
不同水分条件下小叶章光响应曲线变化
2022-02-14 13:21王建波付晓玲刘赢男倪红伟
黑龙江科学 2022年2期
王建波,付晓玲,刘赢男,倪红伟
(黑龙江省科学院自然与生态研究所,哈尔滨 150040)
0 引 言
水分是影响植物生长和功能的主要生境因子[1],水分胁迫也是抑制植物光合作用的主要环境因子之一[2]。小叶章是禾本科野青茅属多年生根茎型草本植物,在我国主要分布于东北、华北、内蒙古等地区的平原低湿地,尤以三江平原分布最为集中[3-4]。该植物水分适应范围较广,为三江平原典型草甸、沼泽化草甸的建群植物和优势植物,沼泽植被中的优势种、亚优势种或重要的伴生种[5]。小叶章在东北草地地区的饲用价值仅次于羊草,同时也是很好的水土保持植物和造纸等轻工业的良好原料[6]。
光合作用是植物对外界环境因素最敏感的生理过程之一。植物光合光响应曲线反映的是净光合速率随光强变化的特性,通过拟合光响应曲线得到的光合速率、表观量子效率、暗呼吸速率、光饱和点和光补偿点等生理参数对了解植物光反应过程非常重要[7],也是判定植物光合机构运转状况、光合作用能力、光合利用率及受环境变化影响程度的依据[8]。目前,关于小叶章在自然水分条件下的光合生理生态特性有一定研究[9-10],但对于其在干旱和水淹条件下的光合情况并不清楚。通过设置不同水分处理,探讨小叶章对不同水分的光合光响应规律及其差异,为小叶章湿地的退化恢复、保护管理等提供科学理论依据。
1 材料与方法
1.1 研究区域概况
研究地点设置在黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站。该研究站位于洪河国家级自然保护区,地处三江平原的东北部,属温带季风气候,多年平均气温为1.9℃,年均降水量为585 mm,50%~70%集中在夏季[11]。依据小叶章湿地水分差异可划分为:小叶章典型草甸,土壤含水量60%~80%,以小叶章为优势种;小叶章沼泽化草甸,土壤含水量80%~120%,呈现季节性积水,以小叶章为优势种;小叶章沼泽,地表常年积水,积水深度5~20 cm,以漂筏苔草(Carexpseudo-conica)和小叶章为共优势种[12]。
1.2 试验设计
采用盆栽控制实验。于2010年5月在黑龙江省科学院自然与生态研究所三江平原湿地生态学实验站采集15个原状土块,体积约为50 cm×50 cm×50 cm,保持原状放置于内部尺寸60 cm×60 cm×80 cm、壁厚1 cm的有机玻璃箱内进行土壤含水量控制实验。试验分为5个处理,分别为极度干旱(土壤含水量20%,ED)、典型草甸(土壤含水量70%,TM)、沼泽化草甸(土壤含水量100%,SM)、沼泽(积水深度10 cm,M)、极度水淹(积水深度30 cm,EF)。土壤含水量采用称重法进行测定与控制。水分处理自2010年6月1日开始,至9月1日结束。
1.3 测定项目与方法
于水分处理两个月后,选择天气晴朗的上午9∶00~12∶00,采用Li-6400便携式光合系统(Li-Cor,Inc,美国)测定叶片光合作用光响应曲线。每一处理随机选择5个株龄相似、节位一致的植物功能叶片,使用开放气路,空气流速为500 μmol·s-1,温度25℃~27℃,相对湿度80%,CO2浓度由小钢瓶提供,浓度设定为400 μmol·m-2s-1。用LI-6400自带红蓝光源模拟光强,测定从2000 μmol·m-2s-1光量子的光强开始,依次降为l 500、1 000、800、600、400、200、100、50和0 μmol·m-2s-1。测定时每一光强下停留3 min,改变光强后,最少稳定时间设置为60 s,当测量结果变异率小于0.05时由仪器自动记录。光合参数根据非直角双曲线模型进行拟合计算。
1.4 数据统计与分析
采用Sigmaplot 2000软件对数据进行处理及作图,采用SPSS 17.0软件进行单因素方差分析及其差异显著性检验。
2 结果与分析
2.1 水分处理对小叶章光合-光响应曲线的影响
由图1可见,当光合有效辐射PAR小于200 μmol·m-2s-1时,随着PAR的增加,小叶章叶片的Pn随着光强呈线性增加趋势。此后,随着PAR的继续增加,Pn在各个处理下的增加幅度放缓慢。不同水分处理下,当PAR>400 μmol·m-2s-1时,Pn-PAR曲线变化规律开始出现差异,其中以TM处理的Pn-PAR实测曲线变化最高,EF处理最低。当光强达到1 000 μmol·m-2s-1时,Pn渐进饱和。
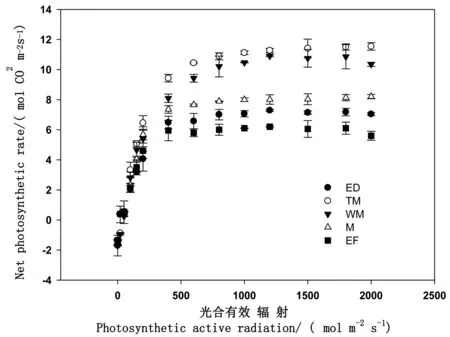
图1 不同水分处理下小叶章的光合光响应曲线Fig.1 Pn-PAR curves of Calamagrostis angustifolia under different water conditions
2.2 水分处理对小叶章光合参数的影响
表1显示光响应曲线特征参数,与TM相比,WM、M、EF和ED处理的Amax,LSP、Rd及AQY均呈现下降趋势。具体而言,WM处理各光合参数与TM相比差异不显著;而M处理的Amax、AQY、LSP、LCP、Rd分别比TM处理降低了28%、39%、13%、26%、39%,差异达到显著水平;与M处理相似,EF的Amax、AQY、LSP、LCP和Rd分别比典型草甸降低46%、31%、30%、23%和42%。对于EF处理,光合参数Amax、AQY、LSP和Rd也呈现显著下降的趋势,分别降低了37%、36%、12%和37%。结果表明,M、ED、EF处理光响应参数Amax、Rd、AQY、LSP、LCP均显著低于TM和WM。
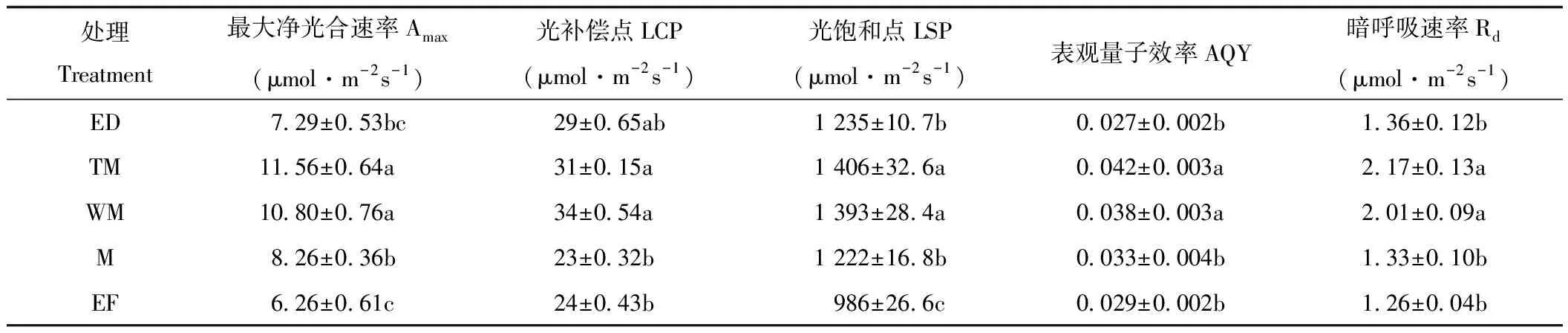
表1 不同水分条件下小叶章光响应曲线参数Tab.1 Pn-PAR curve parameters of Calamagrostis angustifolia under different water supplies
3 讨论
光是植物赖以生存和生长的重要环境因子之一。光合作用光响应曲线可判断植物光合效率受环境变化的影响程度,也是研究植物在逆境条件下的重要手段之一[13]。研究发现,小叶章的光响应曲线中,最大净光合速率Amax在模拟沼泽、极度水淹和极度干旱条件下均显著低于典型草甸,说明沼泽小叶章、极度水淹和极度干旱均影响了小叶章Rusbico活性及电子传递速率。
AQY反映了叶片对光能的利用情况,尤其是对弱光的利用能力,其值越大,表明植物吸收与转换光能的色素蛋白复合体越多,利用弱光的能力就越强[14]。研究认为,植物AQY的理论值在0.083~0.125[15],而在自然条件下远低于理论值(0.04~0.07)。本研究中,不同水分条件下小叶章的AQY在0.027~0.042。水分胁迫通常造成AQY的降低,与典型草甸相比,M、EF、ED三个水分处理的AQY均显著降低,说明在干旱和淹水水分胁迫条件下,小叶章在利用弱光能力方面受到明显抑制。
小叶章叶片Rd在M、EF和ED三个处理条件下也显著降低,表明小叶章主要通过降低呼吸作用来适应逆境胁迫的土壤条件。与此相似,有研究发现芦苇等植物在水淹后会降低Rd,以应对光合能力的下降[16]。但也有一些研究表明,植物Rd在水淹条件下会增加,主要是由于植物同化产物运输受到抑制,增加Rd会消耗叶片中积累的过多同化产物,减缓对光合作用的抑制[17],这表明不同的植物对逆境胁迫的光合生理适应性存在较大差异。
光合参数LSP和LCP是分别衡量植物利用强光与弱光能力的指标,通常来说,植物具有较高的LSP和较低的LCP对光的生态适应能力更强[18-19]。研究发现,当水分过高时,植物会增加LCP,而降低LSP,利用弱光和强光的能力均会降低[20-21]。本研究表明,小叶章在TM、WM条件下的LCP和LSP最高,在M、EF和ED处理中,显著降低了LSP,表明在沼泽、极度干旱和极度水淹条件下,小叶章利用强光的能力降低,缩小了其光的适应能力范围。一般来说,典型阳生植物LCP的范围为9~27 μmol·m-2s-1,LSP的范围为360~900μmol·m-2s-1[22]。5个水分处理的小叶章LCP在23~34 μmol·m-2s-1,而LSP均大于900 μmol·m-2s-1,说明小叶章更适合在阳坡生长。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
汽车实用技术(2022年16期)2022-09-03
青海草业(2022年2期)2022-07-23
测井技术(2022年3期)2022-07-16
福建农林大学学报(自然科学版)(2021年5期)2021-10-08
海外文摘·文学版(2021年3期)2021-05-19
汽车实用技术(2019年20期)2019-11-26
网络安全和信息化(2019年6期)2019-06-28
活力(2019年21期)2019-04-01
科技视界(2018年8期)2018-06-08
