等参与娃娃菜根系形态的调控
众多激素中,生长素在根系生长发育中起着非常重要的作用,其生物合成、运输和信号传递都会影响侧根的形成与主根的伸长,可以影响植物根系生长方向和形态建成[40-41]。生长素介导的侧根生长受氮素影响,局部铵供应会刺激根系维管系统中由地上部运输而来的生长素积累,以促进侧根的出现,并建立高度分枝的根系,而局部的硝供应可以显著促进根的伸长[6]。黄秀等[42]的研究表明,在 0—0.8 mmol·L-1的低浓度铵态氮下,水稻根系对铵态氮有高亲和吸收效应,而在铵态氮浓度大于 1.0 mmol·L-1后,水稻根系生长受到限制。杨娜[43]也在拟南芥的研究中发现,低浓度的铵态氮与低浓度的生长素共同作用可以协同促进侧根发生,0.1 mmol·L-1的铵态氮可以增加侧根密度,而0—1 mmol·L-1硝态氮显著促进侧根伸长生长。本研究中硝态氮促进根系伸长生长作用显著,尤其是在低浓度处理时,数据结果显示低浓度铵态氮也能够明显促进娃娃菜总根长,而且在不同处理下内源生长素含量出现了差异,表明氮素浓度和含量影响生长素调控娃娃菜根系形态,但生长素浓度是否也响应了不同的调控作用,具体响应机制有待进一步探究。茉莉酸类物质能够促进侧根生长,提高根毛密度和长度,并且是与铵态氮协同工作以增强侧根的发生[44-45],笔者在试验中发现随着铵态氮浓度升高,娃娃菜根系茉莉酸减少,而高浓度硝态氮处理中茉莉酸含量高于低浓度处理中的含量,且12-OPDA与根尖数正相关,说明氮素营养参与娃娃菜内源茉莉酸对其根系形态的调控,正如氮素营养响应生长素调控娃娃菜根系一样。植物激素并不能单一的产生作用,它们形成了一个复杂的网络调节植物的生长发育过程,包括对根系形态的调控。生长素能调节乙烯的合成,有研究表明生长素促进根毛生长是通过内源乙烯途径,而内源乙烯的拮抗作用又抑制了铵态氮对侧根生长的刺激[44,46]。细胞分裂素类激素通过负向调控与生长素共同调控植物根系形态结构[47-48],其中,CRZ对主根有促进作用[49],水杨酸作为植物防御激素可以通过阻断IAA信号系统抑制根内细菌侵蚀,从而抑制侧根形成[5],植物体内脱落酸可以调节主根生长,也可以抑制侧根生长[18,50],试验中 4个处理下脱落含量变化不大,在娃娃菜生长发育前期调控作用可能不明显。激素等信号物质可以对植物根系形态进行协同调控,但是他们的作用取决于氮素形态和水平,并且协同不同氮素形态和水平共同产生作用,氮素营养响应激素调控植物根系生长发育的机理仍有很大的研究空间。IAAO通过氧化生长素使其在植物体内保持稳定的水平,生长素水平与IAAO、POD和PPO都有关[51-52],但其活性不能作为判断生根的必要条件,生根与生长素相对含量有关,生长素受到限制时,侧根受到硝态氮的刺激而降低,说明硝态氮参与植物生根调节[53]。本试验中,铵态氮处理下的娃娃菜根内IAAO的活性明显更高,生长素含量较低,促进生根。在生根过程中PPO活性也会升高,促使酚类物质再与生长素结合生成生根复合物,加速生根[54]。硝态氮与铵态氮处理下的植物POD活性相差不大,均有利于植物POD活性的增加,不会随着浓度的增加而变化,试验中IAAO变化趋势与POD一致,与已有研究相符[49]。
NO可以诱导植物根系伸长[10],其产生与硝态氮处理有关,参与硝酸盐调控植物根系生长[53]。NO可能调控生长素来调控乙烯前体 ACC合成乙烯以控制植物根系生长,其作为信号物质可以诱导水杨酸的合成,而水杨酸可以激活植物NO合成途径[12,55]。H2O2对植物而言也是一种信号物质,参与植物的各种生物和非生物性压力的反应,少量H2O2参与野生型拟南芥根毛的形成,也与侧根的生成有关[18,56]。
本研究中,由于4种处理下铵态氮和硝态氮变化规律不像其他指标一样,导致相关性分析结果出现偏差,但根据植物吸收铵态氮与硝态氮的规律[57-58],4种处理下硝态氮与铵态氮含量变化如下所述:植物根系吸收硝态氮后,大部分硝态氮在根际硝酸还原酶(NR)作用下还原为亚硝酸盐,亚硝酸盐又在亚硝酸还原酶(NiR)的作用下还原为铵态氮,使植物根系中铵态氮含量上升,所以 LN处理下还原量较少,HN处理下还原量较多,出现了本研究的结果。同理,根系吸收铵态氮后,大部分可以直接被同化产生氨基酸,而转化为硝态氮的只有很少一部分,因此,LA处理下硝态氮含量极低,铵态氮含量较高,HA处理下硝态氮含量较LA处理下高,铵态氮含量极高。
4 结论
氮素营养不仅从形态上影响娃娃菜根系的生长,而且浓度不同产生的作用也有差异。两种形态氮素调控娃娃菜根系形态的过程中,低浓度氮素作用更加显著;还原糖与根长最具相关性,茉莉酸前体 12-氧植物二烯酸(12-OPDA)与根尖数相关性最强。低浓度硝态氮(LN)影响娃娃菜根系中还原糖的含量以增加娃娃菜根系长度;而低浓度铵态氮(LA)影响 12-OPDA,从而增加娃娃菜侧根数量。
猜你喜欢
农业现代化研究(2022年3期)2022-06-29 09:31:54
生物学通报(2021年4期)2021-03-16 05:41:34
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29 03:15:14
农业环境科学学报(2020年10期)2020-11-11 02:55:28
科学大众(2020年12期)2020-08-13 03:22:38
乡村科技(2019年9期)2019-06-26 03:52:04
数码世界(2018年7期)2018-12-24 15:06:05
湖泊科学(2015年1期)2015-05-06 07:14:56
植物营养与肥料学报(2011年5期)2011-11-06 07:30:02
植物营养与肥料学报(2011年4期)2011-10-26 02:44:56
