
冬季限食对树麻雀能量代谢的影响及其生理适应
2022-02-24 06:09刘佳人徐兴军王有祥林美含邵淑丽
野生动物学报 2022年1期
刘佳人 徐兴军* 王有祥 林美含 邵淑丽
(1.齐齐哈尔大学生命科学与农林学院,齐齐哈尔,161006;2.抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,齐齐哈尔,161006;3.乳山畅园学校,乳山,264599)
温度是影响动物产热的重要因素[1]。在北方,一年四季中,冬季的环境条件最为苛刻,气候寒冷,食物短缺,对冬季环境条件的适应是鸟类生存的重要前提[2]。鸟类消耗能量的速率可能对其健康产生深远地影响,从而影响其生理、行为、生态和进化[3]。通过了解鸟类的代谢规律,能进一步理解鸟类在应对环境季节性变化时的应对策略[4]。本研究以树麻雀(Passermontanus)为试验材料,探讨冬季树麻雀在限食条件下的能量预算对策,为进一步了解北方小型鸟类的生存机制提供理论依据。
1 材料与方法
1.1 材料
树麻雀于2019年12月捕自黑龙江省齐齐哈尔市扎龙国家级自然保护区(46°52′—47°32′N,123°47′—124°37′E)。
1.2 方法
树麻雀在齐齐哈尔大学动物学实验室适应性饲养1周后,按体重随机分为对照组、半饥饿驯化组、1/4饥饿驯化组,组间体重无明显差异(P>0.05)。各驯化组均饲喂谷子,以每天每只鸟的正常食量(20.15±1.08)g为标准,半饥饿驯化组投喂标准食量的1/2,1/4饥饿驯化组投喂标准食量的1/4。每组8只,自由饮水,自然光照,驯化4周。
体重和各器官重量测定:采用BS124S型电子天平测量,精确到0.01 g。驯化开始前,分别测定各组树麻雀的初始体重。驯化结束后,测定各组树麻雀终体重及主要器官重。
基础代谢率(basal metabolic rate,BMR)测定:利用动物气体代谢分析仪测量,每隔7 d测定1次,测量前先将树麻雀禁食4 h,单只称量树麻雀的体重,精确至0.01 g。测定开始前2 h,将氧气和二氧化碳分析仪打开预热,然后对其进行标定,硬件与软件标定完成后分别将树麻雀放入已编号的4个呼吸室内适应1 h,待树麻雀呼吸稳定后开始测量,记录每分钟的耗氧量,共测定30 min,分别取5个连续稳定的数值进行计算。
体脂质量分数测定:采用索氏抽提法[5]测定。体脂质量分数=(胴体干重-抽提后不含体脂的胴体干重)/个体体重×100%。
个体含水量测定:采用质量差法测定。个体含水量=(胴体的鲜重-胴体的干重)/胴体的干重×100%。
摄入热量值及消化率的测定:采取全收粪法测定[6]。
糖原质量分数的测定:采用硫酸蒽酮测定法测定肌糖原、肝糖原质量分数[7]。
淀粉酶活性的测定:清除组织内的残留物,用滤纸吸干并称重,置于研钵内加入10倍组织量的0.02 mol/L、pH=7.5的磷酸缓冲溶液,在冰盒内充分研磨成糊状匀浆,将匀浆液转入离心管置于冷冻离心机,4 ℃、12 000 r/min离心15 min,上清液即为粗酶液。取50 mL的比色管,用记号笔标明对照管与待测管,在2管内分别加入0.04%可溶性淀粉5.0 mL,37 ℃水浴5 min;对照管加0.3 mL蒸馏水,测定管加0.3 mL粗酶液,混匀、37 ℃水浴10 min;再在2管内分别加入0.01 mol/L碘应用液5.0 mL,用蒸馏水稀释至50.0 mL,立即混匀。于660 nm波长处比色,蒸馏水调零,读取对照管和测定管光密度。淀粉酶活力=[(OD对照-OD待测)/OD对照×200]/0.3 mL粗酶液中蛋白克数。
肠道组织的测定:采用石蜡切片法对树麻雀肠道组织进行制片[8]。应用Nikon显微成像系统在10×40倍视野对肠道组织的绒毛高度、绒毛宽度、黏膜层厚度、肠壁截面积进行测量。
1.3 数据分析
利用SPSS 17.0软件对数据进行单因素方差分析。
2 结果与分析
2.1 限食对树麻雀的体重和器官重的影响
由图1可见,驯化开始时,限食组树麻雀的体重组间无明显差异,1周后半饥饿驯化组与1/4饥饿驯化组体重较对照组极显著降低(P<0.01)。随着驯化时间的延长,半饥饿驯化组与1/4饥饿驯化组的体重在1~3周内大幅度降低,在第4周驯化结束时达到最低值,与初始体重相比分别减少1.06、3.81 g。第4周驯化结束后,3组树麻雀体重组间差异均极显著(P<0.01)。
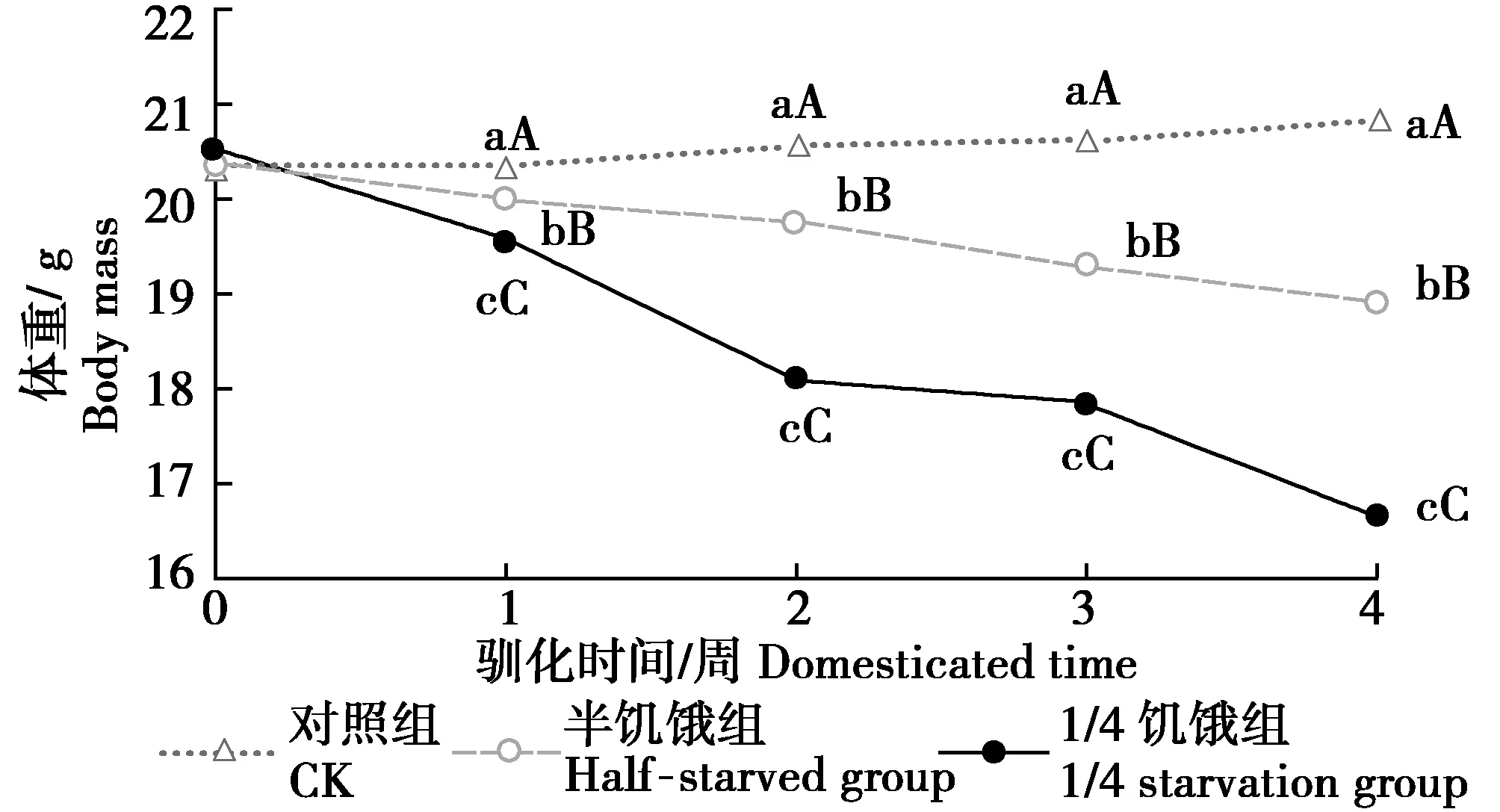
图1 限食对冬季树麻雀体重的影响
半饥饿驯化组、1/4饥饿驯化组树麻雀的心脏、肝脏、肺、胰腺和十二指肠鲜重均比对照组低;大肠、小肠、直肠、肌胃和腺胃鲜重均比对照组高。3组树麻雀的心脏、肝脏、肺、胰腺、大肠、小肠、直肠、十二指肠、肌胃和腺胃鲜重组间差异均极显著(P<0.01)(表1)。
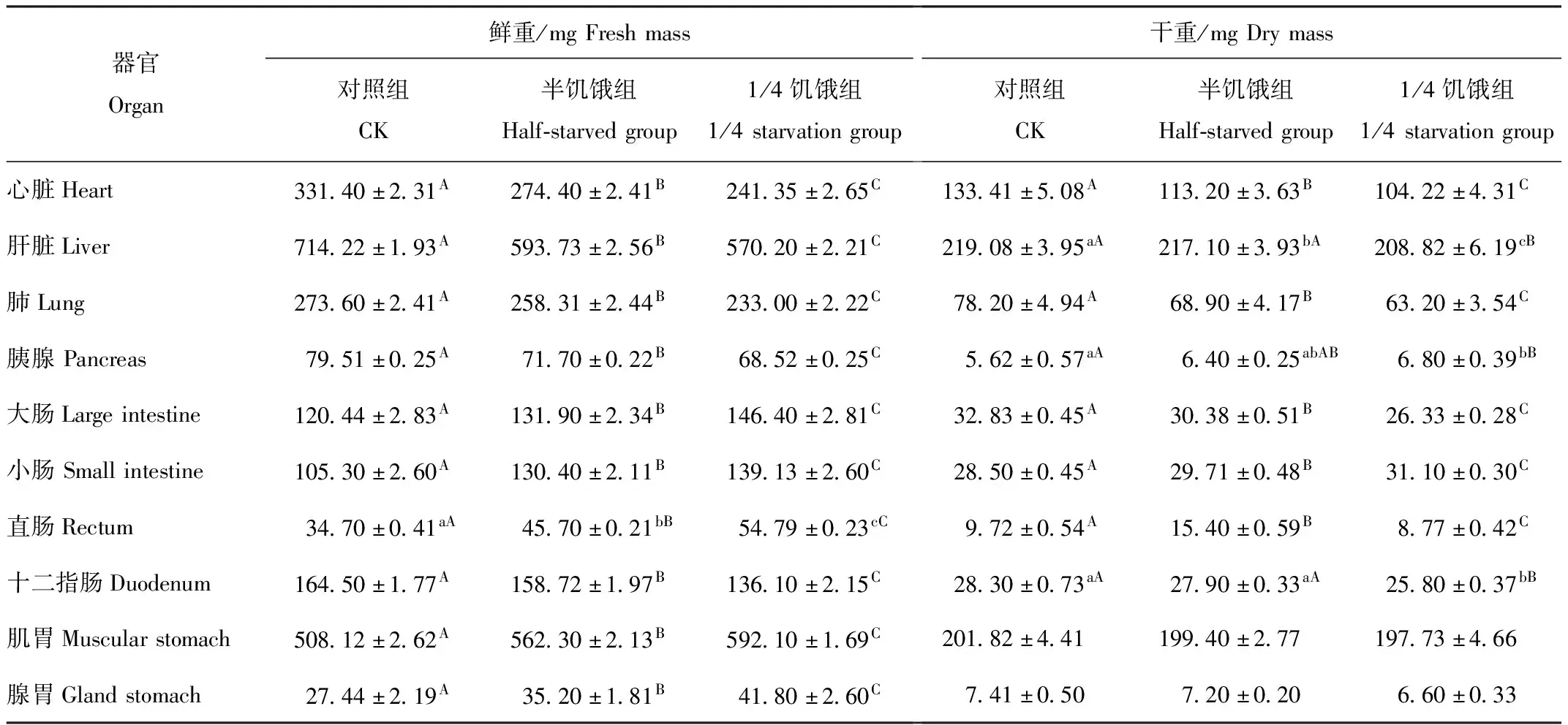
表1 限食对冬季树麻雀器官重量的影响
由表1可见,半饥饿驯化组、1/4饥饿驯化组树麻雀的心脏、肝脏、肺、大肠、十二指肠、肌胃和腺胃干重均比对照组低;胰腺和小肠干重均比对照组高。半饥饿驯化组直肠干重比对照组高58.44%,1/4饥饿驯化组直肠干重比对照组低9.77%。半饥饿驯化组、1/4饥饿驯化组心脏、肝脏、肺、大肠、小肠、直肠、十二指肠干重与对照组相比组间差异均极显著(P<0.01)。对照组与半饥饿驯化组胰腺干重组间差异不显著(P>0.05),与1/4饥饿驯化组相比组间差异极显著(P<0.01)。
2.2 限食对树麻雀BMR的影响
由图2可见,随着驯化时间的延长,半饥饿驯化组与1/4饥饿驯化组BMR在0~1周内大幅度降低,在第4周驯化结束时达到最低值,与初始BMR相比分别降低0.98、1.08 mL/(g·h)。第4周驯化结束后,3组树麻雀BMR组间差异均极显著(P<0.01)。
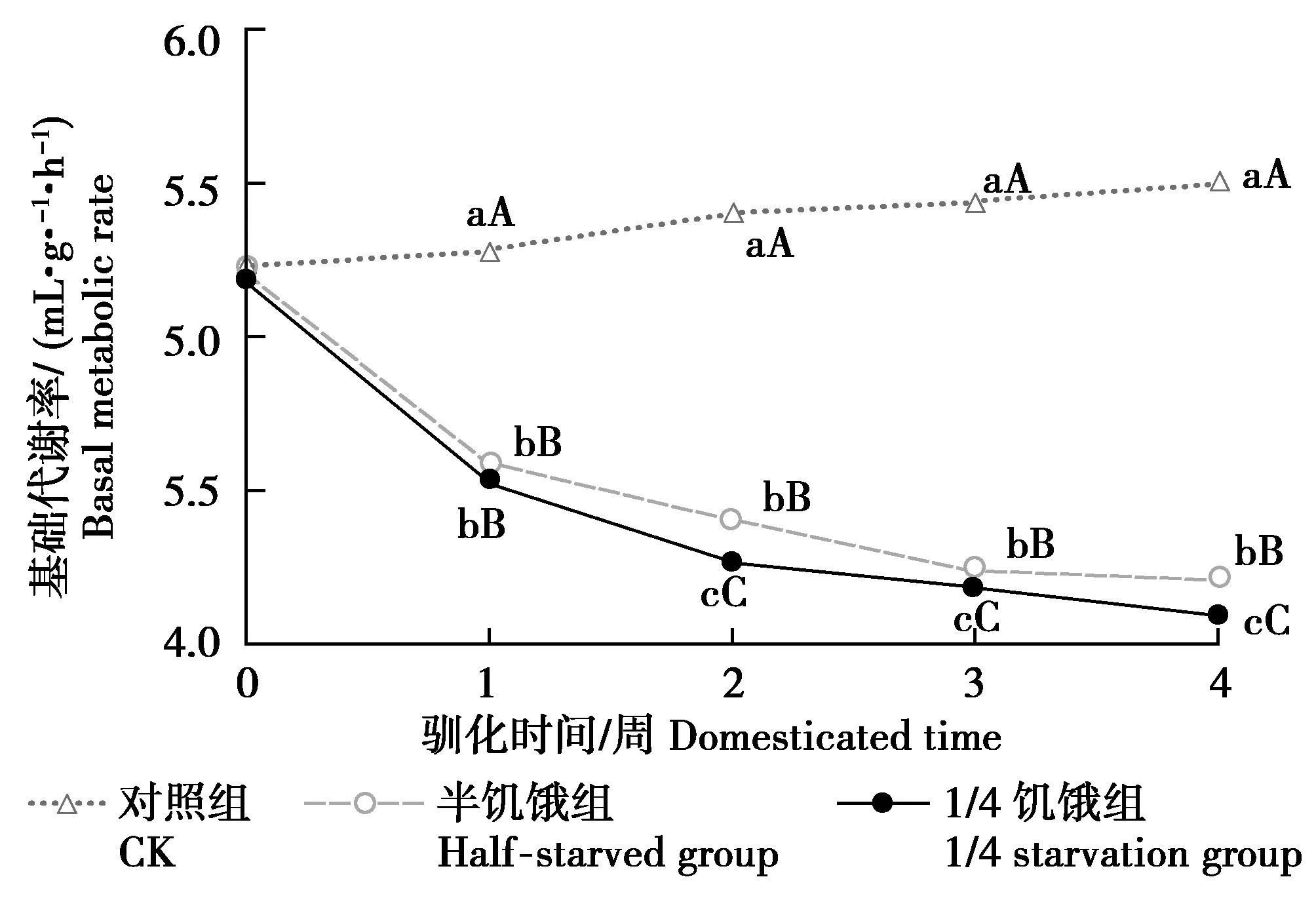
图2 限食对冬季树麻雀基础代谢率的影响
2.3 限食对树麻雀生理指标的影响
由表2可见,1/4饥饿驯化组树麻雀的含水量分别比半饥饿驯化组、对照组高2.11%、2.87%,且差异极显著(P<0.01)。饥饿驯化组的消化能和消化率均极显著高于对照组(P<0.01)。半饥饿驯化组、1/4饥饿驯化组的体脂质量分数均比对照组低,但只有1/4饥饿驯化组与对照差异极显著(P<0.01)。
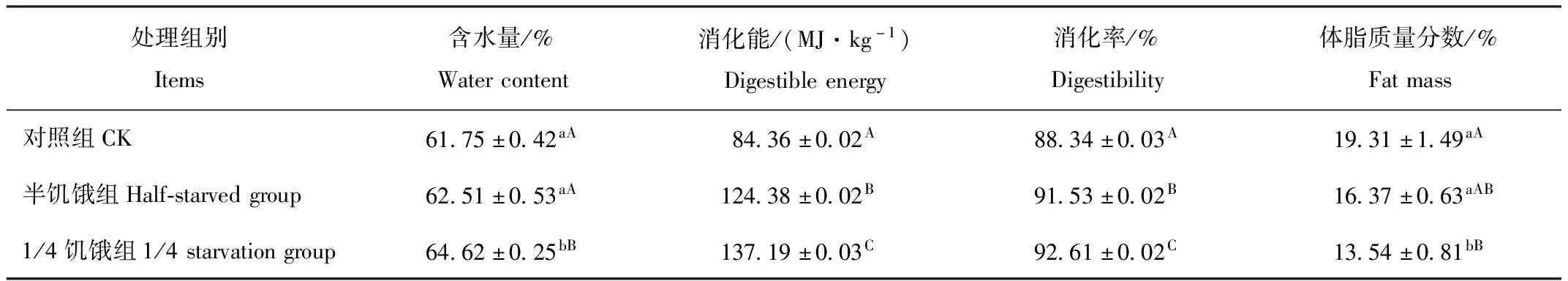
表2 限食驯化4周后树麻雀体内各生理指标
2.4 限食对树麻雀消化道长度的影响
由图3可见,限食驯化后,树麻雀的大肠、小肠、直肠长度由长到短依次为:1/4饥饿驯化组、半饥饿驯化组、对照组,且限食驯化组均极显著长于对照组(P<0.01),但限食驯化对十二指肠的长度没有明显影响(P>0.05)。

图3 限食对冬季树麻雀消化道长度的影响
2.5 限食对树麻雀淀粉酶活性的影响
由表3可见,1/4饥饿驯化组、半饥饿驯化组树麻雀肝脏中的淀粉酶活性较对照组高,但只有1/4饥饿驯化组与对照组差异显著(P<0.05)。1/4饥饿驯化组、半饥饿驯化组胰腺、大肠、小肠、直肠、十二指肠、肌胃和腺胃中的淀粉酶活性较对照组低,其中半饥饿组胰腺、大肠中的淀粉酶活性与对照组差异极显著(P<0.01),1/4饥饿驯化组胰腺、大肠、小肠、肌胃和腺胃与对照组差异极显著(P<0.01)。
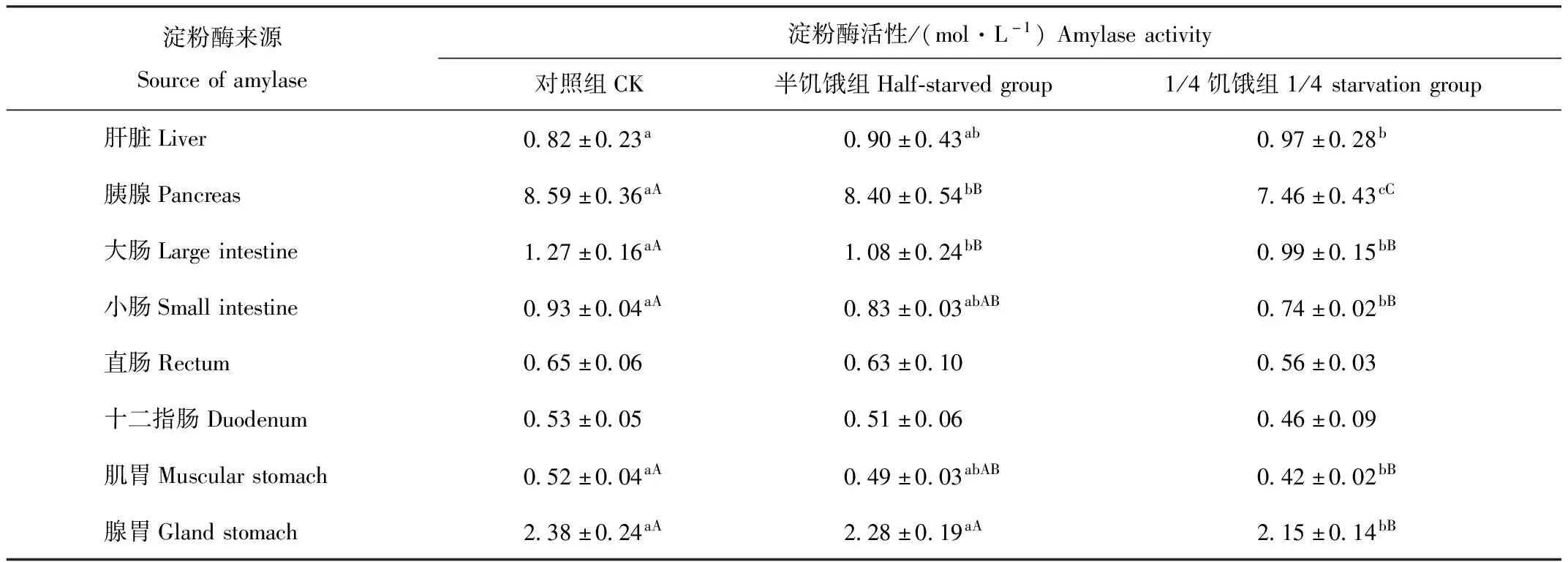
表3 限食对冬季树麻雀淀粉酶活性的影响
2.6 限食对树麻雀糖原质量分数的影响
由表4可见,冬季对照组树麻雀肝糖原质量分数比半饥饿驯化组、1/4饥饿驯化组分别高0.697%、0.754%。对照组肌糖原质量分数比半饥饿驯化组、1/4饥饿驯化组分别高0.244%、0.391%。3组树麻雀肝糖原、肌糖原质量分数组间差异均极显著(P<0.01)。
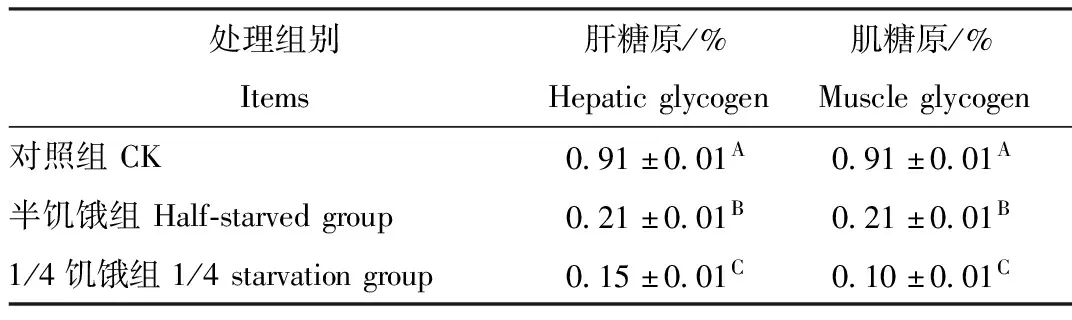
表4 限食对冬季树麻雀糖原质量分数的影响
2.7 限食对树麻雀消化器官各形态指标的影响
随着限食程度的加大,半饥饿驯化组和1/4饥饿驯化组的消化道绒毛高度、宽度与对照组相比呈降低趋势,消化道黏膜厚度呈上升趋势(图4)。由表5可见,3组树麻雀的大肠绒毛高度、直肠绒毛高度和十二指肠壁截面积差异显著(P<0.05),1/4饥饿驯化组与对照组差异极显著(P<0.01)。半饥饿驯化组小肠壁截面积、十二指肠绒毛高度和小肠绒毛高度与对照组差异不显著,但1/4饥饿驯化组却极显著低于对照组(P<0.05),半饥饿驯化组和1/4饥饿驯化组十二指肠绒毛宽度均显著低于对照组(P<0.05)。
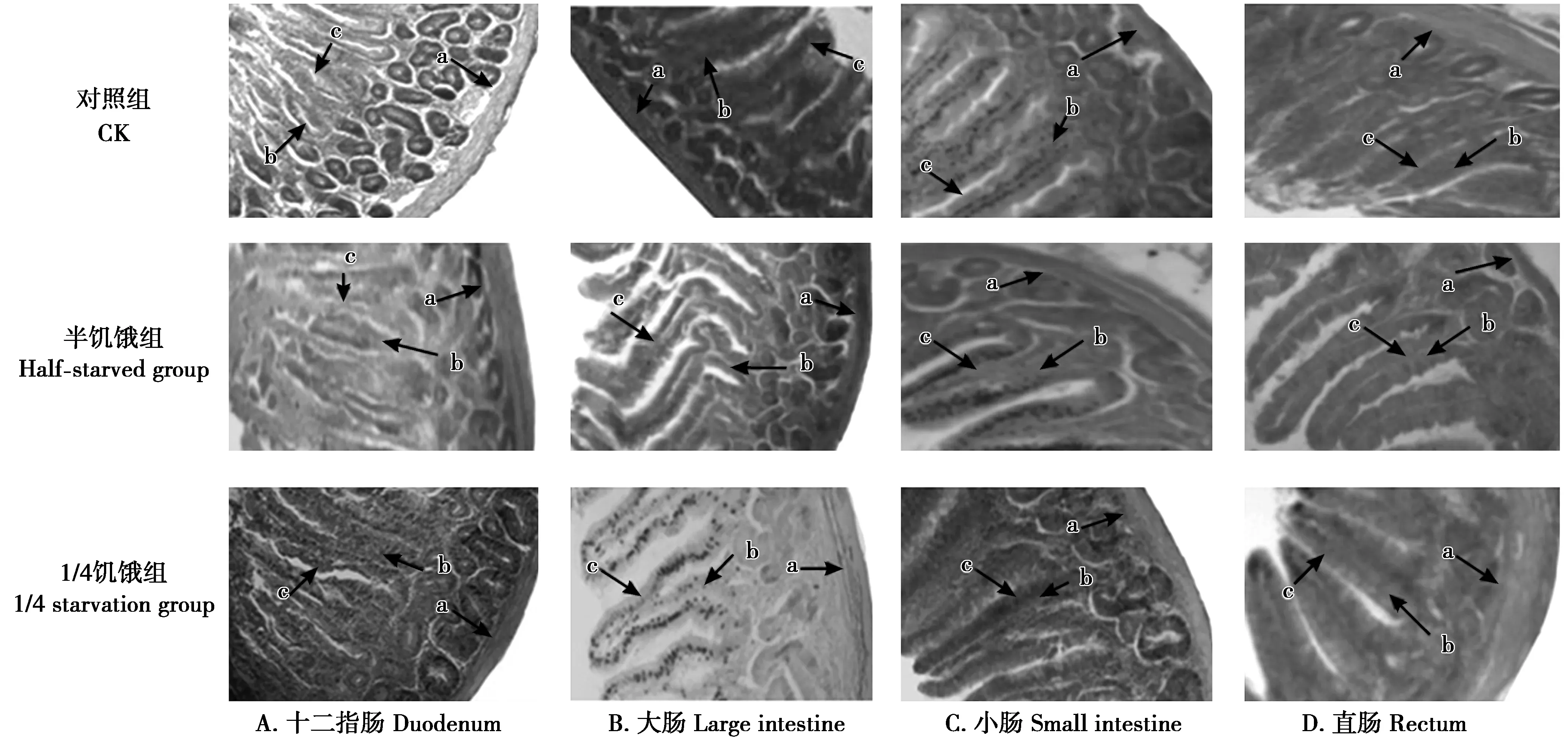
图4 树麻雀消化道石蜡切片(HE×40)
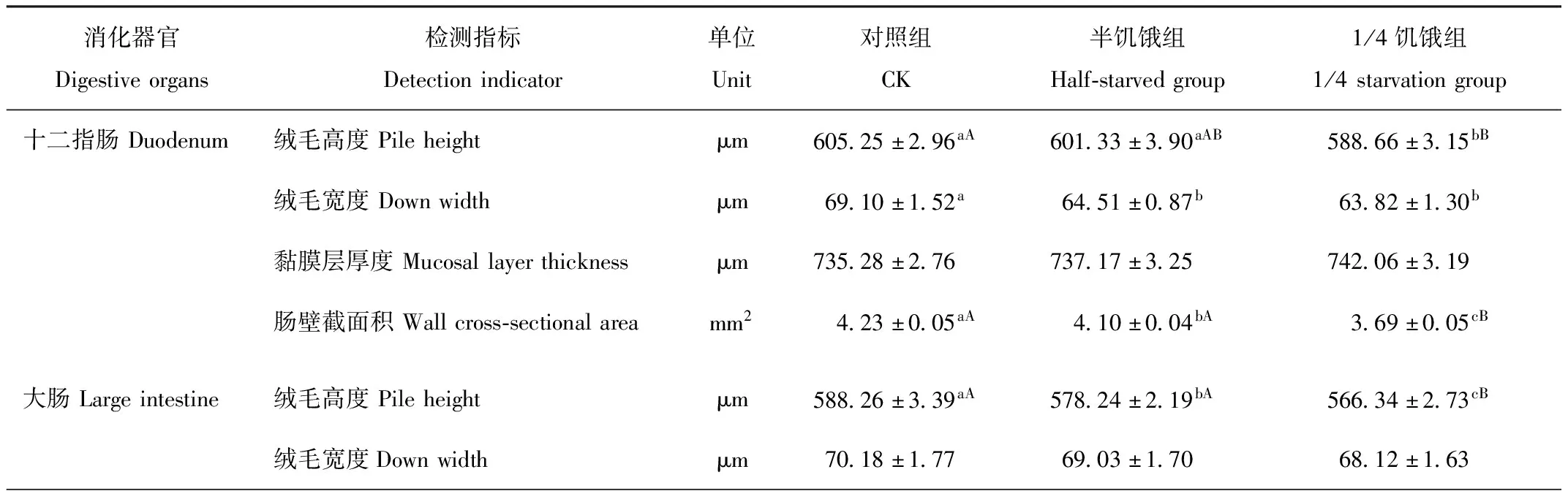
表5 限食对冬季树麻雀消化道绒毛高度、宽度、黏膜层厚度及肠壁截面积的影响
3 讨论
冬季是鸟类觅食压力最大的季节,而觅食压力是鸟类在生存中面临的必然挑战,鸟类生存能力受到能量处理及分配速率的影响[9]。本研究限食条件下,树麻雀体重表现出明显的下降趋势,由于限食使外源性营养物质供应不足,内源性营养在机体代谢中占据主要地位,机体内的储能分子被分解以维持正常的生理活动,最终使体重下降,这与Bales等[10]和赵田等[11]的研究结果相一致。
脂肪用于维持糖异生作用[12],随着限食程度的增加,体内的脂肪被逐渐消耗,使体脂质量分数不断下降。限食条件下,树麻雀消化能增高的原因可能是树麻雀通过提高消化速率增加了有限食物的能量摄入,进而使其增高。由于树麻雀摄食量急剧下降,各组织器官的生理指标也随之发生相应变化以维持基础能耗。
本研究中,限食组树麻雀肝脏、心脏、肺、胰腺鲜重降低,或因为在饥饿胁迫下,大多数动物为维持其基础代谢消耗机体储存的糖原和脂类物质从而使得各器官鲜重有所降低[13]。各代谢器官鲜重的降低直接导致了BMR的降低,表明树麻雀通过减少呼吸代谢,以减少能量的消耗。Aharon-Rotman等[14]研究表明鸟类总代谢率与体重成正比。
由于肌胃具有较强的收缩能力,通过增加重量及容量使食物进行充分混合,促进有限食物的充分消化,故限食组树麻雀肌胃、腺胃鲜重增高。石蜡切片表明,饥饿促使大肠形态结构发生改变,通过降低大肠绒毛高度、宽度,使单位面积内有效的绒毛数量增多,增加有效的吸收面积进而对营养物质充分地吸收,大肠黏膜层厚度的增加是鲜重增加的主要原因。
淀粉酶的分泌量及活力大小与摄食数量的多少密切相关[15],淀粉酶作为消化道内专一的消化酶,其活力的高低决定了对营养物质消化吸收的能力。本研究表明,淀粉酶在各消化器官中均有分布,肠道中的淀粉酶活力的变化幅度较肌胃、腺胃大,说明肠道对限食条件比肌胃、腺胃更敏感,随着限食程度的增加,大肠、小肠、直肠和十二指肠的淀粉酶活力不断下降,可能由于树麻雀在限食条件下,体内代谢发生适应性变化,通过调整各种消化酶的活性,达到合理利用体内贮存物质以维持生命的目的[16-17],这与丁立云等[18]的研究结果相一致。胰脏分泌的胰液中含有大量胰淀粉酶[19],在肠道中分解淀粉等物质,由于限食导致摄入的食物较少,胰淀粉酶的分泌量降低,进而淀粉酶活力降低。肝脏中淀粉酶活性随饥饿程度的增加表现出较高活力的原因,可能是肝糖原在机体内酶促作用下分解为葡萄糖[20-22]进入血液使血糖浓度升高,维持血糖正常水平。限食组树麻雀肝糖原质量分数降低,以此推测,限食使血糖浓度降低[23],肾上腺素和胰高血糖素分泌增加[24],这2种物质与肝脏组织细胞膜受体结合,使无活性的糖原磷酸化酶转化成有活性的磷酸化酶,促进糖原分解为机体提供能量。肝糖原质量分数的变化幅度大于肌糖原质量分数的变化幅度,肝糖原分解用于供能,而肌糖原分解主要维持肌肉收缩。Nord等[25]的研究也表明季节性环境中的动物必须谨慎地管理能源支出以度过冬天,可通过减少用于各种目的的能量分配来实现。限食条件下树麻雀体内消化酶活性的变化可能还与机体本身的能量分配机制及体内能量利用的比例有关,但具体分配机制还有待于进一步研究。
本研究结果显示:冬季限食条件下,树麻雀通过降低BMR,增加消化道长度和黏膜厚度,降低消化道绒毛高度和宽度,加快肝糖原和肌糖原的分解,以及增加消化能来补偿能量消耗等一系列生理生化反应,以适应饥饿寒冷的生存环境。
猜你喜欢
蜜蜂杂志(2022年4期)2022-07-22
文萃报·周五版(2021年34期)2021-09-13
爱你·健康读本(2020年8期)2020-08-23
爱你(2020年22期)2020-08-18
中国医药导报(2019年3期)2019-03-18
人人健康(2017年11期)2017-06-02
现代养生·下半月(2016年6期)2016-10-21
科技传播(2009年17期)2009-10-28
求医问药(2009年7期)2009-08-31
中学生物学(2009年1期)2009-02-23
