
长江口凤鲚繁殖群体年龄与生长研究
2022-02-11 03:32:58黄承伟李增光胡永斌
海洋渔业 2022年6期
黄承伟,万 荣,李增光,胡永斌
(上海海洋大学海洋科学学院,国家远洋渔业工程技术研究中心,大洋渔业资源可持续开发教育部重点实验室,上海 201306)
凤鲚(Coiliamystus)隶属鲱形目(Clupeiformes),鳀科(Engraulidae),主要分布区域在西太平洋,包括中国、朝鲜半岛和日本的海域,其中我国渤海、黄海、东海和南海均有分布[1]。凤鲚属于河口区洄游性鱼类,袁传宓等[2]认为,我国长江中下游和东南沿海凤鲚有3个生态类群,即长江型、闽江型和珠江型,其中长江口凤鲚在春、夏繁殖季节,达到性成熟的繁殖群体从近岸浅海集群洄游至崇明附近的长江口咸淡水区域产卵形成鱼汛[1]。20世纪八十年代,凤鲚平均年产量约占长江口鱼虾类水产品总产量的48.6%,是长江口重要的经济鱼类[3]。近年来,由于水体污染、过度捕捞等因素造成凤鲚资源严重破坏,已不能形成鱼汛[4],且繁殖群体呈现小型化、低龄化趋势[5],表明长江型凤鲚种群出现严重衰退,亟需开展相关的资源评估与保护工作。
2008年之前,大多研究根据鳞片、脊椎骨等传统材料鉴定长江口凤鲚年龄,并普遍认为其年龄结构简单,几乎只有1龄个体,2龄个体鲜见,种群几乎全部由补充群体组成[6-10]。HE等[11]提出了一种可以通过凤鲚耳石切片上特殊的直/弯生长轴线判定年龄的方法,认为长江口凤鲚繁殖群体有5个年龄组(1~5龄),其中1、2龄个体为优势群体。王焕焕等[5]、于晓[12]和毕雪娟[13]也先后采用凤鲚耳石直/弯生长轴线进行了长江口凤鲚年龄结构的研究,获得了大体类似的研究结果。施炜纲和王博[7]提出长江口凤鲚优势体质量区间从20世纪八十年代初的15~22 g下降至九十年代末的6~14 g,认为凤鲚可能存在个体小型化趋势[3]。2003年三峡大坝蓄水后,凤鲚繁殖群体的全长和体质量分布区间重心继续呈现小型化趋势[14],并在2012—2013年出现体长和体质量平均值的历史最低点[4]。
文献检索表明,自2016年至今未见关于长江口凤鲚繁殖群体的年龄结构和生长特性的相关报道。在此背景下,本文利用2018年5—7月和2019年5月采集到的凤鲚繁殖群体样品数据,开展了长江口凤鲚繁殖群体年龄结构与生长特征的研究,以期为开展长江全面禁捕对长江口凤鲚资源保护与管理的效果评估提供基础数据。
1 材料与方法
1.1 样品采集
实验样品的采集和测定均按照《海洋调查规范—第6部分:海洋生物调查》(GB/T 12763.6—2007)[15]进行,分别在2018年5—7月以及2019年5月对长江口水域(图1)采用定置刺网捕获到的凤鲚繁殖群体进行采样,若单网次渔获总重大于30 kg,则随机取样30 kg;否则,全部取样。在实验室再按10%的比例随机取样,对样品进行全长(mm)、体长L(mm)、体质量W(g)等生物学参数的测定。解剖后,采用目测法确定凤鲚的性别。

图1 长江口凤鲚采样站点
1.2 耳石数据测定
对凤鲚左右矢耳石重量进行显著性差异检验,结果表明无显著性差异(P>0.05)。因此,本实验统一采用凤鲚左耳石(图2),用精度为0.1 mg的微量天平(FA1204B)测定耳石质量(weight of otolith,WO)。采用传统耳石形态测量法[16],使用Nikon Zoom645S体式显微镜测量耳石长(otolith length,OL)、耳石宽(otolith width,OW)、耳石周长(otolith perimeter,OP)和耳石面积(otolith area,OA);采用凤鲚矢耳石直/弯生长模式的方法判定凤鲚年轮(图3)[11],并根据年轮数量、捕捞月份和耳石边缘直/弯类型确定凤鲚年龄[11]。

图2 凤鲚左耳石形态示意图

图3 凤鲚年龄鉴定图
1.3 年龄与生长分析
体长和体质量的关系式为:W=aLb。采用t检验比较b值与“3”之间是否存在显著性差异[17]。采用Von Bertalanffy生长方程:Lt=L∞[1-e-k(t-t0)]和Wt=W∞[1-e-k(t-t0)]b表述体长、体质量和年龄的关系。式中,t为年龄,Lt和Wt为t龄时的平均体长和体质量,L∞和W∞为平均渐进体长和体质量,k为生长系数,t0为假设的理论生长起点年龄。对体长和体质量生长方程分别进行一阶和二阶求导得到其对应的生长速度方程和加速度方程,生长拐点方程为:ti=lnb/k+t0。式中,ti为生长拐点年龄[18]。
1.4 数据分析方法
所有实验数据均采用Microsoft Excel 2019和SPSS 25.0处理,图像使用上海银瑞信息科技有限公司开发的YRMV图像测量软件处理。
2 结果与分析
2.1 年龄结构及耳石形态特征
凤鲚耳石样品测定结果表明,长江口水域凤鲚繁殖群体由1~3龄3个年龄组组成。其中,1龄样品142尾,占总样品数73.20%;2龄样品18尾,占总样品数9.28%;3龄样品34尾,占总样品数17.52%。1、2龄个体合计占比82.48%,为优势群体。
表1是凤鲚耳石形态学参数及其质量。协方差分析(ANCOVA)表明,耳石长OL、耳石宽OW、耳石周长OP、耳石面积OA与耳石质量WO之间无显著的线性相关(P>0.05)。线性回归分析表明,耳石长OL、耳石宽OW、耳石周长OP、耳石面积OA与耳石质量WO之间呈现显著的线性相关(P<0.01)(图4)。

表1 凤鲚耳石形态学参数及其质量
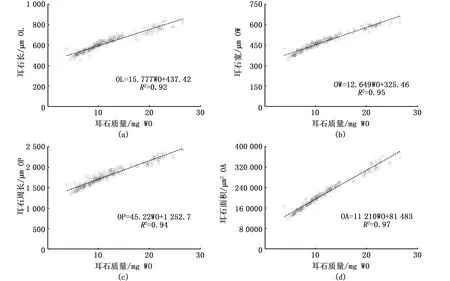
图4 凤鲚耳石各参数与耳石质量关系
2.2 生长分析
2.2.1 体长-体质量关系
凤鲚可鉴别性别的样品共194尾,其中雌性135尾,雄性59尾,雌雄性比为2.29∶1。雌性凤鲚体长范围为82~192 mm,均值为(131.73±24.54)mm,雄性凤鲚体长范围为69~179 mm,均值为(122.31±25.87)mm(图5)。经配对t检验(paired-ttest),雌性数量显著大于雄性数量(P<0.05)。经协方差分析,雄性平均体长显著小于雌性平均体长(P<0.05)。凤鲚总体体长范围为69~192 mm,主要集中在95~175 mm之间,体长平均值为(128.87±25.26)mm;体质量范围为1.5~30.3 g,主要集中在3.5~11.5 g之间,体质量平均值为(9.90±6.61)g。对3个年龄组进行多重比较的结果显示,3个年龄组间的体长和体质量平均值均存在明显差异(P<0.05)(表2)。

图5 凤鲚体长和体质量分布

表2 长江口凤鲚各年龄组的体长和体质量
如图6所示,长江口凤鲚繁殖群体的体长和体质量存在显著的幂函数关系。其中,W=2×10-6L3.17(R2=0.95,n=194),W=9×10-7L3.31(R2=0.95,n=135♀),W=7×10-6L2.87(R2=0.95,n=59♂)。经t检验,雄性和样品总体的幂函数指数b(2.87和3.17)与3无显著差异(P>0.05),雌性的幂函数指数b(3.31)与3有显著差异(P<0.05),即雌性凤鲚表现出正异速生长特性,而雄性凤鲚为匀速生长类型。

图6 凤鲚体长和体质量关系
2.2.2 生长方程
凤鲚体长和体质量生长方程分别为:Lt=212.33[1-e-0.43(t+0.88)]和Wt=48.22[1-e-0.43(t+0.88)]3.17。生长方程各项参数显示,其渐进体长L∞为212.33 mm,渐进体质量W∞为48.22 g,生长系数k为0.43,理论生长起点年龄t0为-0.88龄。凤鲚体长和体质量生长曲线如图7所示。

图7 凤鲚体长和体质量的生长曲线
凤鲚的体长生长速度方程为:dL/dt=91.15e-0.43(t+0.88),体长生长加速度方程为:d2L/dt2=-39.13e-0.43(t+0.88)。凤鲚的体长生长速度和加速度曲线均为不具有拐点的曲线(图8)。体长生长速度曲线随着年龄增加呈下降趋势,表明凤鲚体长增长速度随年龄增加逐渐降低,体长生长加速度曲线一直为负值,且随着年龄增加呈上升趋势,表明凤鲚体长增长加速度随年龄增加逐渐增加。

图8 凤鲚体长生长速度和加速度曲线
凤鲚体质量生长速度和加速度方程分别为:dW/dt=65.67e-0.43(t+0.88)[1-e-0.429 3(t+0.88)]2.11和d2W/dt2=28.19e-0.43(t+0.88)[1-e-0.43(t+0.88)]1.11[3.172 4e-0.43(t+0.88)-1]。如图9所示,体质量生长速度曲线存在一个拐点,该拐点年龄为1.76龄,对应的拐点体长和体质量分别为145.44 mm和14.52 g。在该拐点前,体质量生长加速度曲线为正值,体质量生长速度逐渐增加;在该拐点后,体质量生长加速度曲线为负值,体质量生长速度逐渐减小。

图9 凤鲚体质量生长速度和加速度曲线
3 讨论
HE等[11]指出,凤鲚鳞片没有稳定的年龄结构,并提出了利用凤鲚耳石上的直/弯生长的特征进行凤鲚年龄鉴定的新方法。不同于过去大多数学者关于长江口凤鲚仅为0~2龄的简单年龄结构的认识,该研究认为长江口凤鲚繁殖群体的年龄结构为1~5龄的复杂结构。凤鲚耳石的这一生长特征也在鲚属的刀鲚(Coiliaectenes)和湖鲚(Coiliaectenestaihuensis)得到验证[19],并应用于其年龄鉴定,但对于鲚属鱼类耳石特征的形成机制至今尚不完全清楚[22]。本研究及其他基于耳石年龄鉴定方法的多数研究均表明,长江口凤鲚繁殖群体存在3~5龄的高龄个体,其中1~2龄群体为优势群体,几乎占到全部个体的80%左右[5,11-13,22]。
鱼类生长过程中体长和体质量的关系及相关参数,对于鱼类生态学基础理论研究、鱼类生长的表达和渔业管理有重要意义。目前,最常用的鱼类体长-体质量关系式为W=aLb,式中b值反映了不同生长发育阶段和环境中鱼类的生长特征[23]。处于幼鱼阶段b值一般小于3,呈负异速生长,随着发育逐渐完成后,成鱼阶段b值一般等于或大于3[24]。本研究结果显示,长江口雄性凤鲚体长、体质量均显著小于雌性凤鲚(P<0.05),雄性凤鲚b值(2.87)明显小于雌性凤鲚b值(3.31),与曾强与董方勇[6]及于晓[12]的研究结果基本一致(表3)。进一步t检验结果表明,长江口凤鲚雌、雄性繁殖群体的b值存在较大差异,其中雄性b值(2.87)与“3”无显著性差异,说明其倾向于匀速生长,而雌性b值(3.31)与“3”存在显著性差异(P<0.05),显示其为正异速生长。此外,如表3所示,本研究中长江口雌性凤鲚b值为3.31,远高于王焕焕等[5]2014年调查结果(b=2.75),说明长江口雌性凤鲚个体生长状况得到有效恢复的可能性较大,可能得益于近年来长江口海域所实施的生态环境保护政策等。凤鲚雌雄个体间的生长差异特征,可能与鱼类进化过程中的繁殖策略有关,在一定程度上体现了鱼类对外界压力的适应能力。即鱼类种群为了适应一定环境与捕捞压力的变化,通过调节其雌性个体体质量生长速度,促进雌性个体性腺发育和生殖洄游能量的积累等[25],从而达到相对提高种群繁殖力,促进种群生存和繁衍[26]。
如表3所示,本研究的长江口凤鲚拐点年龄约为1.81龄,与HE等(2006—2007年,1.77龄)[11]、于晓(2012—2013年,1.72龄)[12]的结果大致相同,但与王焕焕等(2014年,1.11龄)[5]的差异较大,一是可能与当年调查的采样时间和样本数量等有关,二是可能得益于近年来长江口海域生态环境的持续改善。由于凤鲚性成熟年龄为1龄[6],其拐点年龄增大,意味着凤鲚性成熟后的体质量生长速度还会持续增加一段时间,有利于提高其繁殖能力。由此可推断出,在长江口凤鲚资源恢复过程中如果采用适当增大刺网网目尺寸和推迟开捕时间等管理措施,降低对0~1龄鱼特别是0龄世代的捕捞,可以达到资源养护和提高资源利用效率的管理目标。此外,表3还显示,自20世纪以来,长江口凤鲚渐进体长L∞及平均体长和体质量均呈明显降低的趋势,但本研究结果是否预示近年来长江口凤鲚个体小型化趋势可能已被有效遏制,尚有待于用进一步的数据监测来验证,其原因也有待进一步探讨。

表3 长江口凤鲚生长特性比较
管卫兵等[9]认为,长江口汛期表层水温是影响长江口凤鲚繁殖特性的环境因子之一,水温升高后凤鲚生殖群体的丰满度和性腺质量下降[9]。其原因可能是水环境中温度升高导致鱼类代谢增强、体内营养物质积累减缓,并最终影响其群体的成熟[27-29]。倪健夫等[29]指出,长江口水域4—6月的表层水温在1982—2014年间总体呈现增高趋势,并曾在1997年前后发生模态转变(regime shift),即在1997年之后,长江口水域4—6月表层平均水温较之前年份平均上升约2 ℃,且与凤鲚总捕捞量和单船全汛捕捞量呈显著负相关关系[30]。有关长江口凤鲚生物学特性变动(如个体小型化等)对水温等环境因素的响应,尚需进一步研究。
综上,在长江全面禁捕背景下,如何开展长江口凤鲚等重要渔业资源评估及其影响机制的研究,还需深入开展工作,以便为长江口渔业管理措施的制定提供更有益的科学依据。
猜你喜欢
保健与生活(2023年23期)2023-12-26 13:10:40
天天爱科学(2022年9期)2022-09-15 01:12:28
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
中华养生保健(2020年7期)2020-11-16 01:14:26
老友(2018年7期)2018-08-08 15:32:30
科学生活(2017年9期)2017-10-16 02:05:08
海洋渔业(2016年6期)2016-04-16 03:00:35
海洋渔业(2016年6期)2016-04-16 03:00:32
华东师范大学学报(自然科学版)(2014年3期)2014-03-11 16:18:19
