
不同尿素施肥量对薄壳山核桃嫁接苗光合作用日变化的影响
2022-02-23 07:29杜洋文邓先珍程军勇
中南林业科技大学学报 2022年1期
杜洋文,邓先珍,程军勇
(1.湖北省林业科学研究院,湖北 武汉 430075;2.黄冈师范学院 a.经济林木种质改良与资源综合利用湖北省重点实验室;b.大别山特色资源开发湖北省协同创新中心,湖北 黄冈 438000;3.湖北省木本粮油林工程技术研究中心,湖北 武汉 430075)
薄壳山核桃Caryaillinoensis为胡桃科山核桃属,又名美国山核桃、长山核桃,是世界著名的干果油料树种之一。目前我国引种栽培地主要集中在江苏、浙江、云南、陕西、安徽、江西、河南和湖南等省[1],种仁含油率达70%以上,是集社会效益、经济效益和生态效益于一体的优良经济树种,具有广阔的国内和国际市场[2]。近年来,国内云南、浙江、安徽、江苏等地对薄壳山核桃干果的需求不断增加,但在实际生产中薄壳山核桃种植并未得到大力推广和实现高产高效,究其原因主要是育种进程缓慢、良种苗木缺乏和配套栽培技术不完善,极大限制了产业发展[2-3]。光合作用是植物生长和果实发育的重要生理基础和生理评价指标,对解释植物响应环境的生理生态学特性具有重要意义[4]。光合作用日变化能够反映一天中植物进行物质积累与生理代谢的持续能力,同时也是分析环境因素影响植物生长和代谢的重要指标,有助于对植物不同品种的光能利用情况和抗逆性进行了解,从而更好地对其进行栽培应用。光合作用受植物内部叶龄、气孔、部位、生育期等生理因子及外部光照、CO2浓度、温度、湿度等环境因素的共同影响,但由于内部生理因子在较短的时间里相对稳定,外部环境因子就成为影响光合作用的主要因素[5]。净光合速率是植物光合作用能力最直接的体现,其大小直接决定着植物光合能力的强弱[6-7]。目前关于薄壳山核桃光合作用的研究主要有韩杰等[8]、杨标等[9]、姜宗庆等[10]、李晓锐等[11]关于修剪措施、干旱胁迫、磷元素以及光合模型模拟等方面对光合特性的影响,汤文华等[12]对不同品种薄壳山核桃的光合特性进行了比较分析,吕芳德等[13]分析了不同树龄薄壳山核桃的光合作用,杜洋文等[14]对薄壳山核桃育苗配方养分需求也做了初步研究。关于尿素不同施肥量对薄壳山核桃嫁接苗苗期光合作用日变化及生理生态因子综合评价方面的研究鲜有报道,本研究主要研究尿素不同施肥量对薄壳山核桃嫁接苗净光合速率及生理生态因子日变化的影响,并运用主成分分析、相关分析、偏相关分析、通径分析、决策分析和聚类分析进行综合评价,以期为薄壳山核桃良种壮苗繁育提供技术支撑。
1 试验地概况
试验地位于武汉九峰试验林场,东经114°29′,北纬31°22′,海拔51~202 m,该区属亚热带大陆季风性湿润气候,年日照时数2 058.4 h,年平均气温16.7℃,极端最高气温41℃,极端最低气温-17.6℃,年平均降水量1 200~1 400 mm,无霜期239 d。土壤以红黄壤为主,土层厚度80 cm以上,土壤pH 值5.3~6.5。
2 材料与方法
2.1 试验设计
光合日变化测定的对象是薄壳山核桃‘波尼’品种2年生嫁接容器苗,2020年5月开始每隔15~20 d 分别浇施0.5%(JJM2)、0.7%(JJM4)、0.9%(JJM6) 和1.1%(JJM8) 的尿素溶液500 mL,以浇施等量清水为对照(JJMCK),共计浇施6 次。每处理10 株,3 次重复。
2.2 测定方法
于2020年9月11日晴朗天气,采用美国LICOR 公司的第三代气体交换测量系统Li-6400XT光合仪,分别测定各施肥处理的净光合速率和生理生态因子的日变化,从7:00—17:00 每隔2 h 测定1 次,每处理选择生长势的植株3 株,均选择植株中部健康完好的当年生叶片进行测定,结果取其均值。主要测定指标有:叶片净光合速率(Pn,μmol·m-2s-1)、气孔导度(Gs,mmol·m-2s-1)、蒸腾速率(Tr,mmol·m-2s-1)、胞间CO2浓度(Ci,μmol·mol-1)、光合有效辐射(PAR,μmol·m-2s-1)、大气CO2浓度(Ca,μmol·mol-1)、大气温度(Ta,℃)、叶片温度(Tl,℃)、空气相对湿度(RH,%)、叶面饱和蒸气压亏缺(Vpdl,MPa)等;利用光合和环境参数计算气孔限制值(Ls=1-Ci/Ca)和水分利用率(WUE=Pn/Tr),计算方法参见文献[15]。
2.3 数据分析
利用SPSS 16.0 软件对生理和生态因子进行主成分分析,再对各生理生态因子及Pn进行相关分析及偏相关分析,进一步进行通径分析和决策分析,最后进行聚类分析。
3 结果与分析
3.1 不同施肥量对薄壳山核桃嫁接苗净光合速率日变化的影响
由图1可知,4 个不同施肥浓度和对照处理的Pn日变化曲线基本相似,曲线呈不规则抛物线。其中JJM2、JJM4、JJM6 和JJMCK 处理日变化呈升高—降低的单峰曲线,无“午休”现象;JJM8日变化呈升高—降低—升高—降低的双峰曲线,在13:00 出现“午休”现象,二者第1 个峰值均在11:00 出现,JJM8 第2 个峰值在15:00 出现。5 个处理的Pn日变化总体表现为JJM4>JJM8>JJM6>JJM2>JJMCK,但Pn最高峰值表现为JJM8(12.215 μmol‧m-2s-1)>JJM4(11.783 μmol‧m-2s-1)>JJM6(9.821 μmol‧m-2s-1)>JJM2(9.244 μmol‧m-2s-1)>JJMCK(7.518 μmol‧m-2s-1)。其中JJM4 的Pn在日变化中基本都高于其他处理,表现出了较强的光合能力;JJM8 的Pn日变化曲线表现为快升—快降,在“午休”后,表现为慢升—慢降;JJMCK的Pn在日变化过程中都处于较低水平,对照的光合能力明显低于施肥处理。

图1 不同施肥量对薄壳山核桃嫁接苗净光合速率日变化的影响Fig.1 The diurnal changes of the net photosynthetic rate
3.2 不同施肥量对薄壳山核桃嫁接苗生理生态因子日变化的影响
由图2可知,气孔导度和胞间CO2浓度与净光合速率曲线呈相反的变化趋势,曲线呈“U”形。气孔导度日变化曲线总体呈降低—升高—缓慢降低的趋势,具体表现为7:00—9:00 下降,9:00—11:00 上升,11:00—17:00 缓慢下降,各处理间表现 为JJM2>JJM6>JJM8>JJMCK>JJM4;胞间CO2浓度日变化曲线总体呈降低—平缓—升高的趋势,具体表现为降低(7:00—9:00)—平缓(9:00—15:00)—升高(15:00—17:00),各处理间表现为JJMCK>JJM8>JJM4>JJM2>JJM6。蒸腾速率、水分利用率、气孔限制值与净光合速率日变化曲线基本相似,呈不规则抛物线,但这3 个生理因子日变化曲线间存在一定差异,其中蒸腾速率日变化曲线表现为升高—降低,JJM4和JJMCK 在7:00—13:00 升高,13:00—15:00 缓慢降低,15:00—17:00 急剧降低,JJM8、JJM2 和JJM6 在7:00—15:00 升高,15:00—17:00 急 剧 降低,各处理间表现为JJM4>JJMCK>JJM8>JJM2>JJM6;水分利用率日变化曲线表现为升高(7:00—9:00)—降低(9:00—13:00)—平缓(13:00—15:00)—升高(15:00—17:00),各处理的日变化曲线基本一致,相对大小为JJM6>JJM8>JJM2>JJM4>JJMCK;气孔限制值日变化曲线表现为升高(7:00—9:00)—平缓波动(9:00—15:00)—下降(15:00—17:00),各处理的日变化曲线趋势基本一致,相对大小为JJM6>JJM2>JJM8>JJM4>JJMCK,其中JJM6 和JJM2 的曲线变化更为相似,JJM8 和JJMCK 的曲线变化更为相似,JJM4 则介于二者之间。

图2 不同生理生态因子的日变化曲线Fig.2 The diurnal changes of various physiological and ecological factors
薄壳山核桃嫁接苗的叶面饱和蒸汽压亏缺和叶片温度及空气温度日变化曲线呈倒“V”形,表现为逐步升高(7:00—15:00)—急剧降低(15:00—17:00),峰值出现在15:00,与净光合速率日变化曲线形状基本一致,但曲线峰值出现时间存在较大差异;空气CO2浓度日变化曲线为降低(7:00—13:00)—升高(13:00—15:00)—降低(15:00—17:00);大气相对湿度日变化曲线呈“V”形,峰谷出现在15:00,与叶面饱和蒸汽压亏缺、叶片温度及空气温度峰值出现时间相同,与净光合速率日变化曲线相反,这也说明了两者之间存在一定的相关关系。
3.3 影响薄壳山核桃嫁接苗净光合速率日变化的生理生态因子分析
3.3.1 生理生态因子的主成分分析
将对薄壳山核桃嫁接苗净光合速率Pn日变化产生影响的因子分为生态因子PAR、Ta、Ca、RH、Vpdl、Tl和生理因子Gs、Ci、Tr、WUE、Ls,并分别对生态因子和生理因子进行主成分分析(表1)。对生态因子分析可知,仅只有1 个主成分特征值大于1,累计贡献率为83.343%,该主成分的特征向量以Ta和Tl最大,其次为Vpdl和RH,表明这4 个因子为主导因子,其中RH 为限制性因子,称第1 主成分为温度、气压和湿度因子,它们在影响薄壳山核桃嫁接苗光合作用日变化的生态因子中处于主导地位;PAR 和Ca主要为光照和CO2因子,在生态因子中处于次要地位。对生理因子分析可知,特征值大于1 的主成分主要有两个,分别为第1 主成分和第2 主成分,累计贡献率为84.909%,第1 主成分的特征向量以Ls最大,其为限制性因子,其次为Ci和Gs,称第1 主成分为气孔和胞间CO2浓度因子,它们在影响薄壳山核桃嫁接苗光合作用日变化的生理因子中处于主导地位;第2 主成分的特征向量以Tr最大,其为限制性因子,其次为WUE,综合表明对净光合速率日变化影响的生理因子以气孔限制值、胞间CO2浓度和气孔导度为主导因子,蒸腾速率和水分利用率为次要因子。
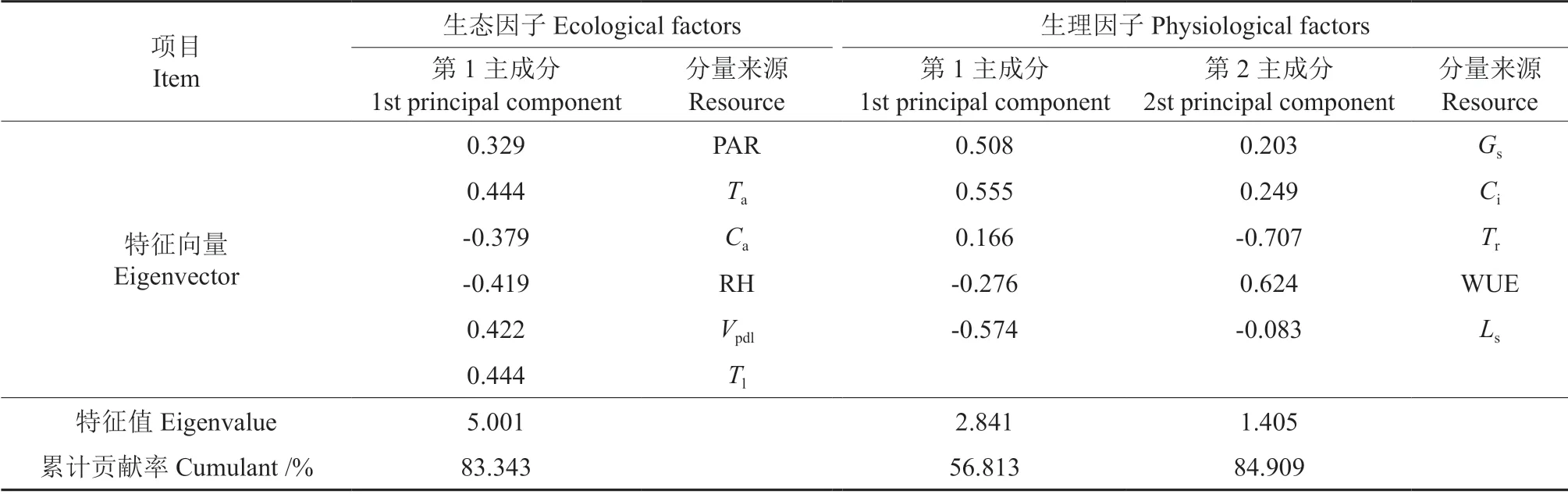
表1 光合作用日变化的生理生态因子主成分分析Table 1 Principal component analysis of physiological and ecological factors in diurnal changes of photosynthesis
3.3.2 生理生态因子的相关分析
对生态因子PAR、Ta、Ca、RH、Vpdl、Tl和生理因子Gs、Ci、Tr、WUE、Ls及Pn进行简单相关分析(表2)可知,薄壳山核桃嫁接苗Pn与Ci存在显著负相关(P<0.05),与Vpdl存在极显著负相关(P<0.01),与Tr、WUE、RH、PAR 存在极显著正相关(P<0.01),与Ls存在显著正相关(P<0.05)。
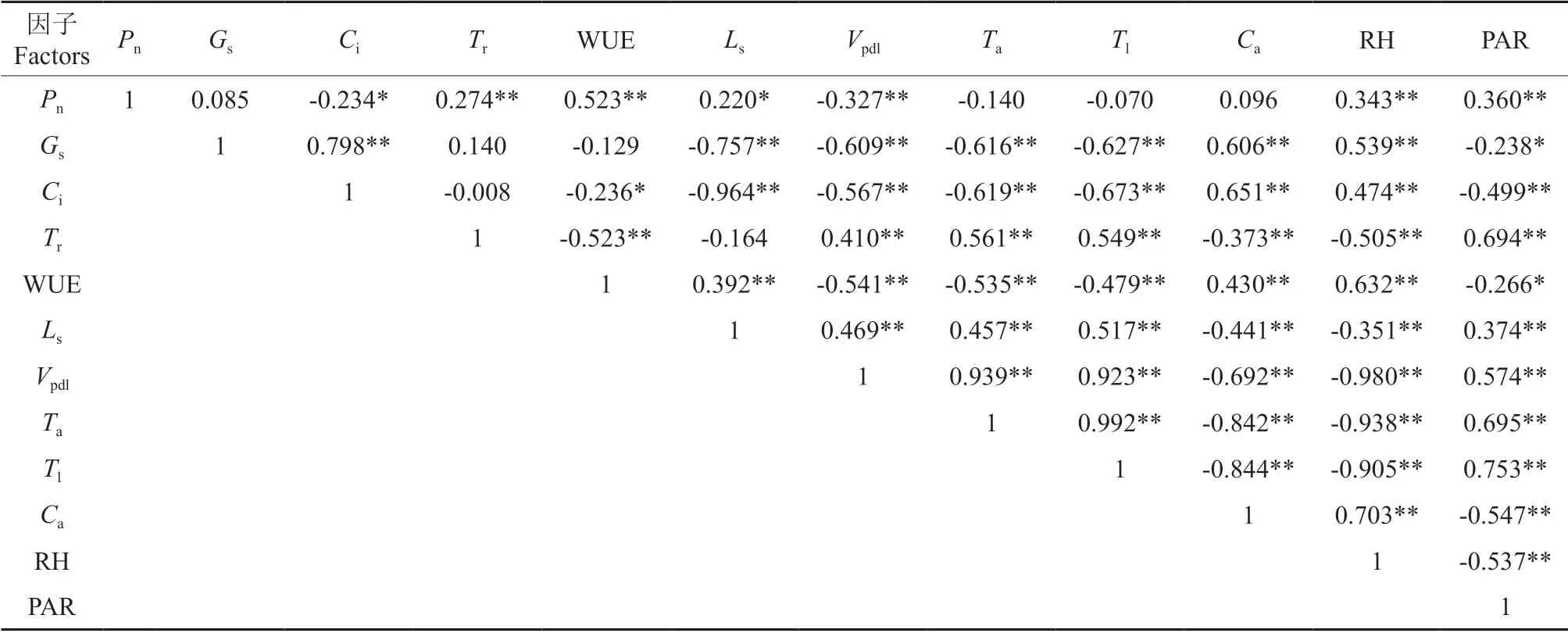
表2 生理生态因子的相关分析†Table 2 Correlation analysis between net photosynthetic rate and physiological and ecological factors
偏相关分析是指当2 个变量同时与第3 个变量相关时,将第3 个变量的影响剔除,只分析这2 个变量之间的相关程度。由于简单相关关系不能很好地反映多因子影响下的相关性,为进一步确定生理生态因子对薄壳山核桃嫁接苗净光合速率的影响程度,对生态因子PAR、Ta、Ca、RH、Vpdl、Tl和生理因子Gs、Ci、Tr、WUE、Ls与Pn分别进行偏相关分析(表3)。结果表明,净光合速率与Ci、WUE、Ls、Vpdl、Ca、RH、Tr呈显著偏相关,Gs、Ta、Tl、PAR 对其影响不显著,同时表明Ca、Ci和Ls对净光合速率影响最大,WUE、Vpdl、RH 和Tr对其影响相对较小。Ca、Ci和Ls偏相关系数明显大于简单相关系数,表明在其他因子变化的情况下,掩盖了Ca、Ci、Ls对Pn的真实影响效果。

表3 净光合速率与生理生态因子的偏相关分析Table 3 Partial correlation analysis between Pn and eco-physiological factors
3.3.3 生理生态因子的通径分析
通过对影响净光合速率的生理生态因子PAR、Ta、Ca、RH、Vpdl、Tl、Gs、Ci、Tr、WUE、Ls进行多元线性回归分析,得到回归方程Pn=52.750-0.587WUE+0.359Vpdl+0.581RH+3.652Ca-12.496Ci-9.753Ls(R2=0.982,P<0.01),表明WUE、Vpdl、RH、Ca、Ci和Ls是影响Pn的主要因子。进一步对这些因子与Pn的作用进行通径分析,可知直接作用绝对值大小排序为Ci>Ls>Ca>WUE>RH>Ta>Vpdl(表4),Ci对Pn的直接作用最大,且为负作用,与二者的相关性一致,说明Pn的降低可能是由非气孔因素导致的。间接通径系数表明,Ci是通过Ls(9.402)和Ca(2.377)作用于Pn,Ls是通过Ci(12.046)和Ca(-1.611)作用于Pn,Ca是通过Ci(-8.135)和Ls(4.301)作用于Pn,三者相互作用的间接通径系数最大,表明Ci、Ls、Ca和Pn间具有密切关系,是影响Pn的主要因子。WUE、Vpdl、RH 和Ta虽然具有相对较大的相关系数,但其直接通径系数的绝对值均小于通过Ca、Ci、Ls对Pn的间接通径系数的绝对值,表明它们对Pn的直接作用小于通过Ca、Ci、Ls的间接作用。
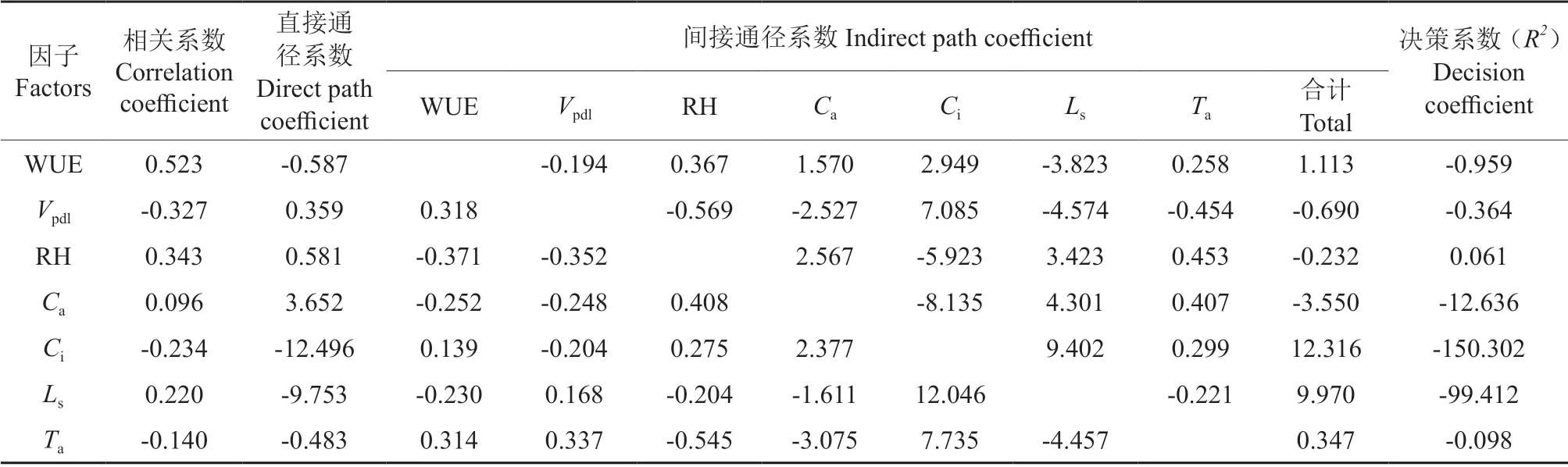
表4 光合生理生态因子的通径分析Table 4 Path analysis of Pn and eco-physiological factors
决策系数(R2)是通径分析中的决策指标,用它可以把各自变量对因变量的综合作用进行排序,排序最大的变量为主要决策变量,但未必它的直接决定作用大;排序最小的变量,若其决策系数为负,则为主要限制性变量,但未必它的直接决定作用小[16]。分析表明,各因子的决策系数R2大小顺序为RH>Ta>Vpdl>WUE>Ca>Ls>Ci,可知,RH 决策系数最大,为主要决策变量(直接决定作用较小),其原因是它通过Ca、Ls和Ta的间接作用分别为2.567、3.423 和0.453,协助RH 起增进作用,WUE(-0.371)、Vpdl(-0.352)和Ci(-5.923)对它起较大的限制作用。其余变量决策系数均为负值,以Ci和Ls最小,为主要限制性变量(直接作用最大),原因是它通过其他变量的间接作用为较大正值;Ca次之,也为限制性变量。因此,要保持较高的净光合速率,必须增加空气相对湿度,限制胞间CO2浓度、气孔限制值和大气CO2浓度,其他变量基本保持不变。
3.3.4 不同施肥量对薄壳山核桃嫁接苗光合系统的聚类分析
以Pn和Gs、Ci、Tr、WUE、Ls、Vpdl、Ti、Tl、Ca、RH、PAR 等11 个生理生态因子为指标进行标准化数据处理,采用离差平方和进行聚类分析(图3)。从图3中可知,可将5 种不同施肥量的薄壳山核桃嫁接苗的光合作用分为三类:第一类是0.5%、0.7%和0.9%施肥量,第二类是1.1%施肥量,第三类是不施肥。

图3 不同施肥量的薄壳山核桃光合系统聚类分析Fig.3 Clustering analysis of Carya illinoensis photosynthesis under different treatments
4 讨 论
一般条件下植物的光合作用日变化呈“双峰型”或“单峰型”的变化趋势[17-19],薄壳山核桃净光合速率日变化曲线总体表现为升高—降低的单峰曲线,无“午休”现象,这与汤文华等[12]的研究结果一致。净光合速率变化是受气孔或非气孔因素调节的[20],JJM8 处理呈双峰曲线,随着净光合速率的降低,气孔导度下降,胞间CO2浓度增高,说明净光合速率受非气孔因素限制[21],这也与许大全[22]的胞间CO2浓度升高是判断非气孔限制的可靠依据的结果一致,这可能是高温和相对湿度的下降对叶片的光合系统造成损伤,使Rubisco 等关键酶活性和叶肉细胞光合活性降低,造成净光合呼吸速率降低[23-24]。蒸腾速率、水分利用率、气孔限制值、净光合速率和叶面饱和蒸气压亏缺、叶片温度和空气温度日变化呈不规则抛物线,与净光合速率日变化曲线基本一致,表明薄壳山核桃苗木对环境条件的变化有较好的适应能力[25-26]。
主成分分析、偏相关分析、回归分析等统计分析表明,影响薄壳山核桃净光合速率的主导生态因子为Ta、Tl、Vpdl和RH,主导生理因子为Ls、Ci和Gs,其中RH 和Ls为限制性因子;净光合速率与Ca、Ci和Ls有较大的偏相关系数;WUE、Vpdl、RH、Ca、Ci和Ls是影响净光合速率的主要因子。进一步进行通径分析和决策系数表明,Ci、Ls和Ca在净光合速率日变化中起主要作用,RH、Vpdl和Ca的直接通径系数远大于间接通径系数,表明RH、Vpdl和Ca通过自身的直接作用对净光合速率产生影响,WUE、Ci、Ls和Ta直接通径系数小于间接通径系数,表明其通过其他生理生态因子的间接作用对净光合速率产生影响。决策系数为通径系数的决策指标,决策系数可以确定主要决策变量和限制变量[27-29]。本试验表明RH 为主要决策变量,WUE、Vpdl、Ca、Ls和Ta为辅助限制性变量,Ci为主要限制性变量。因此,通过提高RH,可以使薄壳山核桃保持较高的净光合速率,另外,由于胞间CO2浓度大小取决于空气CO2浓度大小,通过限制空气CO2浓度,也能提高薄壳山核桃净光合速率。聚类分析将不同施肥浓度分为三类:第一类是0.5%、0.7%和0.9%施肥量,称为低施肥水平;第二类是1.1%施肥量,称为高施肥水平;第三类是不施肥。由于本研究仅限于薄壳山核桃嫁接苗在生长期对不同尿素施肥量的光合响应,下一步将继续开展N、P、K 等多元素形成的养分配方对苗木生长和光合生理等方面的影响的研究。
5 结 论
不同施肥量下薄壳山核桃嫁接苗净光合速率的日变化曲线总体呈不规则抛物线,各处理间净光合速率也存在一定差异。浇施氮肥能显著提高叶片的净光合速率和增强光合作用,相同施肥量对不同生理生态因子的日变化值的影响程度及大小存在差异,不同施肥量对同一生理生态因子相同时刻的影响也存在差异。经主成分分析、相关分析、通径分析和聚类分析可知,空气相对湿度为影响薄壳山核桃净光合速率的主要决策因子,胞间CO2浓度为主要限制性因子,以0.9%和1.1%较高施肥水平有利于薄壳山核桃嫁接苗提高光合效率和增强光合效能。
猜你喜欢
中国蔬菜(2022年6期)2022-06-22
农业工程学报(2022年4期)2022-04-24
数学大王·中高年级(2021年3期)2021-03-10
农业与技术(2020年12期)2020-07-04
阅读(中年级)(2018年11期)2018-01-19
新课程·下旬(2017年7期)2017-08-14
现代农业研究(2017年6期)2017-07-17
农家顾问(2016年4期)2016-11-19
